Изменение состава эфирного масла и уровня салициловой кислоты у растений Mentha piperita L. в онтогенезе (вторичные метаболиты в онтогенезе мяты)
")
Автор: Шелепова Ольга Владимировна, Кондратьева Вера Валентиновна, Воронкова Татьяна Владимировна, Олехнович Людмила Серегеевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
Изучали состав эфирного масла (ЭМ) и содержание салициловой кислоты (СК) в листьях и соцветиях клона Mentha piperita L. и двух сортов, выведенных на основе M. рiperita (Краснодарская 2 и Митчамская), в вегетативную фазу, фазы цветения и отцветания. Установлено, что выход ЭМ наибольший в фазу цветения, а уровень СК достигает максимума при отцветании. Выявлено сопряженное изменение ряда компонентов ЭМ, обладающих протекторным действием, и СК в процессе онтогенеза при воздействии природных биотических (тля, листогрызущие насекомые, грибные инфекции) и абиотических (температура, влага) стрессах.
Эфирное масло, салициловая кислота, стресс, ментол, ментофуран, сесквитерпен
Короткий адрес: https://sciup.org/148202033
IDR: 148202033 | УДК: 581.135.51-292.94
Changes in the essential oil levels and salicylic acid levels in plants Mentha piperita L. in the ontogeny (secondary metabolitas in the mint ontogeny)
Composition of essential oils and content of salicylic acid in the leaves and inflorescences of the Mentha piperita clone, and the two varieties, produced on the basis of M. piperita, Krasnodarskaya 2 and Mitchamskaya, in the vegetative phase, the phase of flowering and in the end of flowering was studied. Found that the yield of essential oils in the vegetative and flowering phases was the most, and salicylic acid reaches a maximum level after flowering. However, revealed the dual change some components of essential oils, having a protective effect, and the salicylic acid during ontogeny under the influence of natural biotic (aphids, leaf-eating insects, fungal infection) and abiotic (temperature, moisture) stress.
Текст научной статьи Изменение состава эфирного масла и уровня салициловой кислоты у растений Mentha piperita L. в онтогенезе (вторичные метаболиты в онтогенезе мяты)
Для эфирного масла растений рода Mentha L. характерно наличие органических летучих компонентов, то есть веществ, легко переходящих в газообразное состояние при изменении внешних условий и создающих особую атмосферу вокруг растений [5]. Физиологические функции этих веществ весьма разнообразны. Они связаны с защитой растений от биотических и абиотических стрессов, аллелопатическим взаимодействием растений, привлечением или отпугиванием насекомых. В состав этой группы входят многие терпеноиды, в первую очередь монотерпены и сесквитерпены, жирные кислоты, производные салициловой кислоты и ряд других компонентов [2]. На интенсивность эмиссии летучих веществ, особенно из листьев, сильно влияют абиотические факторы внешней среды, в первую очередь температура воздуха и почвы. Это можно рассматривать как часть программного ответа на нейтрализацию негативных последствий стресса. Испарение летучих компонентов масла корректирует влажность и температуру, стабилизирует среду, окружающую растение, нормализует обмен веществ [6]. Одним из индукторов устойчивости растений к различным стрессам выступает салициловая кислота (СК). Ее уровень в растениях, подверженных стрессу, повышается и сопряжен с содержанием активного кислорода [7]. СК является частью пускового механизма неспецифической антиоксидантной защиты.
В экспериментальных работах есть данные, указывающие на связь уровня эндогенной СК и экзогенных обработок этим веществом с выходом и качественным составом эфирного масла [3,4].
Целью представленной работы было выявление взаимосвязи уровня СК и количества эфирного масла, выделяемого листьями и соцветиями M. piperita , а также изменение содержания компонентов этого масла в процессе онтогенеза растения.
МАТЕРИАЛ И МЕТОДЫ
Объектами исследования были взяты клон M. piperita , посадочный материал получен из Никитского ботанического сада (Украина, Крым), и два сорта, выведенные на основе M. piperita – Краснодарская 2 и Митчамская, более двадцати лет выращиваемые в условиях средней полосы России.
Для анализа брали листья и соцветия растений второго года вегетации в фазе вегетативного роста, массового цветения и отцветания. Эфирное масло получали методом гидродистилляции по Гинзбегру [1]. Качественный анализ эфирного масла проводили на базе сервисной лаборатории комплексного анализа химических соединений кафедры физической и органической химии РГАУ-МСХА им. К.А. Тимирязева методом хроматомасс-спектрометрии на аналитическом комплексе “Clarus 600M” фирмы “Perkin Elmer”. Для определения свободной СК применяли метод ВЭЖХ на изократической системе приборов “Стайер”, фирмы Аквилон.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Динамика общего содержания эфирного масла в процентах от сырого веса одинакова у обоих сортов и клона M. piperita . Как в листьях, так и в соцветиях максимум приходится на фазу цветения. В эту фазу на 1 см² нижней поверхности листьев, закончивших формирование, расположено в 4-7 раз больше железистых образований, чем на листьях, сформировавшихся в вегетативную фазу развития растений (рис. 1).
