Изменение климата и его влияние на жизненное состояние лесостепных популяций сосны обыкновенной русской равнины

Автор: Кузнецова Н.Ф.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесоведение и лесоводство
Статья в выпуске: 2, 2023 года.
Бесплатный доступ
Рассмотрены проблемы изменения климата и последствий его воздействия на сосновые леса лесостепного района Русской равнины. Анализ среднегодовых температур показал, что в начале XXI в. регион вступил в фазу быстрых климатических изменений. За последние 15 лет темпы потепления ускорились в 1,7 раза, число засух удвоилось, количество оптимальных лет за 10-летний период снизилось с 8 до 5. Обобщены результаты 35-летнего мониторинга систем семенного размножения сосны обыкновенной (Воронежская обл., Ступинский тест-объект, 30 деревьев). Исходно лесостепные популяции представляли собой устойчивые равновесные системы, и основные элементы семенного размножения - самофертильность, полнозернистость, число семян на шишку - находились в динамическом равновесии со средой обитания. Потепление климата, 8-летняя тепловая волна и засухи вызвали в 2015 г. переход сосновых лесов в более низкое жизненное состояние - слабонеравновесной системы. Установлено, что для возврата в равновесие им потребовалось 3 оптимальных года. В настоящее время северная и центральная популяции находятся в устойчивом равновесии, жизненное состояние южной лесостепной более низкое - неустойчивого равновесия. Дальнейшее потепление климата повышает вероятность повторной дестабилизации сосновых лесов, а уменьшение числа оптимальных лет исключает возвращение в исходное состояние.
Сосна обыкновенная, лесостепь, потепление климата, мониторинг, жизненное состояние, семенная продуктивность
Короткий адрес: https://sciup.org/143180083
IDR: 143180083 | УДК: 630.161 | DOI: 10.24419/LHI.2304-3083.2023.2.02
Climate change and its impact on the vital state of forest-steppe populations of the common pine of the Russian plain
The problems of climate change and its effects on the pine forests of the forest-steppe region of the Russian plain are considered. The analysis of average annual temperatures showed that at the beginning of the XXI century the region entered a phase of rapid climatic changes. Over the last 15 years the rate of warming accelerated 1.7 times, the number of droughts doubled, the number of optimal years decreased from 8 to 5 / 10 years. The results of 35-year monitoring of seed multiplication systems of Scots pine (Voronezh region, Stupino test object, 30 trees) are summarized. Initially, forest-steppe populations represented stable equilibrium systems and the main elements of seed reproduction - self-fertility, proportion of plump seeds, number of seeds per cone were in dynamic equilibrium with the habitat. Climate warming, an 8-year heat wave, and droughts caused pine forests to transition to a lower life state in 2015, a weakly non-equilibrium system. It was found that it took them three optimal years to return to equilibrium. At present, the northern and central populations are in stable equilibrium, the life state of the southern forest-steppe is lower - unstable equilibrium. Further climate warming increases the probability of pine forests destabilizing again, and a decrease in the number of optimal years precludes a return to the initial state.
Текст научной статьи Изменение климата и его влияние на жизненное состояние лесостепных популяций сосны обыкновенной русской равнины
Лесостепь Русской равнины характеризуется недостаточным и неустойчивым увлажнением. Отличительной особенностью рельефа является разветвленная овражно-балочная система и широкие равнины, которые чередуются с низменными участками. Формирование климата определяется преимущественно западным переносом воздушных масс [1]. Соотношение тепла и влаги близко к оптимальному. Испаряемость меньше количества выпадающих осадков. Основной тип погодного стресса – засуха с частотой 2–3 раза в 10-летие. Природно-климатические условия лесостепи способствуют формированию лесной и лесостепной растительности. Естественный уровень лесистости – около 26 % [2]. Это зона физико-географического оптимума сосны обыкновенной ( Pinus sylvestris L.), где путем естественного отбора сформировались равнинные популяции, для которых характерен оптимальный набор генотипов без четких различий по морфологической и генетической структуре [3].
При анализе состояния сосновых лесов в центре лесостепного района, северной и южной переходных зонах Русской равнины мы использовали такое понятие, как принцип эмерджентности [4], согласно которому каждому иерархическому уровню присуща своя генетическая организация. Учитывался тот факт, что леса на территории ареала могут находиться в равновесном, неравновесном и мертвом состоянии [5]. Равновесные и неравновесные жизненные состояния в свою очередь подразделяются на 2 формы – более и менее устойчивую, популяции и деревья в которых различаются по структурным и функциональным показателям (класс бонитета, урожайность, реакция на стресс и др.).
С середины ХХ в. Земля вступила в эпоху быстрых климатических изменений [6, 7]. В XXI в. увеличилось число экстремальных явлений, засух и тепловых волн. По прогнозам, дальнейшее потепление климата может превысить экологическую емкость лесов. Несоответствие среды обитания и биологии лесных древесных видов ведет к ослаблению лесов, переходу в более низкое жизненное состояние и даже гибели [7–12]. В России темпы потепления климата в 2,5 раза выше роста глобальной температуры. Воронежская и Курская области входят в так называемую красную зону страны. Поэтому сосновые леса Центрально-Черноземного района (ЦЧР), произрастающие на южной границе лесного покрова Русской равнины, где баланс взаимодействий «генотип – среда» находится на нижнем пределе устойчивости, подвержены самым большим рискам [11, 13].
В последние десятилетия актуальной проблемой современности стал поиск решений по адаптации лесов к потеплению климата. Установлено, что устойчивость к засухе – это не столько выживание вида в период ее воздействия, сколько его способность воспроизводить жизнеспособное потомство [14]. В сельском хозяйстве работы в данном направлении принято проводить по результатам репродуктивной способности растений в условиях полевого эксперимента [15]. У сосны обыкновенной, как вида-индикатора для оценки жизненного состояния, используют показатели биопродуктивности шишек [11, 16].
