Изменение параметров жизнеспособности гибридных особей, гетерозиготных по vestigial Drosophila melanogaster, в ходе длительной направленной селекции

Автор: Антосюк О. Н., Унжакова Д. В., Марвин Н. А.
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Биологические науки. Науки о Земле
Статья в выпуске: 2 т.25, 2022 года.
Бесплатный доступ
Изучение реализации наследственной изменчивости на фоне уже имеющихся мутантных аллелей определенных генов является важной частью биомедицинских и популяционных исследований, в том числе и человеческих заболеваний. В работе изучали изменение параметров жизнеспособности у особей дикого типа Белгород и у мутантной линии vestigial Drosophila melanogaster в процессе селекции. Регистрировали изменение параметров жизнеспособности в течение селекции у гибридных особей, гетерозиготных по vestigial, а именно изменение показателей плодовитости и эмбриональной летальности потомства у гибридных особей Белгород × vestigial в ходе направленной селекции, наличия регулируемой клеточной гибели в крыловом имагинальном диске у особей исходных линий Белгород и vestigial, у гибридных особей на начальном этапе селекции и спустя 30 поколений отбора. Изучались изменения формы крыла у межлинейных гибридных особей на разных этапах селекции в отношении особей дикого типа Белгород. Применялись методы исследования: селекция, изучение частоты гибели потомства на эмбриональной стадии развития, исследование фертильности, морфометрический анализ крыла, определение наличия и места локализации регулируемой клеточной гибели в крыловом имагинальном диске, статистический анализ. Выявлены маркерные параметры крыловой пластинки, чувствительные к присутствию мутантного аллеля vg в генотипе, не связанные с изменением интенсивности регулируемой клеточной гибели в крыловом имагинальном диске.
Vestigial, морфометрический анализ, крыловой имагинальный диск, селекция, вырезка на крыле, morphometric analysis, wing imaginal disc, breeding, cutting on wing
Короткий адрес: https://sciup.org/142231971
IDR: 142231971 | УДК: УДК 575.224 | DOI: 10.21443/1560-9278-2022-25-2-91-100
Текст статьи Изменение параметров жизнеспособности гибридных особей, гетерозиготных по vestigial Drosophila melanogaster, в ходе длительной направленной селекции
Антосюк О. Н. и др. Изменение параметров жизнеспособности гибридных особей, гетерозиготных по vestigial Drosophila melanogaster, в ходе длительной направленной селекции. Вестник МГТУ. 2022. Т. 25, № 2. С. 91–100. DOI:
Изменение частоты возникновения мутаций является одним из важнейших вопросов как популяционной, так и эволюционной генетики. Существует множество механизмов, отражающих это явление, например, изменение уровня метилирования, генная экспрессия и т. д. Для авторов наиболее интересно понятие "каскадный характер наследственной изменчивости", подразумевающее ускоренные темпы накопления мутаций, каждая из которых стимулирует появление все новых и новых мутаций. В таком случае вновь возникающие мутации выступают в качестве провокационного фона для того, чтобы запускать всплески мутационных частот ( Марвин и др., 2010 ). Используя линию vestigial (II, 67,0; vg), обладающую генотипом с присутствием данного аллеля в гомозиготной мутантной форме – выступающего в качестве провокационного фона, планируется проследить первичные изменения косвенных параметров жизнеспособности, так как наличие подобного аллеля вполне может запустить каскадный характер наследственной изменчивости в ходе направленной селекции именно на гетерозиготном состоянии vestigial. При этом присутствие мутантного аллеля в генотипе в гетерозиготном состоянии провоцирует возникновение мутации в дополнительных или определенных аллелях. Чаще всего спонтанные мутации, в том числе и в природе, затрагивают гены yellow , white и другие, связанные с инактивацией гена либо с состоянием псевдодоминирования при утрате аллеля на гомологичной хромосоме в процессе гаметогенеза самки ( Голубовский, 2011 ). Также может возникать состояние Beadex, когда появляется повреждение крыла по всему периметру крыловой пластинки, т. е. активная митотическая активность в крыловом имагинальном диске. Вместе с тем эти мутации могут возникать при измененном генотипе в гетерозиготной линии по vestigial ( vg ) друг за другом и накапливаются каскадом. Необходимо понять, как происходит становление измененного генотипа, что за этим следует и каким образом можно управлять этим процессом.