Кроме того, секреторные железы образуются на лепестках и чашелистиках цветков. Диаметр железок в фазу цветения также увеличивается. В фазу отцветания количество железок и их диаметр имеют тенденцию к увеличению, но эти изменения не столь существенны.
Однако объем эфирного масла в процентах к сырому весу сокращается, но уровень основного мажорного компонента эфирного масла M. piperita , монотерпеноида ментола возрастает до максимума на стадии отцветания (рис. 2А, табл.). Возможно, это связано с метаболизмом ментона, максимальное содержание которого в листьях отмечено на стадии вегетатив-
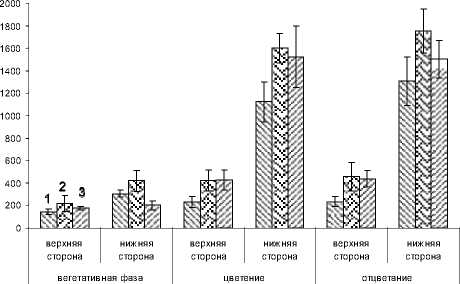
Рис. 1 . Количество желез на верхней и нижней сторонах M. piperita по фазам онтогенеза, шт/см2. 1- сорт Краснодар-ская2, 2- сорт Митчамская, 3- клон M. piperita (Крым)
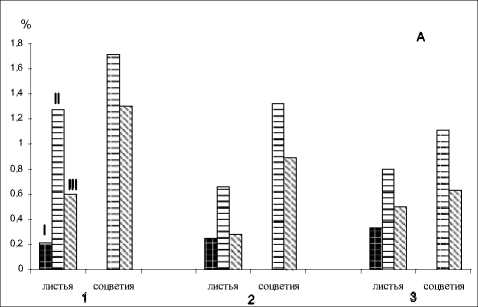
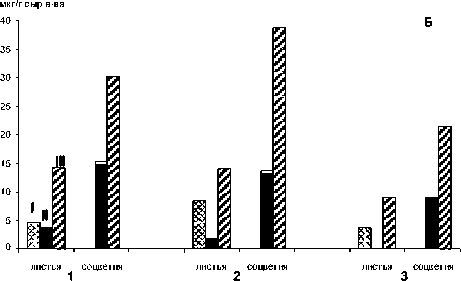
Рис. 2. А: Продуцирование эфирного масла органами M. piperita по фазам онтогенеза, % сырого веса. Б: Содержание салициловой кислоты в тканях органов M. piperita, мкг/г сырого веса. 1 - сорт Митчамская;
2 - сорт Краснодарская 2; 3 - клон M. piperita (Крым); I- вегетативная фаза, II- массовое цветение, Ш- отцветание
ного развития, а у соцветий - в начале цветения. В листьях и соцветиях на стадии отцветания его уровень 7-10 раз ниже. В соцветиях увеличение процентного содержания ментола в фазу отцветания может возрастать также в связи с превращением в него мажорного монотерпеноида пулегона, максимум которого в эфирном масле характерен для стадии цветения. В листьях этот терпеноид появляется только в фазу цветения в минорных количествах.
Следует отметить, что пулегон при стрессовых внешних воздействиях, а также при старении растительных тканей может превращаться в ментофуран. Возможно, увеличение его синтеза из пулегона в это
Таблица . Состав эфирного масла M. piperita по фазам онтогенеза, %
время связано со стабилизацией окружающей растение атмосферы и старением листьев. Так, у клона M. piperita ментофуран зафиксирован в листьях только в стадию отцветания. В день взятия проб при массовом цветении мяты была жаркая сухая погода (температура 25-27°С, относительная влажность 46-49%). В это время у сорта Митчамская отмечено максимальное содержание ментофурана. В листьях обоих сортов при отцветании возрастает уровень минорных сесквитерпенов, с фунгицидными свойствами - в—бурбонена и в—кариофилена. У клона их уровень почти не меняет -ся. В соцветиях всех изученных растений в-бурбонена не обнаружено, а уровень в-кариофилена при отцветании у клона M. piperita и сорта Митчамская снижается. По визуальным наблюдениям установлено, что в период отцветания на растениях мяты всех сортов и клонов появляется большое число листогрызущих насекомых, тли, грибной инфекции, а также насекомых, повреждающих формирующиеся семена. В составе эфирного масла в это время в листьях и соцветиях клона M. piperita , и листьях сортов, увеличивается уровень 1,8-цинеола, обладающего инсектицидным действием (табл.). Таким образом, на выход эфирного масла и его качественный состав влияют не только стадии онтогенеза, но и условия окружающей среды.
