Изменение оптических характеристик гидросферы в присутствии синтетически активных веществ

Автор: Захаров Валерий Павлович, Тимченко Елена Владимировна, Тимченко Павел Евгеньевич, Золотухина Александра Дмитриевна, Алембеков Сергей Валерьевич
Журнал: Компьютерная оптика @computer-optics
Рубрика: Дифракционная оптика, оптические технологии
Статья в выпуске: 2 т.35, 2011 года.
Бесплатный доступ
Представлены результаты экспериментальных исследований изменения спектральных характеристик водного растения - Элодеи бразильской (Elodea Brazilian, Egeria densa) под действием синтетических активных веществ и солей кадмия. В качестве критерия использован дифференциальный коэффициент обратного рассеяния, выражающий изменение концентрации хлорофилла в растении при воздействии поллютантов. Методом флуоресцентной конфокальной микроскопии проведён детальный анализ структурных изменений в клетках растения. Показано, что синтетические поверхностно-активные вещества (СПАВ) солюбилизируют белки мембран клетки, приводя к существенному изменению проницаемости и, как следствие, оптических характеристик биологической среды.
Метод дифференциального обратного рассеяния, конфокальная микроскопия, оптический коэффициент, хлорофилл, гидросфера, ионы кадмия, синтетические поверхностно-активные вещества
Короткий адрес: https://sciup.org/14059010
IDR: 14059010
Alteration of hydrosphere optical properties by synthetic active compounds
The results of experimental studies of spectral characteristics alteration of the water plant - Elodie Brazilian (Elodea Brazilian, Egeria densa) under the action of synthetic active compounds and salts of cadmium are submitted. The criterion used in The differential backscattering coefficient were used as criterion and expresses the changes in the concentration of chlorophyll in the plant under the influence of pollutants. The detailed analysis of structural changes in plant cells was performed by fluorescent confocal microscopy. It is shown that surfactants solubilize membrane-bound proteins, leading to a significant change in membrane permeability and, consequently the optical characteristics of the biological tissue.
Текст научной статьи Изменение оптических характеристик гидросферы в присутствии синтетически активных веществ
Увеличение объёмов активной добычи полезных ископаемых и химического производства привело к накоплению в гидросфере синтетических поверхностно-активных веществ (СПАВ) [1] и тяжёлых металлов [2], которые оказывают существенное влияние на функционирование живых систем гидросферы [3]. Чаще всего контроль содержания поллютантов в сточных водах осуществляется такими физическими методами анализа, как жидкостная тонкослойная хроматография [4] и полярография [5-6]. Данные методы контроля требуют достаточно больших временных затрат для получения требуемого результата, что ограничивает их применение только количественным статическим контролем величины концентрации поллютанта.
Известно, что выбросы СПАВ и солей тяжёлых металлов, как правило, носят залповый характер с последующим достаточно быстрым их аккумулированием гидросферой. Следовательно, эффективность выборочного концентрационного контроля гидросферы крайне низка – даже отсутствие поллютантов в водной среде на данной момент времени не может служить гарантией её нормального экологического состояния. Вместе с тем конечной целью контроля поллютантов является предотвращение их негативного влияния на живые системы гидросферы. Следовательно, принципиальным является, прежде всего, контроль состояния живых систем и его изменение под действием СПАВ. Ввиду способности аккумулировать различные типы поллютантов водные растения являются эффективными маркерами экологического состояния гидросферы, которые в отличие от традиционных способов измерения концентраций поллютантов выражают резистивные функции биологической среды на внешнее воздействие, включающее в себя множественные факторы и типы поллютантов.
Накопление поллютантов в водных растениях носит интегрирующий характер, т.е. фактически растения содержат в себе «историю» выбросов загрязнителей. Данные эффекты неизменно приводят к изменению концентрации хлорофилла a и b в растении, что, с одной стороны, меняет его биологический цикл, а с другой – приводит к изменению спектральных свойств среды. Следовательно, контролируя данные спектральные изменения, можно не только судить о количестве выбросов поллютантов, но и одновременно оценивать степень их влияния на гидросферу. Данный контроль может быть осуществлён методом дифференциального обратного рассеяния, который обладает высокой чувствительностью и даёт наиболее целостную информацию о состоянии живой системы [7 - 8].
Для понимания механизмов накопления и воздействия поллютантов на живые системы крайне важным является анализ изменений в структуре растения на клеточном уровне. Учитывая высокую эффективность флуоресценции хлорофилла, в настоящей работе для микроскопического анализа использован метод конфокальной флуоресцентной микроскопии [9], обеспечивающий контрастные микроснимки высокого разрешения.