Использование мухи-дрозофилы или плодовой мушки ( Drosophila melanogaster ) в качестве модельного объекта для последующей экстраполяции полученных результатов на млекопитающих и человека вполне обоснованно, так как согласно литературным данным гены семейства Vestigial Like (VGLL) млекопитающих являются гомологами гена vestigial Drosophila melanogaster . Белки VGLL, в частности домен TONDU, участвуют во многих сигналинговых путях (например, Wnt-сигналинг, Hippo), а также являются супрессорами опухолей ( Simon et al., 2016 ).
У человека также может наблюдаться каскадный характер наследственной изменчивости. Некоторые мутации в аллелях у гетерозигот способны накапливаться и влиять на изменение частоты возникновения новых мутаций в популяциях, которые при этом становятся частью генетического груза, если они рецессивные, и в гетерозиготной форме остаются незамеченными. Эта скрытая наследственная изменчивость имеет негативное проявление, например, в виде наследственного заболевания. При определенных внешних воздействиях этот внутренний скрытый генотипический провокационный фон может проявить себя. В качестве факторов геномной дестабилизации может действовать не только направленная селекция, но и другие, такие как экология, продукты любого химического синтеза.
Цель работы – оценить влияние длительной направленной селекции на примере межлинейных гибридов Белгород × vestigial.
Материалы и методы
В качестве модельного объекта использовали D. melanogaster , которая является классическим объектом исследований.
В ходе исследования изучались следующие линии D. melanogaster :
-
1) vestigial – фенотипически отличается рудиментарными крыльями;
-
2) линия дикого типа Белгород, отловленная в окрестностях г. Белгорода в 2006 г. и инбредизированная в лабораторных условиях.
Гетерозиготные по vestigial гибридные особи характеризуются наличием вырезок крыловой пластинки, что позволяет проводить направленную селекцию гетерозиготных особей из поколения в поколение.
Все анализируемые линии содержались на традиционной питательной среде Альдерстона (250 мл воды, 2 г агара, 25 г глюкозы ч.д.а., 25 г дрожжей) в термостате при температуре +24 °С.
Для создания гибридного потомства виргинные самки линии Белгород отбирались не позднее 4 часов после вылета из пупария. После чего около 20 особей помещали в специальные колбы для массового разведения совместно с самцами линии vestigial.
На дне колбы устанавливали агаровые чашки Петри, обильно смазанные дрожжами. Спустя 72 часа чашки Петри извлекались, и полученное потомство переносили на питательную среду Альдерстона.
После вылета имаго гибридных особей F1 анализировали относительно морфологии крыловой пластинки. Селекцию вели по направлению двустороннего повреждения крыла типа "вырезка". Подобное ведение селекции способствует отбору именно гетерозиготных особей по vestigial.
В общей выборке потомства присутствуют следующие фенотипы (и соответственно генотипы) особей:
-
1) цельнокрылые (б/в) vg+/vg+ ;
-
2) с односторонним повреждением крыла (св 1) vg+/vg ;
-
3) с двусторонним повреждением крыла (св 2) vg+/vg ;
-
4) рудиментарнокрылые (vg) vg/vg .
Постоянное присутствие данных фенотипов в потомстве гетерозиготных особей свидетельствует о классическом расщеплении.
Оценка уровня регулируемой клеточной гибели (РКГ) в крыловом имагинальном диске (КИД) личинок проводится следующим образом:
-
I. Для анализа КИД извлекаются крыловые имагинальные диски из отобранных личинок D. melanogaster начала третьего возраста. В течение нескольких минут готовится прижизненный препарат органа в акридиновом оранжевом красителе в концентрации 0,005 % на физиологическом растворе Рингера для насекомых. Для каждой экспериментальной группы используется выборка из 25 КИД.