Цель исследований – оценить темпы потепления климата на территории лесостепного района ЦЧР, определить взаимосвязь признаков семенной продуктивности сосны обыкновенной по результатам 35-летнего мониторинга на базе Ступинского тест-объекта, охарактеризовать текущее состояние популяций сосны обыкновенной в центре лесостепного района Русской равнины, южной и северной переходных его зонах.
Объекты и методы исследований
Объектом мониторинга являются 56-летние культуры сосны обыкновенной, произрастающие вблизи Воронежского биосферного природного заповедника на землях с ненарушенным почвенным горизонтом [17] (Воронежская обл., Рамонский район, центральная часть лесостепи, Ступинский тест-объект, контроль). Лесистость Рамонского района составляет 30 %, что превышает средний областной показатель в 3,7 раза и соответствует исходному уровню лесистости
Воронежской обл. (31 %). Песчаные почвы на левом берегу р. Воронеж полностью соответствуют биологии вида Pinus sylvestris L. Северный опытный объект (Калужская обл., Козельский район) расположен в переходной зоне между двумя районами – лесостепным и широколиственных (смешанных) лесов и характеризуется достаточным увлажнением (650 мм осадков в год). Южный опытный объект (Белгородская обл., Красненский район) находится на границе между лесостепью и степью.
Популяционные выборки контрольного и опытных объектов включали по 30 случайно отобранных деревьев. Объем пробы с каждого дерева составлял 10–15 неповрежденных шишек. Состояние систем семенного размножения Ступинского тест-объекта оценивалось по глазомерной 6-балльной шкале В.Г. Каппера (0–5 баллов) и трем признакам семенной продуктивности – самофертильность (%), полнозернистость (%) и число семян на шишку (шт.), которые отражают разные этапы формирования генотипического состава семян [16]. Искусственное самоопыление проводили на одних и тех же деревьях ступинской популяции в течение 12 лет (случайная выборка, 50 деревьев, 1986–1998 гг.) по методике Н.В. Котеловой [18]. У калужской и белгородской популяций при свободном опылении изучали обилие шишек и 2 продукционных признака – полнозернистость и число семян на шишку.
Коэффициент самофертильности (Ксф) деревьев вычисляли как соотношение полнозер-нистости семян при самоопылении и свободном опылении. Уровень полнозернистости определяли как отношение полных семян к общему их числу при свободном опылении. При подсчете числа семян в шишках учитывали общее количество полных семян и пустых капсул, которые достигли минимального размера семени для данной шишки. Максимальный интервал значений этих признаков составляет: коэффициент самофертильности – 0–0,75, полнозернистость – 0–100 % и число семян на шишку – 0–50 шт. Для оценки внутрипопуляционной структуры выборок диапазон изменчивости продукционных признаков был разбит на 4 класса продуктивности согласно методике [19]: по полнозернистости семян: I – 0,0–24,9 %; II – 25,0–49,9 %; III – 50,0–74,9 %; IV – 75 % и выше; по числу семян на шишку: I – 0,0–9,9; II – 10,0–19,9; III – 20,0–29,9; IV– 30 семян и более. Средний показатель выборки по каждому признаку определяли путем суммирования индивидуальных данных и деления полученной суммы на число анализируемых деревьев.
Для статистической обработки использовали пакет программного обеспечения MS Excel-2010. Оценку силы влияния фактора «гидротермический коэффициент (ГТК)» на показатели пол-нозернистости семян объектов проводили с помощью двухвыборочного t-теста с различными дисперсиями.
Результаты и обсуждение
Экологический мониторинг является основным инструментом контроля природной среды. Для оценки состояния и устойчивости сосновых лесов на юге Русской равнины, определения границ значений климатических факторов, выход за пределы которых вызывает переход их в более низкое жизненное состояние, нами отобрана типичная по семенной и вегетативной продуктивности равнинная лесостепная популяция на севере Воронежской обл. Природно-климатические условия Ступинского тест-объекта исходно соответствовали биологии вида Pinus sylvestris L. (ТЛУ А2, сосняк травяной). Среднегодовая температура в центре лесостепи по сравнению с югом ниже на 2,2 °С. Осадков выпадает больше на 150 мм (500–600 мм в год). ГТК находится в пределах от 0,9 до 1,2.
Изменение климата в центре и на юге ЦЧР
В настоящее время в ЦЧР сложилась сложная климатическая ситуация. Темпы потепления климата на его территории превышают общероссийский уровень. Проведен сравнительный анализ изменения регионального климата за годы мониторинговых исследований в центре (Ступинский тест-объект) по сравнению с исходными его показателями для юга лесостепного района (Каменная Степь) ЦЧР.
Первые сведения о состоянии лесостепного климата Русской равнины получены в Каменной Степи (Воронежская обл., Таловский район, юг лесостепной зоны), где метеорологический мониторинг осуществляется с 1894 г. [20]. По данным НИИ сельского хозяйства Центрально-Черноземной полосы им. В.В. Докучаева, среднегодовая многолетняя температура за 56-летний период (1894–1950 гг.) составляла 5,3 °С, диапазон среднегодовых показателей – 3,5–7,7 °С. ГТК по годам изменялся в довольно узких пределах – 1,0–1,4. Летом его уровень снижался до 62–67 %, в засушливые годы – до 50 %. Число холодных и засушливых лет колебалось в пределах 3–5 за 10-летие. Среднегодовое количество осадков – 400–500 мм при варьировании по годам 300–700 мм и более. При анализе изменения лесостепного климата в центре ЦЧР показатели Каменной Степи мы использовали в качестве нормы, которая в течение многих веков была оптимальной для произрастания лесной и степной растительности.