Материалы и методы исследования
В качестве объекта исследования был использован пресноводный макрофит Элодея бразильская (Elodea Brazilian, Egeria densa). Растения были разделены на 6 групп. Контрольная группа растений находилась в среде фильтрованной водопроводной воды. Две опытные группы помещались в водные растворы соли нитрата кадмия с концентрациями 100 и 10 мг/моль соответственно, а другие три группы составляли водные растворы общедоступного СПАВ с концентрациями 0,2, 0,02 и 0,002%. Данные поллютанты вносились однократно в начале эксперимента, т.е. фактически ими- тировался однократный залповый выброс загрязнителей с фиксированной их концентрацией. Опыты проводились в лабораторных условиях в резервуарах с водой, исключающих внешние воздействия. Обеспечивались единые климатические условия для всех групп растений, а также регулярность и постоянство светового потока, воздействующего на среду.
Экспериментальный стенд, реализующий методы дифференциального обратного рассеяния и конфокальной микроскопии, представлен на рис. 1.
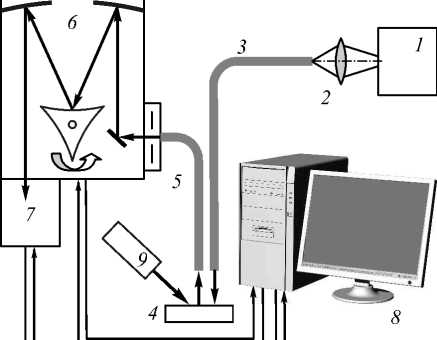
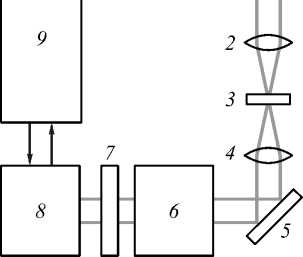
Рис. 1. Экспериментальный стенд:
а) спектрофотометрическая установка:
1 – источник света (галогеновая лампа),
2 – фокусирующая линза, 3 – подающее волокно,
4 – объект исследования, 5 – приёмное волокно,
6 – спектрограф SR-303i, 7 – цифровая камера ANDOR DV-420A-OE, 8 – компьютер,
б) установка флуоресцентной конфокальной микроскопии: 1 – источник видимого света (галогеновая лампа), 2 – коллиматор, 3 – объект, 4 – объектив, 5 – поворотное зеркало, 6 – конфокальный сканирующий блок, 7 – фильтры, 8 – камера, 9 – компьютер
Спектрофотометрическая установка (рис. 1а) включала в себя источник излучения, волоконную систему сбора и подачи излучения и спектрофотометр SR-303i (фокальная длина 303 мм) с интегрированной цифровой камерой ANDOR DV-420A-OE (1024x256, 26 мкм2, 16 бит). Экспериментальная установка позволяет работать с излучением в спектральном диапазоне 180^1200 нм с погрешностью регистрации 0,2 нм. В качестве источника излучения использовалась галогеновая лампа (спектральный диапазон излучения вырезался с помощью фильтров и составлял 400…1000 нм).
Установка флуоресцентной конфокальной микроскопии (рис. 1 б ) состояла из источника излучения (галогеновая лампа), оптической системы фокусировки, конфокального блока и камеры регистрации (1024 x 1024, время экспозиции - 40 мс).
Эксперименты проводились на протяжении всего жизненного цикла водоросли с ежедневным многократным контролем оптических характеристик растения и регистрацией микроснимков.
В качестве основного оптического параметра, характеризующего изменение концентрации хлорофилла при действии поллютантов, был использован дифференциальный оптический коэффициент ΔК, который был ранее применён в работе [7] для контроля атмосферных загрязнителей:
^ K =
K ( t ,0) - K ( t , C * )
K ( t ,0) ,
K = I 750
I 550
где C * – концентрация поллютанта, t – время, I 750 и I 550 – интенсивности обратного рассеяния на длинах волн 750 нм и 550 нм соответственно.
Дифференциальный оптический коэффициент Δ К является отношением экспериментально измеренного оптического коэффициента К к его значению в условиях отсутствия загрязнителей, т.е. соответствующих естественным условиям. Поскольку длина волны 550 нм соответствует максимуму поглощения хлорофилла, а длина волны 750 нм связана с поглощением клетчатки растения, то дифференциальный коэффициент Δ К однозначно отражает динамику изменения оптических характеристик растения вследствие влияния загрязнения среды.
Результаты исследований
Динамика дифференциального оптического коэффициента Δ К для растений различных групп (при различных концентраций СПАВ и солей кадмия) представлена на рис. 2.
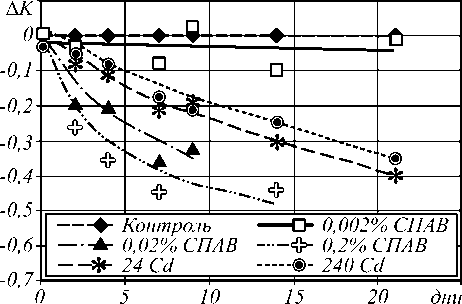
Рис. 2. Зависимость дифференциального оптического коэффициента от времени для растений различных групп
Видно, что в обоих случаях наблюдается спад дифференциального коэффициента вследствие деградации хлорофилла в биоткани. В то время как в контрольной группе растений значение коэффициента остаётся практически неизменным.