-
II. Препараты рассматриваются под флюоресцентным микроскопом Leica DM500 B на увеличении в 300 раз при ультрафиолетовом облучении препарата. Для анализа изображения фиксируются с помощью встроенной камеры. Определение РКГ проводится по таким признакам, как конденсация хроматина, уплотнение, изменение цитоплазмы.
-
III. КИД подразделяется на 4 сегмента (компартмента) ( Булгакова, 2003 ), которые обозначаются следующим образом (рис. 1):
-
1) задне-брюшной (PV или первый);
-
2) передне-брюшной (AV или второй);
-
3) задне-спинной (PD или третий);
-
4) передне-спинной (AD или четвертый).
Каждый компартмент оценивается полуколичественным методом со следующими рангами:
-
0 – отсутствие апоптоза;
-
1 – апоптоз от единичных проявлений до четверти площади компартмента органа;
-
2 – площадь фиксируемого апоптоза составляет от четверти до половины площади компартмента имагинального диска;
-
3 – от половины до трех четвертей площади компартмента диска заполнено апоптирующими клетками;
-
4 – наблюдается от трех четвертей до полного апоптотического повреждения компартмента диска.
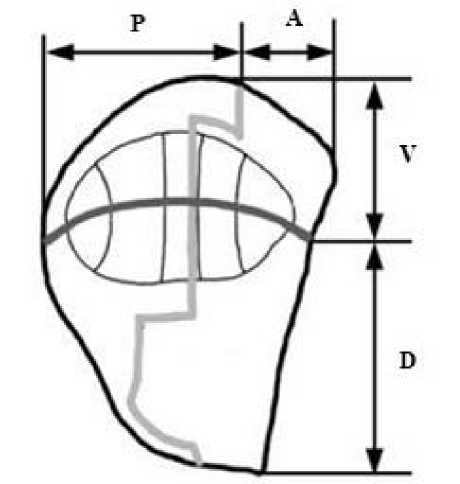
Рис. 1. Крыловой имагинальный диск и его компартменты ( Булгакова, 2003 ): Р – задний; А – передний; V – вентральный; D – дорсальный
Fig. 1. The wing imaginal disc and its compartments: P – posterior; A – anterior; V – ventral; D – dorsal ( Bulgakova, 2003 )
В процессе исследований использовались метод изучения фертильности и частоты гибели потомства на эмбриональной стадии развития и морфометрический анализ крыла.
Метод изучения фертильности и частоты гибели потомства на эмбриональной стадии развития (учет эмбриональных доминантных леталей)
Данный метод заключается в том, что на протяжении 10 дней учитывается индивидуальная плодовитость каждой пары особей в количестве 25 пар. На основании полученной плодовитoсти за весь период эксперимента высчитывается средняя индивидуальная плодовитость (СИП) каждой пары. Ежедневно собирали яйца, отложенные каждой парой за сутки, и помещали их на агаровые пластинки на смоченную водой фильтровальную бумагу в чашки Петри. Дополнительно определяли частоту гибели особей и их стерильность в течение всего периода эксперимента.
Доминантную эмбриональную летальность потомства учитывали как на ранней стадии развития (до 6 часа) РЭЛ (ранние эмбриональные летали), так и на поздней (после 6 часов) стадии ПЭЛ (поздние эмбриональные летали) эмбрионального этапа онтогeнеза.
Морфометрический анализ крыла
Изменение морфологии крыла определяли с использованием морфометрического анализа крыла по 24 показателям (18 линейных и 6 двумерных) (рис. 2).