Мониторинговые исследования сосны обыкновенной ступинской популяции начали проводить в 1986 г. Усредненные за 5-летние периоды данные среднегодовой температуры, количества осадков и диапазон годовых показателей в течение 7 временных интервалов с 1986 по 2020 г. представлены в табл. 1. Отметим, что с 1945 по 1985 г. отклонения среднегодовых температур от средней многолетней были несущественными: превышение составляло 0,11 °С за 10 лет. Среднегодовое количество осадков соответствовало региональной норме.
Анализ семи 5-летних интервалов выявил, что за годы мониторинга в районе исследований самая низкая среднегодовая температура отмечена в 1-й период (1986–1990 гг.) – 6,3 °С, при варьировании годовых показателей от 3,7 до 8,0 °С (см. табл. 1). Сравнение данных первого 5-летнего периода в центре лесостепного района с региональной нормой на южной его границе (Каменная Степь) показало, что в конце 1980-х гг. на территории ЦЧР наметился явный сдвиг температурного режима в направлении потепления. Так, среднегодовая температура первой половины ХХ в. на юге лесостепи составляла 5,3 °С, что на 1,0 °С ниже, чем в центральной части в 1986–1990 гг. (6,3 °С). Диапазон минимальных и максимальных температур в Каменной Степи и на Ступинском тест-объекте за
Таблица 1. Изменение среднегодовой температуры и количества осадков за 35 лет мониторинга, Ступинский тест-объект*
|
Период, годы |
Температура, °С (диапазон значений ) |
Осадки, % нормы (диапазон значений) |
|
1986–1990 |
6,3 (3,7–8,0) |
108 (83–130) |
|
1991–1995 |
6,5 (5,5–7,9) |
100 (82–123) |
|
1996–2000 |
6,7 (5,9–7,8) |
94 (72–109) |
|
2001–2005 |
7,3 (6,3–7,8) |
110 (87–130) |
|
2006–2010 |
7,9 (6,7–8,5) |
96 (75–116) |
|
2011–2015 |
8,0 (7,1–8,7) |
97 (73–141) |
|
2016–2020 |
8,2 (7,6–9,5) |
110 (81–149) |
* Контроль: 5,3 °С (3,5–7,7 °С) – Каменная Степь, 1896–1950 гг.
эти годы также несколько сместился в сторону более высоких значений: 3,5–7,7 и 3,7–8,0 °С соответственно. При этом количество выпавших осадков по-прежнему находилось в пределах региональной нормы.
За годы наблюдения среднегодовые температуры постоянно повышались (см. табл. 1). Однако их увеличение происходило неравномерно. Усредненные данные отчетливо выявляют 2 тренда изменения 5-летних среднегодовых температур. Для региона выявлен год перехода от плавного равномерного к ускоренному этапу изменения климата.
При плавном потеплении климата (20-летний временной интервал – 1986–2006) увеличение среднегодовой температуры составляло 0,2 °С каждые 5 лет. В сторону повышения смещалась только ее нижняя граница: с 3,7 до 5,9 °С. Верхний ее предел в течение 20 лет оставался на одном и том же уровне – 7,7–7,9 °С.
Переход от плавного к ускоренному этапу изменения климата совпал с началом 8-летней тепловой волны, которая на территории ЦЧР протекала на фоне снижения уровня грунтовых вод и череды засух 2007, 2010, 2012, 2014 гг. По напряженности 3 засухи были сильными или уровня экологической катастрофы (2010 г.). Возникновение тепловой волны связано с блокированием западного переноса воздушных масс, перемещением жаркого тропического воздуха и длительным его стационированием в центре и на юге Русской равнины [21]. В эти годы темпы потепления климата резко возросли. ГТК на южной границе лесостепного района снизился до уровня полупустынь – 0,5–0,6. Если в летний период 2000–2006 гг. наблюдалось некоторое превышение увлажнения по индексу SCEI (от 0 до +0,5), то с 2007 по 2012 г. он составлял от -1 до -1,5. Это означает, что практически весь лесостепной район Русской равнины стал засушливым. Уровень активных температур по сравнению со среднемноголетним показателем повысился на 250–350 °С и достиг 2 600–3 000 °С [22].
В центре лесостепного района среднегодовая температура в четвертый 5-летний период
(2006–2010 гг.) составила 7,9 °С, т.е. повысилась на 0,6 °С (см. табл. 1). При этом обе ее границы сместились в сторону более высоких значений (6,7–8,5 °С): максимальная температура – с 8,0 до 8,5 °С, минимальная – с 6,3 до 6,7 °С. Несмотря на то что в 2011–2015 гг. темп прироста среднегодовой температуры снизился до 0,2 °С, нижняя ее граница достигла нового максимума – 7,1 °С. Во время последнего 5-летнего периода (2016–2020 гг.) наблюдалась аналогичная ситуация. Рост среднего показателя составил всего 0,2 °С, нижняя граница среднегодовой температуры поднялась на 0,5 °С (с 7,1 до 7,6 °С), а верхняя – сразу на 0,8 °С (с 8,7 до 9,5 °С). Резкое смещение максимального показателя обусловлено тем, что два последних года седьмого периода (2019–2020 гг.) оказались самыми теплыми за всю историю метеонаблюдений. Среднегодовая температура составила 8,3 °С в 2019 г. и 9,5 °С в 2020 г. в центре лесостепи, что выше максимального среднегодового показателя Каменной Степи на 0,6 °С и 1,8 °С соответственно. В 2021 г. среднегодовая температура была также высокой – 8,4 °С.
За годы мониторинга самым дождливым оказался 2016 г. В этот год в районе исследований выпало 1,5 нормы годовых осадков, большая часть из которых пришлась на весенний период: превышение составило 263 %. Количество выпавших осадков за последний 5-летний период близко к норме – 110 % (81–149 %).
Проведенный анализ свидетельствует о том, что потепление климата в лесостепи, с которым тесно взаимосвязаны процессы роста и развития сосны, продолжается. Более того, в XXI в. его темпы ускорились.