Наряду с терпенами, важную роль в регуляции продуцирования и состава эфирного масла играют салицилаты, которые через серию каскадных биохимических реакций активизируют гены протекторных механизмов. В итоге обеспечивается оптимальный окислительно-восстановительный гомеостаз клеток, транспорт ионов через плазмолемму, интенсивность фотосинтетических процессов [3]. У обоих сортов и клона M. piperita наибольшее содержание СК, как и выход эфирного масла, отмечено в тканях соцветий, то есть, органов, наиболее важных для формирования семян и возобновления растений (рис. 2Б). В тканях листьев уровень СК во все три стадии онтогенеза мяты был ниже, чем в соцветиях, также как и продуцирование эфирного масла. Следует отметить, что у сортов разница была более существенной, чем у клона M. piperita. Как отмечено выше, на стадии отцветания, когда активизируются биогенные стрессовые факто- ры, возрастает содержание отдельных компонентов эфирного масла, обладающих инсектицидным и фунгицидным действием. Наряду с этим, и в листьях, и в соцветиях всех изученных растений M. piperita в стадию отцветания отмечен максимальный уровень иммуномодулятора СК. Наибольшее содержание СК зафиксировано в тканях соцветий сортов Митчамская и Краснодарская 2.
Таким образом, установлено, что существует сопряженное изменение уровня индуктора устойчивости растений к стрессам, салициловой кислоты, и ряда компонентов эфирного масла, обладающих бактерицидными, фунгицидными и инсектицидными свойствами при воздействии на растение негативных биогенных и абиогенных факторов (температура, влажность воздуха, патогенные грибы и насекомые вредители).
Работа выполнена при частичной поддержке гранта РФФИ 11-04-01820.
|
Сорт |
клон M.piperita (Крым) |
M.piperita, сорт Митчамская |
M.piperita, сорт Краснодарская 2 |
||||||
|
Фаза онтогенеза |
s |
s S |
s S о |
s O’ |
s s |
s S о |
s O’ |
s s |
S |
|
листья |
|||||||||
|
1,8-цинеол |
2,12 |
6,07 |
7,49 |
4 |
0,12 |
6,57 |
3,3 |
3,43 |
4,1 |
|
ментон |
24,49 |
9,93 |
2,48 |
38,03 |
8,34 |
2,88 |
35,92 |
12,61 |
5,34 |
|
ментофуран |
- |
- |
0,68 |
0,37 |
1,05 |
0,4 |
- |
- |
- |
|
ментол |
20,59 |
65,31 |
68,85 |
40,65 |
67,44 |
66,91 |
39,86 |
34,88 |
36,65 |
|
пулегон |
- |
0,2 |
- |
- |
0,56 |
- |
- |
0,78 |
0,36 |
|
β-кариофилен |
1,37 |
1,29 |
1,02 |
1,35 |
0,88 |
1,3 |
1,44 |
0,85 |
1,44 |
|
β-бурбонен |
0,25 |
0,21 |
0,2 |
0,25 |
0,15 |
0,32 |
0,39 |
0,12 |
0,37 |
|
соцветия |
|||||||||
|
1,8-цинеол |
1,88 |
2,65 |
2,73 |
1,93 |
1,1 |
1,2 |
|||
|
ментон |
22,16 |
9,01 |
22,29 |
9,17 |
15,01 |
15,99 |
|||
|
ментофуран |
34,48 |
29,57 |
26,58 |
24,21 |
15,38 |
15,1 |
|||
|
ментол |
14,07 |
42,93 |
23,23 |
48,75 |
- |
26,22 |
|||
|
пулегон |
18,13 |
3,33 |
12,07 |
2,58 |
42,49 |
7,65 |
|||
|
β-кариофилен |
0,61 |
0,46 |
0,98 |
0,6 |
0,37 |
0,5 |
|||
|
β-бурбонен |
- |
- |
- |
- |
- |
- |
|||
Список литературы Изменение состава эфирного масла и уровня салициловой кислоты у растений Mentha piperita L. в онтогенезе (вторичные метаболиты в онтогенезе мяты)
- Гинзберг А.С. Упрощенный способ определения эфирного масла в эфироносах//Хим.-фарм. промышленность. 1932. № 8 (9). С. 326-329.
- Holopainen J.K., Gershenzon J. Multiple stress factors and the emission of plant VOC's//Trends Plant Sci. 2010. V. 15. N 3. P. 176-184.
- Mateo A., Funck D., Mühlenbock P. et al. Controlled levels of salicylic acid are required for optimal photosynthesis and redox gomeostasis//J. Exp. Bot. 2006. V. 57. N 8. P. 1795-1807.
- Rowshan V., Khosh M., Javidnia K. Effects of salicylic acid and quantity of essential oil components in Salvia macrosiphon//J. Biol. Environ. Sci. 2010. N 4 (11). P. 77-82.
- Spinelli F., Cellini A., Marchetti L. Emission and function of volatile organic compounds in response to abiotic stress//Abiotic stress in plants -mechanisms and adaptations/Eds Shanker A.K., Venkaterswarlu B. Rijeka: InTech, 2011. P. 367-394.
- Stevović S., Ćalić-Dragosavac D., Mikovilović V. et al. Correlation between environment and essential oil in medical plants//Adv. Environmental Biol. 2011. V. 5. N 2. P. 465-468.
- Yang Y., Qi M., Mei C. Endogenous salicylic acid protects rice plants from oxidative damage caused by aging as well as biotic and abiotic stress//Plant J. 2004. V. 40. N 6. P. 909-919.