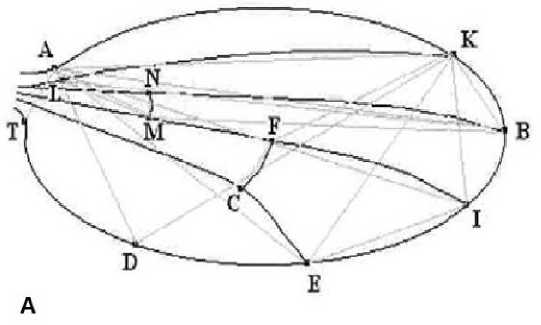
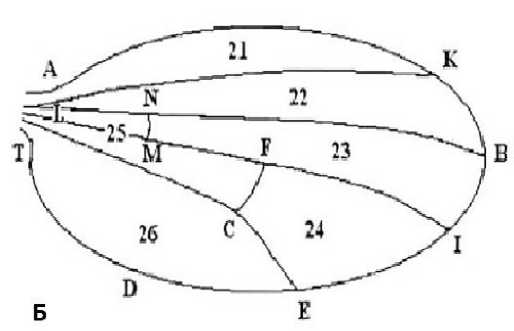
Рис. 2. Расположение точек, линейных ( a ) и двумерных параметров ( б ) на схеме крыла дрозофилы
Fig. 2. Location of points, linear ( a ) and two-dimensional parameters ( б ) on the drosophila wing diagram
Крылья особей фиксировали в 70%-м спирте на предметном стекле в виде временного препарата и фотографировали. Обрабатывали фотографии с помощью программы Universal Dekstop Ruler.
Статистический анализ проводился с использованием пакета программ STATISTICA 13. Для оценки результатов морфометрии использовали дискриминантный анализ. Для анализа плодовитости и летальности на эмбриональной стадии потомства применяли критерий Стьюдента.
Результаты и обсуждение
В ходе анализа особенностей гибридных мух Белгород × vestigial нами было обнаружено, что при скрещивании исходных линий Белгород и vestigial все особи F1 имели цельную крыловую пластинку, несмотря на то, что генотипически все мухи являлись vg+/vg. По нашему предположению, данное обстоятельство можно объяснить низким уровнем изменчивости исходной линии Белгород, что сказывается и на невыраженной изменчивости формы крыла. Вместе с тем, вероятно, восстановление целостной структуры крыла может происходить при достаточно низком уровне фермента гипоксантин-гуанинфосфорибозилтрансфераза (ГГФТ) у особей дикого типа Белгород. Согласно литературным данным количество данного фермента связано с фенотипом vg, вырезками при гетерозиготном генотипе и частоте появлений вырезок у особей дикого типа Белгород в норме с генотипом vg+/vg+ в естественных условиях. Хотя, по данным некоторых исследователей (Goux, 1973), уже в первом поколении наблюдаются повреждения крыла типа "вырезка" у нескольких линий дикого типа Белгород и лабораторных линий, послуживших материнскими при межлинейном скрещивании.
В ходе дальнейшей селекции во втором поколении согласно закономерностям наследования для моногибридного скрещивания появлялись особи с рудиментарнокрылым фенотипом (генотип vg / vg ), которые удалялись и не использовались для получения потомства. Среди цельнокрылых особей отчетливо выраженные гетерозиготные морфологические типы впервые появились в F4, что способствовало дальнейшей селекции исключительно только этих форм. Полученные особи с повреждениями крыла составили менее 1 % среди всех особей выборки четвертого поколения, что свидетельствует в пользу нашего предположения о низком содержании фермента ГГФТ.