В настоящее время превышение среднегодовой температуры в центре лесостепи над средней многолетней на юге составляет 2,9 °С. Число оптимальных лет уменьшилось с 8 до 5 за 10-летие. Уровень грунтовых вод снизился до 4–5 м. Тревожным фактом является то, что засушливыми были 4 года подряд (2018–2021 гг.). Среднегодовое количество осадков сохраняется на уровне региональной нормы, что, безусловно, положительный фактор.
Мониторинг систем семенного размножения ступинской популяции сосны обыкновенной
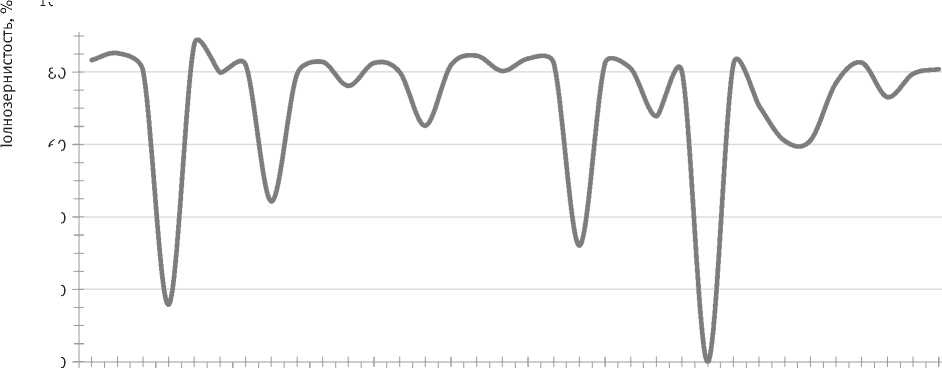
Развитие всех саморегулирующихся систем, к числу которых относятся лесные экосистемы, базируется на 3-х принципах сохранения: сохранение нормы вида, генетического разнообразия и устойчивости [23]. Большинство сосновых лесов Русской равнины характеризуется высокой вегетативной и семенной продуктивностью, сбалансированностью генотипического состава и устойчивостью к региональным стрессорам [24]. Ступинская популяция сосны обыкновенной относится к их числу, является объектом мониторинговых и комплексных исследований [11, 16, 25]. Лесной массив до 2015 г. развивался в режиме устойчивого равновесия, при котором генеративная сфера и основные элементы систем семенного размножения в оптимальные годы характеризовались довольно высокой стабильностью. Урожайность большинства деревьев высокая, реакция на раннюю засуху пропорциональна силе погодного стресса, после засухи генеративная сфера всегда возвращалась к норме (рисунок).
Проводимые с 1986 г. мониторинговые исследования и 12-летний опыт по самоопылению показали, что основными элементами систем семенного размножения сосны на популяционном и организменном уровне являются самофертильность, полнозернистость и число семян на шишку. Между данными показателями существует обратно пропорциональная зависимость, и они как генетически обусловленные и экологически зависимые признаки принимают участие в адаптации семенного генофонда к погодным условиям года [11]. Их внутрипопуляционная структура находится в прямой зависимости от погодных условий года, тесно связана с адаптивной нормой реакции вида, популяции и генотипа. В оптимальные годы данные признаки задействованы в формировании по принципу взаимодействия «генотип – среда» сбалансированного генотипического состава лесостепных популяций, в передаче от поколения к поколению нормы вида [26, 27]. Сдвиг нормы реакции в аномальные годы тестирует степень влияния среды на их вегетативную и семенную продуктивность. В раннюю засуху по ее изменению на организменном уровне определяется уровень засухоустойчивости [16]. Происходит дифференциация популяций на контрастные по уровню засухоустойчивости группы деревьев – чувствительная, модальная (деревья с модальными значениями) и устойчивая. Показано, что отклик на засуху представляет собой генетически регулируемый процесс, который на базе неспецифической
2010 2012 2014
Изменение уровня полнозернистости ступинской популяции сосны обыкновенной в годы мониторинга (по горизонтали обозначены годы засух)
и специфической реакции вызывает направленный сдвиг адаптивной нормы реакции признаков самофертильности, полнозернистости и числа семян на шишку, корректируя таким образом генотипический состав выживших семян в направлении его адаптации к более теплому и сухому климату.
Необходимо отметить, что 22 года из 35 лет мониторинга по погодным условиям были близки к региональной норме. В оптимальные годы сосна характеризуется устойчивым и сбалансированным развитием генеративной сферы, что подтверждается способностью большинства деревьев сохранять на разных иерархических уровнях свои количественные характеристики до определенных пределов флуктуации внешней среды. Цветение и семеношение ежегодное и достаточно обильное – 4 балла по шкале Каппера. Уровни полнозернистости изменяются в пределах 76,2–87,7 % с модой 81,6±1,3 %. Частотное распределение деревьев по классам продуктивности приблизительно одинаковое: 4/5 деревьев относятся к IV высокопродуктивному классу, III класс включает приблизительно 1/4 форм, II класс представлен 1–2 растениями, деревья I класса отсутствуют. Число семян на шишку составляет 21,2±1,2 шт. при варьировании по годам средних значений от 19,6 до 26,1 шт., при этом 3/4 значений приходится на центр вариационного ряда. Частота патологий митоза в 3,5 раза ниже нормы спонтанного мутирования (5 %) [11]. Средний уровень самофертильности выборки низкий – 0,13–0,14. Доля инбредного потомства в семенной продукции года составляет 7–8 % [16]. Уровень пролина в хвое деревьев низкий – 3,0–3,7 мкМ/г [11].