Первая группа растений, выращиваемых в среде с очень высокой концентрацией СПАВ (0,2%), к концу эксперимента практически полностью утрачивала пигменты, что отчётливо отражалось на изменении дифференциального оптического коэффициента (рис. 2).
Следует отметить, что, несмотря на общую тенденцию, функциональные зависимости изменения оптических характеристик среды при воздействии СПАВ и солей тяжёлых металлов качественно раз- личаются.
Если при добавлении СПАВ наступает насыщение (за характерный период времени ~10-15 дней – рис. 2), то в случае воздействия тяжёлых металлов динамика изменения дифференциального коэффициента имеет квазилинейный характер.
Данные качественные различия удалось понять, проведя микроскопический анализ растений на клеточном уровне с использованием метода конфокальной флуоресцентной микроскопии. Характерные микроснимки (180 x 110 мкм) ткани растений различных контрольных групп (10 день) представлены на рис. 3. Реализованная установка конфокальной микроскопии обеспечивает разрешение, достаточное для уверенной регистрации хлорофилла (рис. 3 г ), что позволяет рассчитать его концентрацию на клеточном уровне (рис. 3 д ).
в)
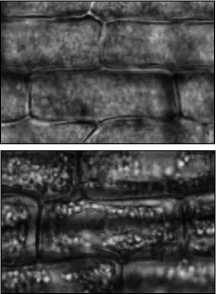
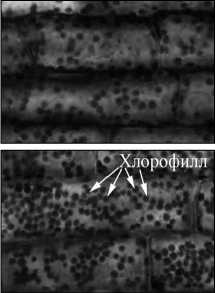
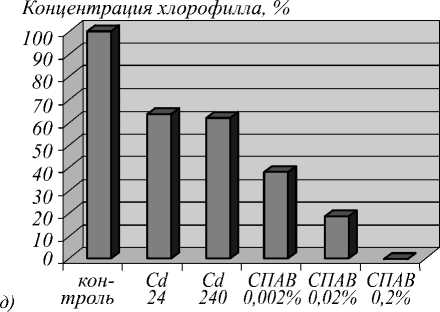
Рис. 3. Микроснимки (180 x 110 мкм) ткани растения при воздействии СПАВ 0,2% (а), СПАВ 0,002% (б), кадмия 100 мг/моль (в) и контрольной группы (г);относительная концентрация хлорофилла в группах растений (д)
Из микроскопического анализа следует, что в случае воздействия солями тяжёлых металлов идёт процесс «связывания» молекул хлорофилла, который микроскопически проявляется как их «слипание» вблизи мембран клеток (рис. 3 в ). При этом общее число молекул хлорофилла меняется незначительно, а вот неравномерность их распределения внутри клетки нарастает со временем. Причём эффективный объём, занимаемый хлорофиллом, уменьшается квазилинейно, приводя к аналогичной зависимости в изменении дифференциального оптического коэффициента. Напротив, в присутствии СПАВ сохраняется равномерность распределения хлорофилла по клетке (рис. 3 б ), но концентрация его падает с увеличением концентрации в залповом выбросе СПАВ, приводя к полной деградации хлорофилла при достижении концентрации СПАВ порядка десятых долей процента (рис. 3 а ).
Полученные результаты связаны с тем фактом, что СПАВ солюбилизируют мембраносвязанные белки мембраны клетки, нарушая её свойства [10]. Это способствует проникновению СПАВ во внутриклеточное пространство. Глубина действия настолько велика, что отражается даже на двумембранных органоидах, таких, как пластиды. Это способствует высвобождению пигментов из растения, тем самым снижая их концентрацию и приводя к изменению дифференциального оптического коэффициента. По мере увеличения доли СПАВ во внутриклеточном пространстве скорость данного процесса падает вследствие уменьшения хлорофилла, что проявляется в виде эффекта насыщения дифференциального оптического коэффициента.
Выводы
В результате проведённых исследований установлены функциональные зависимости оптического коэффициента от концентрации поллютантов (СПАВ и солей кадмия). Выявлено, что наибольшее влияние на изменение оптических характеристик живых систем в условиях гидросферы оказывают синтетические поверхностно-активные вещества.
Микроскопически доказано, что механизм деградации хлорофилла в присутствии СПАВ связан с солюбилизацией белков мембраны и изменением её проницаемости.
Работа выполнена при поддержке Федеральной целевой программы «Научные и научно-педагогические кадры инновационной России» (2009 - 2013гг.).