При определении фертильности мух из исходных линий установили, что линия vg характеризуется низким уровнем плодовитости относительно линии дикого типа Белгород. Средний показатель СИП у линии vg составил 15,96, тогда как у линии Белгород – 25,72 (рис. 3). После скрещивания исходных линий полученные гибриды первого поколения демонстрируют так называемый эффект гетерозиса, проявляющийся в виде генеративной формы. Самки F 1 откладывают больше яиц как в сутки (от 339 до 1 199 яиц в день у гибридных особей F 1 , тогда как vestigial – от 18 до 248 яиц в сутки, линия Белгород демонстрирует диапазон 285–1 141 яйцо в сутки), так и согласно всему периоду эксперимента относительно исходных линий. За весь десятидневный период эксперимента особи линии Белгород суммарно отложили 5786 яиц, особи линии vestigial – всего 3 854 яйца, а гибридные особи первого поколения – 7 045 яиц. Средний показатель СИП линии Белгород × vestigial F1 достигает значения 28,18 и является математически значимо выше, чем тот же показатель у исходной линии дикого типа Белгород –25,72. В свою очередь, в ходе анализа плодовитости у гибридов спустя 10, 20 и 30 поколений отбора не обнаружено определенной тенденции в изменении уровня фертильности. На протяжении 30 поколений селекции наблюдали колебания в различных направлениях в количестве откладываемых яиц. Сходный эффект гетерозиса не наблюдается при дальнейшей селекции у гибридов F 10 , F 20 и F 30 , что в данном случае согласуется с эффектом затухания. Наблюдаемые колебания уровня фертильности могут являться свидетельством того, что полученная гибридная линия находится в дестабилизированном состоянии под действием селекции и в результате исходного аутбредного скрещивания стала еще более гетерогенной. Гетерогенность также выражается в увеличении коридора колебания на графике в отношении СИП у особей F 1 .
Экспериментальные линии D. melanogaster
Рис. 3. Средняя индивидуальная плодовитость экспериментальных линий D. melanogaster
Fig. 3. Average individual fertility of the experimental group of D. melanogaster
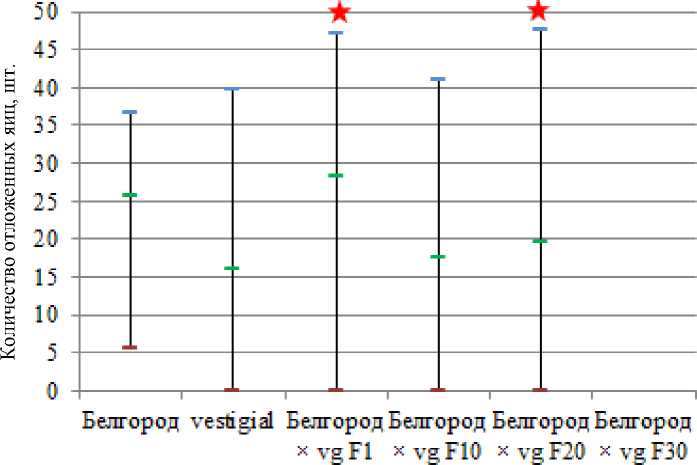
При анализе гибели потомства на эмбриональной стадии обнаружили связь присутствия алелля vg в гомозиготной мутантной форме и высокого уровня летальности потомства на самых ранних этапах эмбриогенеза (РЭЛ, до 6 часа развития). Коридоры колебания значений РЭЛ не перекрываются у мутантной линии vestigial и линии дикого типа Белгород, потому что показатель РЭЛ у особей линии vestigial не опускается ниже значения 20,77 % (рис. 4). Эффект гетерозиса у гибридов F 1 мы наблюдали и в отношении данного показателя. В ходе же направленной селекции наблюдается тенденция к восстановлению уровня летальности на раннем этапе эмбрионального развития у потомства до наблюдаемого среднего значения 35,97 % у линии vestigial. Согласно статистическому анализу уже в 20-м поколении СИП не отличается от значений линии vestigial, и в поколении F30 среднее значение СИП, равное 28,69, становится еще более близким к среднему значению СИП vestigial – 35,97. Подобное явление вполне может быть связано с тем, что в ходе селекции на гетерозиготность по vestigial неизбежно идет неосознанный отбор других генов, тесно связанных в своем функционировании с vestigial напрямую и косвенно, например генов scalloped , decapentaplegic и др. Известно, что продукт гена scalloped димеризует белок vestigial для попадания последнего в ядро, к тому же именно димерный комплекс scalloped – vestigial является транскрипционным фактором, активно влияющим на пролиферацию клеток ( Halder et al., 1998 ). В этой связи крайне важен генотип особей, подвергающихся селекции в отношении гена scalloped , мутантные проявления которого фенотипически идентичны гетерозиготному состоянию vg+/vg .