Вегетативная сфера сосны по сравнению с генеративной более устойчива к засухе. Наиболее сильное воздействие на развитие генеративной сферы оказывает ранняя засуха. Погодный стресс выводит сосновые леса из равновесия, вызывает смещение нормы реакции экологически зависимых признаков и фазовый переход деревьев из одного равновесного жизненного состояния в другое в направлении поддержания жизнеспособности самого растения и адаптации его семенного потомства к более сухому и теплому климату. Ответная стресс-реакция, которая протекает на базе неспецифической и специфической реакций, повышает устойчивость выживших семян к более сухому и теплому климату путем увеличения доли инбредных генотипов и потомств от засухоустойчивых форм [16].
В коррекции генотипического состава семян задействованы механизмы, среди которых на популяционном и организменном уровне основными являются самофертильность, полно-зернистость и число семян на шишку (табл. 2). Принцип их действия представляет авторегули-руемый процесс, который заложен в самой природе признаков. Показатели алгоритма изменения нормы реакции, периода действия и порога чувствительности к погодному стрессу взаимно перекрываются. Инбридинг и кроссбридинг
Таблица 2. Формула вычисления и характеристика признаков семенной продуктивности сосны обыкновенной
|
Признак |
Формула |
Алгоритм изменения |
Период действия |
Порог чувствительности |
|
Самофертильность, Ксф |
Рсо Ксф =----- Рсв оп |
Расширение |
Гаметогенез, эмбриогенез |
Теплые, холодные годы |
|
Полнозернистость, Pсв оп |
Nполн Рсвоп =----------- Nполн + пуст |
Расширение – сужение |
Эмбриогенез |
Слабая засуха |
|
Число семян на шишку, Nполн+пуст |
– |
Сужение |
Гаметогенез |
Сильная засуха |
Примечание: Ксф – коэффициент самофертильности, Рсо – полнозернистость семян при самоопылении, Рсв оп – полно-зернистость семян при свободном опылении, Nполн+пуст – число семян на шишку.
в рамках свободного опыления также подвижны. На изменение погоды в формулах вначале реагирует числитель, при усилении погодного стресса изменяются числитель и знаменатель. В стрессовом градиенте засушливых лет норма реакции признака самофертильности только расширяется, по числу семян сужается, по полнозер-нистости вначале максимально расширяется, а в сильную засуху меняет направление и смещается вплоть до 0. Вероятно, эти особенности структурной и функциональной взаимосвязи признаков составляют основу устойчивости систем семенного размножения равновесных сосновых лесов в нормально колеблющейся среде и в засушливые годы.
За годы мониторинга в районе исследований произошло 7 весенне-летних засух разной напряженности (1991, 1995, 2001, 2007, 2012, 2014, 2019 гг.) и летняя засуха 2010 г. (см. рис. 1). Самая сильная и продолжительная из них – засуха 2010 г. Так как по срокам она была летней, то слабо отразилась на показателях биопродуктивности шишек. Из-за аномальной жары потомство сосны из семян урожая данного года оказалось практически полностью нежизнеспособным.
Установлено, что анализируемые признаки по-разному реагируют на весенне-летнюю засуху разной напряженности. Начало снижения уровня полнозернистости зафиксировано в слабую засуху. Показатели ступинской популяции в такие годы (2001, 2014, 2019) уменьшились на 10–25 %. Диапазон изменчивости деревьев стал шире: группа устойчивых деревьев не отреагировала на погодный стресс, степень редукции чувствительной группы составила от 1,2 до 1,6 раз. В умеренную засуху 1995 г. урожайность снизилась почти вдвое (44,3±3,4 %), диапазон значений признака достиг максимума (0–85 %), разница между контрастными группами стала 3-кратной. В сильную засуху отличия с оптимальными годами достигли 3-х (2007 г.) и 5-кратного (1991 г.) уровня. При переходе от умеренной к сильной засухе произошла смена алгоритма изменчивости нормы реакции полнозернистости – от расширения к сужению (см. табл. 2). Более сильный погодный стресс весной 2012 г. привел к полному опаду шишек
2-го года развития в сосновых лесах ЦЧР. Число семян на шишку характеризуется самым низким порогом чувствительности к засухе. В слабую и умеренную засуху средние значения этого признака оставались на уровне оптимальных лет. Их количество резко снизилось в сильную засуху 2007 и 1991 г.
Данные мониторинга свидетельствуют о том, что генотипический состав семян засушливых лет напрямую зависит от силы погодного стресса. Так, количество инбредных семян в семенной продукции года увеличилось с 1/12 (оптимальный год) до 1/9 и 1/3 (умеренная и сильная засуха соответственно). Число потомств от засухоустойчивых форм при слабой, умеренной и сильной засухе повысилось от 1/11 (оптимальный год) до 1/7, 1/5, 1/3 соответственно. В урожаях засушливых лет существенно изменился вклад модальной группы деревьев. Известно, что она выполняет такую важную функцию половой репродукции, как сохранение и передача от поколения к поколению нормы вида. Как показал анализ, ее объем в стрессовом градиенте засушливых лет сократился с 70 % (оптимальные годы) до 67,8, 51,2 и 32,3 % соответственно при слабой, умеренной и сильной засухе. В отношении реализации данной функции в аномальные годы ведущая роль принадлежит именно модальной группе деревьев, несмотря на 2-кратный уровень редукции.
Высокие темпы потепления климата лесостепного района Русской равнины и уязвимость сосновых лесов на юге бореальной зоны повышают вероятность перехода их в более низкое жизненное состояние – слабонеравновесной системы, которое является энергетически менее затратным. Высвобождающаяся при этом энергия расходуется на перестройку метаболизма и поддержание гомеостаза в условиях более сухого и теплого климата. Функционирование, жизненное состояние и продолжительность жизни равновесного и неравновесного леса существенно отличаются. Для последнего характерна более высокая разбалансированность структуры и процессов. Следствием дисбаланса является увеличение изменчивости морфологических, анатомических, цитогенетических и других признаков. Так как структуры и функции связаны между собой, то в этом случае страдают вегетативная и генеративная сферы деревьев. По-ви-димому, это один из способов выживания вида, при котором его биология и экологическая ниша совпадают лишь частично [28]. Неравновесные сосновые леса – обычное явление в природе (северная и южная границы ареала, болота, высокогорья, техногенно загрязненная территория и др.). Все они характеризуются пониженной семенной и вегетативной продуктивностью, а также низким качеством семян.