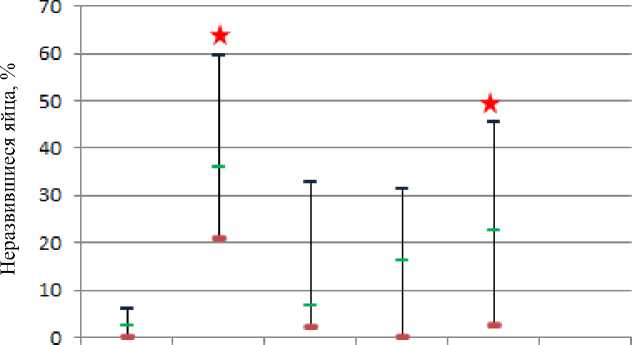
Белгород vestigial Белгород Белгород Белгород Белгород xvgFl xygFlD vgF20 xVgF50
Экспериментальные линии D. melanogaster
Рис. 4. Частота ранней эмбриональной летальности потомства F1 экспериментальных групп D. melanogaster
Fig. 4. The frequency of early embryonic lethality of the experimental group of D. melanogaster
Нижняя граница коридора колебания значений РЭЛ в группах F 20 и F 30 не достигает нулевой отметки, что также наблюдается и в группе vestigial.
Поздняя эмбриональная летальность реализуется после 6-го часа развития. В группе гибридных особей F 1 наблюдали наиболее высокий показатель частоты летальности потомства на поздней стадии эмбрионального развития из всех анализируемых экспериментальных групп. Среднее значение в данной группе также является максимальным (2,69 %) из всех анализируемых выборок. Подобная картина может наблюдаться в соответствии с негативным проявлением эффекта гетерозиса в связи с высокой гетерогенностью выборки.
Высокой частоты летальности потомства на поздней стадии развития не наблюдается ни у одной из исходных линий, она стабилизируется уже в 10-м поколении отбора и не достигает тех значений среднего, как наблюдается в группе из поколения F 1 . Повышение гетерозиготности может приводить к такого рода эффектам благодаря наличию в большем количестве доминантных аллелей генов, реализующихся по типу межаллельных взаимодействий, в том числе комплементарно.
Анализируя имагинальные крыловые диски личинок начала третьего личиночного возраста у следующих групп: Белгород, vestigial, Белгород × vestigial F1, Белгород × vestigial F30 обнаружили, что у исходных линий РКГ не встречается в нотумной части, а ведь по литературным данным именно там наблюдается самая активная пролиферация в этот интервал личиночного развития (Lawrence et al., 1977). Сравнительный анализ локализации РКГ среди дисков экспериментальных групп показал, что наиболее распространенной является кромка РКГ по внешней стороне ткани диска, редко встречаются центральные очаги, по большей части – точечного характера (рис. 5).