В сложных системах, к числу которых относятся лесные экосистемы, процессы могут протекать в прямом и в обратном направлении. Немаловажной особенностью устойчивого равновесия является то, что при отклонении от него система стремится вернуться в исходное состояние. Поэтому генеративная сфера ступинской популяции на следующий год после 6-ти засушливых лет всегда возвращалась к региональной норме – минимальному уровню инбредных семян и высоким показателям продуктивности шишек (см. рисунок), подтверждая, что ее развитие до 2015 г. происходило в режиме устойчивого равновесия.
Весной 2015 г., несмотря на оптимальные погодные условия, на всей территории ЦЧР произошла дестабилизация сосновых лесов. В конце 8-летней тепловой волны в регионе отмечено 2 засухи – слабая весенняя и сильная продолжительная осенняя. Почвенная засуха 2014 г. стала толчком для перехода сосны на 7 объектах Белгородской, Липецкой, Курской и Воронежской областей из основного равновесного в слабонеравновесное состояние. Биопродуктивность шишек в 2015 г. стала ниже региональной нормы на 25 % и меньше уровня засушливого 2014 г. на 13,5 %. Число семян уменьшилось на 1/3, уровень пролина весной повысился в 2,8 раза [11]. Снизился годовой прирост деревьев, наблюдался массовый опад хвои, вспышки массового размножения вредителей и болезней леса. Выход сосновых лесов из состояния дестабилизации продолжался в течение 3-х лет (2015–2017), которые по погодным условиям соответствовали региональной норме.
Состояние популяций сосны в центре лесостепного района, северной и южной переходных зонах
Текущее состояние генеративной сферы сосны изучалось на случайных выборках 3-х равнинных популяций – калужской, воронежской и белгородской. Все они расположены на экологически благоприятной территории в северной, центральной и южной части лесостепной зоны Русской равнины. В 2020 г. оптимальный температурный режим на анализируемой территории сохранялся вплоть до августа. Среднее значение ГТК за май–июль на объектах, а также сила влияния фактора ГТК на полнозернистость сосны с севера на юг представлены в табл. 3.
Калужская популяция – северный опытный объект, который расположен между южной подзоной широколиственных лесов и лесостепью. Анализ состояния генеративной сферы насаждения проводился по признаку полнозернистости. В Калужской обл. количество выпавших осадков в 2020 г. превысило многолетнюю норму на 15 %. Средний показатель ГТК в мае–июле был равен 2,1, что в 1,5 раза выше региональной нормы
Таблица 3. Влияние фактора ГТК на полнозернистость семян калужской, ступинской и белгородской популяций сосны обыкновенной
|
Популяция |
ГТК (май–июль) |
Полнозернистость семян, % |
Сила влияния фактора, % |
|
Калужская |
2,1 |
81,6±1,5 |
- |
|
Воронежская |
1,0 |
80,8±1,3 |
2,5 |
|
Белгородская |
0,7 |
61,4±3,0* |
57,8 |
Различия достоверны при Р < 0,05.
(см. табл. 3). Интенсивность семеношения по сравнению со ступинской популяцией более низкая (3 балла). Уровень полнозернистости семян (81,6±1,5 %) укладывается в пределы модальных значений Ступинского тест-объекта (76,2–87,7 %). Диапазон изменчивости деревьев составляет 62,4–92,6 %. Распределение деревьев по классам продуктивности имеет ярко выраженную асимметричную кривую с модой в зоне высокопродуктивных деревьев IV класса. Выборка представлена деревьями двух высших (III и IV) классов продуктивности; коэффициент вариации признака составляет 11,4 %, что свидетельствует об ее однородности. Семена калужской популяции характеризуются высоким качеством: лабораторная всхожесть – 86,0±2,1 %.
В Воронежской обл. ГТК в мае–июле находился в пределах региональной нормы – 1,0 (см. табл. 3). По месяцам наблюдался значительный разброс значений: дождливый май – 1,6, оптимальный июнь – 1,0, засушливый июль – 0,4. Высокая урожайность сосновых лесов в 2020 г. обусловлена тем, что наиболее чувствительные стадии репродуктивного цикла сосны приходились на конец апреля, май и июнь. Так, полно-зернистость семян ступинской (воронежской) популяции составила 80,8±1,3 % при диапазоне значений 56,6–90,4 %. Выборка представлена деревьями только III и IV классов продуктивности. Внутрипопуляционная изменчивость признака низкая (CV = 9,2 %). Лабораторная всхожесть семян – 86,5±1,2 %. Несмотря на то что ГТК на севере лесостепи по сравнению с центральной ее частью в 2,1 раза выше, сила его влияния крайне мала – 2,5 %, т.е. калужская и воронежская популяции находятся в состоянии устойчивого равновесия.
По спутниковым данным, сосновые леса юго-запада Среднерусской возвышенности в последние годы подвержены усыханию [29]. Изучение самой южной белгородской популяции показало, что она характеризуется обильным цветением и семеношением (4–5 баллов). Уровень полнозернистости (61,4±3,0 %) достоверно ниже по сравнению с северной и центральной популяциями сосны (см. табл. 3). Диапазон значений признака шире и равен 26,3–88,9 %. Коэффициент вариации – 21,3 %, что в 2,3 раза выше, чем у воронежской популяции. Существенно отличается и структурированность выборки. Число классов продуктивности увеличилось до трех, при этом к III классу относится половина деревьев. Представительство II и IV классов по объему сопоставимо: 22,6 и 23,8 % соответственно. Доля изменчивости, обусловленная фактором ГТК, составляет 57,8 % общей изменчивости и существенно влияет на урожайность. При этом важно отметить, что показатели продуктивности шишек белгородской популяции после 2017 г. характеризуются стабильностью: пониженный уровень полнозернистости и много семян в шишках (32,5±1,8 шт.). Лабораторная всхожесть семян высокая (79,7±2,4 %). Из проведенного анализа следует, что все 3 объекта находятся в состоянии равновесия, но в разных его формах: калужская и воронежская популяции – устойчивого, а белгородская – неустойчивого.
Известно, что в число основных функций полового воспроизведения входит сохранение, восстановление и передача от поколения к поколению нормы вида [30]. На основании полученных данных можно констатировать, что внутренняя разбалансировка белгородской популяции, присущая всем системам в неустойчивом равновесии, по отношению к генеративной сфере нивелируется стабилизирующим отбором и проявляется в виде повышенного уровня пустосемянности. Большая часть генетически нарушенных, инбредных, отклоняющихся по форме и размерам зародышей удаляется в ходе эмбриональной селекции. Установлено, что мерные признаки зародышей в раннем эмбриогенезе характеризуются высокой изменчивостью (CV = 54 %), при созревании семян их уровень оценивается как низкий [31]. Опыты по химическому мутагенезу показали, что деревья сосны обыкновенной, выращенные из обработанных химическими мутагенами семян, отличаются низкорослостью и высоким уровнем пустосемянности (28,6±6,4 %, при диапазоне 0–76,6 %) [32]. Большая часть растений следующего поколения по высоте, морфологии и уровню патологий митоза соответствует региональной норме, подтверждая высокую эффективность стабилизирующего отбора в период половой репродукции.
Заключение
Анализ среднегодовых температур показал, что лесостепь Русской равнины характеризуется высокими темпами потепления климата. Данный процесс начался с середины ХХ в. и его протекание подразделяется на 2 этапа – плавного и ускоренного изменения. Различие этапов выражается в разном алгоритме прироста среднегодовых температур. На I-м этапе происходил сдвиг только нижней границы, на II-м отмечался рост максимальных и минимальных температур, увеличение числа засух и тепловых волн, уменьшение в 2 раза количества оптимальных лет.
Согласно данным мониторинга, ступинская популяция сосны до 2015 г. развивалась в режиме устойчивого равновесия. Система семенного размножения и ее элементы (самофертильность, полнозернистость, число семян на шишку) находились в динамическом равновесии со средой обитания. Взаимосвязь, биология и экологическая зависимость данных признаков семенной продуктивности обеспечивали стабильность и высокую урожайность большинства лесостепных популяций сосны в оптимальные годы, упорядоченный на базе неспецифической и специфической реакции отклик на засуху и возвращение генеративной сферы на следующий год к региональной норме. Потепление климата, продолжительная тепловая волна 2007–2014 гг. и 2 засухи в последний год ее протекания вызвали дестабилизацию сосновых лесов ЦЧР и переход в более низкое жизненное состояние – слабонеравновесной системы. Процесс возврата сосны в исходное состояние в оптимальные годы происходил в течение 3-х лет, подтверждая обратимость жизненных состояний.
Изучение текущего состояния лесостепных сосновых лесов свидетельствует о том, что калужская, воронежская и белгородская популяции являются равновесными системами. При этом северная калужская и центральная воронежская популяции находятся в устойчивом равновесии, а форма жизненного состояния южной белгородской популяции более низкая – неустойчивое равновесие. Важная особенность процессов половой репродукции, протекающих в равновесных системах, заключается в том, что большая часть отклоняющихся от видовой нормы зародышей удаляется в процессе стабилизирующего отбора, что проявляется в виде повышенного уровня пу-стосемянности и высоком качестве семян.
Дальнейшее потепление климата, увеличение числа засух и тепловых волн на юге Русской равнины увеличивает вероятность перехода высокопродуктивных сосновых лесов в более низкое (неустойчиво равновесное, слабонеравновесное) жизненное состояние, а уменьшение числа оптимальных лет исключает возможность реализации обратного процесса – возвращение в состояние устойчивого равновесия.
Список литературы Изменение климата и его влияние на жизненное состояние лесостепных популяций сосны обыкновенной русской равнины
- Золотокрылин, А.Н. Районирование России по природным условиям жизни населения / А.Н. Золотокры-лин, А.Н. Кренке, В.В. Виноградова. - Москва: Геос, 2012. - 156 с.
- Молчанов, А.А. Влияние леса на окружающую среду / А.А. Молчанов. - Москва: Наука, 1973. - 359 с.
- Санников, С.Н. Дифференциация популяций сосны обыкновенной / С.Н. Санников, И.В. Петрова. - Екатеринбург: УрО РАН, 2003. - 247 с.
- Stephan, А. Varieties of emergentism / А. Stephan // Evolution and cognition. - 1999. - V. 5. - № 1. - P. 49-59.
- Kramer, K. Equilibrium and non-equilibrium concepts in forest genetic modeling: population- and individually-based approaches / К. Kramer // Forest Systems. - 2011. - Vol. 4. - № 3. - P. 100-109.
- IPCC. Climate Change 2022: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. - Montreal, 2017. - P. 3-29.
- Smith, M.D. The ecological role of climate extremes: current understanding and future prospects / M.D. Smith // J. Ecology. - 2011. - Vol. 99. - P. 651-655.
- Transdisciplinary global change research: the co-creation of knowledge for sustainability / W. Mauser, G. Klepper, M. Rice, B.S. Schmalzauer [et al.] // Curr. Opin. Environ. Sustain. - 2013. - Vol. 5. - Iss. 3-4. - P. 916-917.
- Brack, D. Background analytic study Forest and Climate / D. Brack. - Global Forest Goals. - 2019. - 56 p.