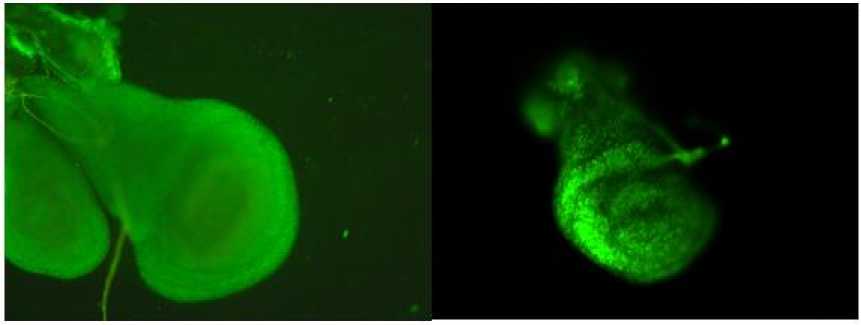
Рис. 5. Крыловые имагинальные диски особей линии Белгород без РКГ (слева) и с РКГ (справа) Fig. 5. Wing imaginal discs of individuals of the Belgorod line without RCD (left) and with RCD (right)
Нами было установлено, что согласно теории доминирования у гетерозигот по vg процессы вовлечения в РКГ нотумных областей также реализуются на этапе третьего личиночного возраста. Это может быть связано с гетерогенностью гибридов, так как, ведя селекцию на гетерозиготность по vestigial, мы отбираем и ряд других генов, играющих ключевую роль в процессах клеточного деления и его контроля. Это может быть и негативная роль, в том числе за счет комплементарного взаимодействия межгенных продуктов. Негативные эффекты определяются по следующим показателям: во-первых, обнаружена тенденция в восстановлении исходного по vestigial показателя РЭЛ в ходе селекции, во-вторых, нотумная часть у гибридов включена в РКГ процесс, более того, выяснилось, что данный аллель vg не влияет на апоптоз на этом этапе. В литературе описано отсутствие влияния на интенсивность РКГ определенных форм гена vg при использовании окраской акридиновым оранжевым красителем ( Van de Bor et al., 1999 ). Авторами показано, что только мутантный белок vestigial с определенной усеченной морфологией способствует индукции гибели клеток. К тому же vestigial как ген с плейотропным эффектом может вызывать гибель клеток в одних дисках, но при этом не инициировать РКГ в других, действуя таким же образом в отношении центральной нервной системы и грудных мускулов и т. д. ( Garg et al., 2007 ).
В ходе анализа изменения формы крыловой пластинки межлинейных гибридных особей Белгород × vestigial в ходе направленной селекции провели сравнение крыльев у особей следующих экспериментальных групп: исходной линии дикого типа Белгород, межлинейных гибридных особей Белгород × vestigial F 1 , F 20 и F 30 . Согласно сравнению морфометрических линейных параметров не дискриминирующими являются только 5 из 18 параметров (рис. 6).

Рис. 6. Недискриминирующие крыловые линейные параметры у экспериментальных групп D. melanogaster Fig. 6. Non-discriminating wing linear parameters in experimental groups of D. melanogaster
Параметры AI, FC, NM, KB, KF затрагивают центральную область крыловой пластинки и самую вариабельную область апикальной части крыла. Именно апекс крыла наиболее подвержен морфологическому изменению у гетерозиготных особей по vestigial за счет наличия разных по размеру повреждений типа "вырезка" на крыле.
При анализе двумерных параметров крыла (площади отдельных ячеек крыловой пластинки) обнаружили, что все 6 областей ячеек крыла дискриминируют у анализируемых выборок. Согласно этим результатам можно предположить, что меняется не столько сама форма крыловой пластинки, сколько ее размер (табл.).
Таблица. Средние показатели площадей ячеек крыла у особей экспериментальных групп D. melanogaster , см Table. Average values of wing cell areas in individuals of the experimental groups of D. melanogaster , cm
|
Линия / Ячейки |
21 |
22 |
23 |
24 |
25 |
26 |
Общая |
|
Белгород |
0,22 |
0,33 |
0,35 |
0,2 |
0,35 |
0,49 |
1,94 |
|
Белгород × vestigial F 10 |
0,26 |
0,35 |
0,4 |
0,39 |
0,2 |
0,48 |
2,08 |
|
Белгород × vestigial F 20 |
0,26 |
0,37 |
0,4 |
0,42 |
0,2 |
0,49 |
2,13 |
|
Белгород × vestigial F30 |
0,17 |
0,24 |
0,28 |
0,29 |
0,13 |
0,34 |
1,44 |
Нами было высказано мнение, что в ходе направленной селекции идет увеличение размеров крыловой пластинки относительно исходной линии дикого типа Белгород, но в течение селекции к 30-му поколению отбора процесс меняется на противоположный: размер крыловой пластинки резко уменьшается, становясь почти по всем средним показателям площадей (за исключением 24) меньше, чем даже у линии дикого типа. По нашему предположению, подобная картина может наблюдаться при некоторых генотипических комбинациях генов, включенных в процессы клеточной пролиферации и контролирующих дифференцировку ткани крылового имагинального диска. Также стоит отметить, что прирост размера крыла осуществляется за счет обратно пропорциональных изменений в площади 24 и 25 ячеек крыла и незначительного увеличения площади 21 ячейки.