- Climate change and Russian forests: impacts, vulnerability and adaptation needs / C. Reyer, M. Lindner, R. Zamolodchikov, A. Shvidenko [et al.] // Russian forests and climate change. What Science Can Tell Us. - Europ. For. Inst. - 2020. - P. 53-72.
- Кузнецова, Н.Ф. Высокопродуктивные сосновые леса в условиях изменяющегося климата / Н.Ф. Кузнецова, Е.С. Клушевская, Е.С. Аминева // Лесной журнал. - 2021. - № 6. - С. 9-23. DOI: https://doi. org/10.37482/05 36-1036-2021-6-9-23.
- Forests resilience and tipping points at different spapio-temporal scales: approaches and challenges / C. Reyer, N. Brouwers, A. Ramming, B.W. Brook [et al.] // J. Ecology. - 2015. - Vol. 103. - № 1. - P. 5-15.
- Кузнецова, Н.Ф. Состояние лесов и динамика их породного состава в Центральном федеральном округе. - Текст: электронный / Н.Ф. Кузнецова, М.Ю. Сауткина // Лесохозяйственная информация. - 2019. - № 2. -С. 25-45. DOI: 10.24419/LHI.2304-3083.2019.2.03. - Режим доступа: URL: http://lhi.vniilm.ru.
- Akinci, S. Plant water-stress response mechanisms / S. Akinci, D.M. Lösel / Abiotic stresses in major pulses: Current status and strategies. - India: Springer Publ., 2014. - P. 149-172.
- Quarrie, S.A. Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture / S.A. Quarrie, M.M. Olivera // J. Exp. Bot. - 2004. - Vol. 53. - № 5. - P. 1-21.
- Kuznetsova, N.F. Self-fertility in Scots pine as a system for regulating close relationships and species survival in advance environment / N.F. Kuznetsova / Advances in genetics research. - Vol. 9. - New York: Nova Science Publ., 2012. - P. 83-106.
- Середа, Л.О. Оценка эколого-геохимического состояния почвенного покрова городского округа город Воронеж / Л.О. Середа, Л.А. Яблонских, С.А. Куролап // Вестник ВГУ. Сер. География. Геоэкология. - 2015. - № 4. - С. 59-65.
- Котелова, Н.В. Влияние самоопыления и перекрестного опыления на качество семян и сеянцев сосны обыкновенной / Н.В. Котелова // Науч.-техн. информ. - 1956. - № 23. - С. 1-74.
- Кузнецова Н.Ф. Чувствительность генеративной сферы сосны обыкновенной к засухе в Воронежской области / Н.Ф. Кузнецова // Лесоведение. - 2010. - № 6. - С. 46-53.
- Глобальное потепление и особенности изменения климата ЦЧР / И.И. Бартенев, Д.С. Гаврин, М.В. Кра-вец [и др.] / Актуальные вопросы современных сельскохозяйственных наук: матер. Междунар. науч. конф. -Екатеринбург, 2016. - С. 18-20.
- Виноградова, В.В. Волны тепла на европейской территории России в начале XXI века / В. В. Виноградова // Изв. РАН. Сер. География. - 2014. - № 1. - С. 47-55.
- Виноградова, В.В. Динамика увлажнения и теплообеспеченности в переходных ландшафтных зонах по спутниковым и метеорологическим данным в начале XXI века / В.В. Виноградова, Т.Б. Титкова, Е.А. Черенкова // Современные проблемы дистанционного зондирования Земли из космоса. - 2015. - Т. 12. - № 2. - С. 162-172.
- Динамика популяционных генофондов при антропогенных воздействиях / под ред. Ю.П. Алтухова. - Москва: Наука, 2004. - 278 с.
- Романовский, М.Г. Продуктивность, устойчивость и биоразнообразие равнинных лесов Европейской России / М.Г. Романовский. - Москва: МГУЛ, 2002. - 92 с.
- Кузнецова, Н.Ф. Ультраструктурные аспекты физиологической несовместимости у сосны обыкновенной / Н.Ф. Кузнецова, Ю.Н. Исаков // Лесоведение. - 1987. - № 3. - С. 11-16.
- Глотов, Н.В. Норма реакции генотипа и взаимодействие генотип-среда в природной популяции / Н.В. Глотов, В.В. Тараканов // Журн. общей биологии. - 1985. - Т. 44. - № 6. - С. 760-770.
- Модель эколого-генетического контроля количественных признаков растений / В.В. Драгавцев, П.П. Ли-тун, Н.М. Шкель, Н.Н. Нечипоренко // Докл. АН СССР. - 1984. - Т. 274. - № 3. - С. 720-723.
- Кузнецова, Н.Ф. Смена жизненного состояния как способ выживания Pinus sylvestris L. на техногенно загрязненной территории / Н.Ф. Кузнецова, Е.С. Клушевская // Принципы экологии. - 2020. - № 2. - С. 40-47.
- Терехин, Э.А. Оценка нарушенности лесов лесостепной зоны в начале XXI века по спутниковым данным / Э.А. Терехин // Современные проблемы дистанционного зондирования Земли из космоса. - 2020. - Т. 17. - № 2. - С. 134-146.
- Щербаков, В.П. Эволюция как сопротивление энтропии: II. Консервативная роль полового размножения / В.П. Щербаков // Журнал общей биологии. - 2005. - Т. 44. - № 4. - С. 300-309.
- Свинцова, В.С. Характер изменчивости признаков репродуктивной сферы Pinus sylvestris L. в условиях среднерусской лесостепи (Воронежская область): автореф. дисс. ... канд. биол. наук / В.С. Свинцова. - Воронеж, 2002. - 22 с.
- Кузнецова, Н.Ф. Реакция на стресс и ее последействие у сосны обыкновенной в онтогенезе и при смене поколений / Н.Ф. Кузнецова, О.С. Машкина // Хвойные бореальной зоны. - 2011. - Т. 28. - № 1-2. - С. 83-90.