В ходе канонического анализа линейных морфометрических показателей крыла выборок исходной линии Белгород и межлинейных гибридов в ряду поколений определили различия в форме крыла у гибридных особей и исходной линии. Крылья межлинейных гибридных особей разных поколений селекции также меняются с течением селекционного процесса, что отображено на рис. 7.
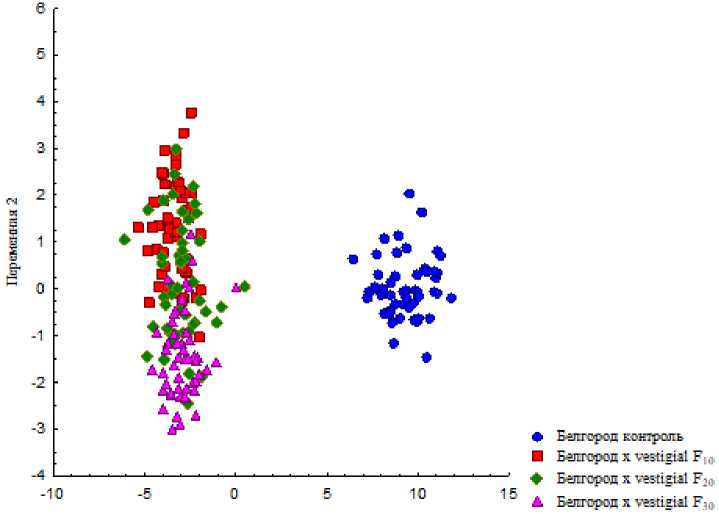
Переменная 1
Рис. 7. Канонический анализ линейных морфометрических показателей крыла выборок линии Белгород и межлинейных гибридных линий Белгород × vestigial F 10 , F 20 и F 30 Fig. 7. Canonical analysis of linear morphometric parameters of the wing samples of the Belgorod line and interlinear hybrid lines Belgorod × vestigial F10, F20, and F30
В отношении канонического анализа двумерных показателей крыла у экспериментальных групп, связанных с линией Белгород, выявили схожую тенденцию в различиях морфологии крыла линии дикого типа и межлинейных гибридных особей, но с менее выраженной разницей изменения формы крыла у гибридных линий разных поколений. В связи с этим можно предположить, что направленная селекция на повреждение крыла типа "вырезка" эффективнее сказывается на изменении формы крыла, чем пропорций ячеек.
Заключение
В результате исследования получены данные по жизнеспособности гибридных линий Drosophila melanogaster в ходе направленной селекции в присутствии мутантного аллеля vestigial, который способствует снижению фертильности и повышению частоты доминантной эмбриональной летальности потомства на ранних этапах развития. Также его присутствие вызывает изменение формы крыла у гибридных особей Белгород × vg по отношению к форме крыла особей линии дикого типа Белгород как в отношении линейных, так и двумерных параметров крыловой пластинки. При этом уровень регулируемой клеточной гибели в крыловых имагинальных дисках личинок третьего возраста линии vestigial и гибридных особей Белгород × vg F 1 и F 30 не отличается от уровня клеточной гибели, наблюдаемого у особей линии дикого типа Белгород. В этой связи можно сделать вывод, что ген vestigial не участвует в изменении формы крыла через повышение уровня регулируемой клеточной гибели.
Работа выполнена при финансовой поддержке: постановление № 211 Правительства Российской Федерации, контракт № 02.A03.21.0006.
Авторы благодарят сотрудников кафедры биоразнообразия и биоэкологии института естественных наук и математики Уральского федерального университета имени первого Президента России Б. Н. Ельцина О. С. Павлову и В. Л. Вершинина за оказанную помощь при написании статьи.
