Изменение жирнокислотного состава в растениях при гипотермической адаптации

Автор: Живетьев М.А., Граскова И.А., Дударева Л.В., Столбикова А.В., Войников В.К.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 4 т.6, 2010 года.
Бесплатный доступ
Исследование межвидовой изменчивости состава жирных кислот в тканях листьев трех видов лекарственных растений, произрастающих в условиях Предбайкалья, показало, что этот состав имеет видоспецифичный характер. В составе ненасыщенных жирных кислот всех изученных видов преобладают полиненасыщенные жирные кислоты - линолевая и леноленовая, что обуславливает высокие индексы двойных связей (до 2,19 у тысячелистника). Наряду с высокими значениями коэффициента ненасыщенности (до 4,7 у тысячелистника) этот факт свидетельствует о высоком потенциале холодоустойчивости изучаемых видов. Анализ сезонных изменений в составе жирных кислот трех видов лекарственных растений показал, что по мере снижения среднесуточной температуры происходит закономерное снижение содержания насыщенных и увеличение ненасыщенных жирных кислот. Установлено, что изменения в жирнокислотном составе липидов и уровне активности 9- и 3-десатураз в тканях листьев при адаптации к понижению среднесуточной температуры воздуха у исследуемых растений также имеют межвидовые отличия. Полученные данные дают основание полагать, что высокий потенциал холодоустойчивости у изучаемых видов связан, среди прочего, с особенностями процессов биосинтеза жирных кислот в составе их липидов.
Низкотемпературный стресс, жирные кислоты, активность ацил-липид- ных десатураз
Короткий адрес: https://sciup.org/14323502
IDR: 14323502
Текст научной статьи Изменение жирнокислотного состава в растениях при гипотермической адаптации
Проблема адаптации растений к экологическое значение, т. к. способность низкотемпературному стрессу имеет большое растений адаптироваться к конкретным условиям – это один из факторов, определяющих ареалы распространения диких видов и возможность их интродукции (Колесниченко, Войников 2003). Считается, что липиды клеточных мембран играют ключевую роль в процессах адаптации и формировании устойчивости растений к неблагоприятным факторам внешней среды, прежде всего к холоду. Эта устойчивость коррелирует с наличием в клеточных мембранах полиненасыщенных жирных кислот (ЖК) (Лось, 2005). Известно, что именно ненасыщенные жирные кислоты (НН ЖК) в структуре мембран определяют ее текучесть и сохранение жидкостных свойств при данных температурах. В существующей модели молекулярного механизма адаптации клеток к гипотермии первичная роль отводится мембранным липидам, в частности их способности к фазовым переходам в зависимости от температуры окружающей среды, и ферментам десатуразам, катализирующим синтез НН ЖК. В соответствии с этой моделью при снижении температуры уменьшается текучесть мембран, это приводит к усиленному синтезу десатураз в клетке, их активации и, как следствие, к ускорению синтеза полиненасыщенных ЖК в мембранных липидах (Ильинская, Озерецкая, 1998). В результате этих процессов текучесть мембран восстанавливается. Таким образом, именно активность десатураз является одним из ключевых моментов в адаптации растений к низким температурам (Лось, 2001).
Известно, что у высших растений существуют значительные межвидовые различия по холодоустойчивости. Этим объясняется особый интерес к подробному изучению механизмов низкотемпературной адаптации у растений, произрастающих в суровых климатических условиях. Логично предположить, что анализ жирнокислотного состава суммарных липидов из тканей исследуемых растений, а также оценка активности десатураз при действии низких температур будет способствовать лучшему пониманию их роли в формировании устойчивости растений к этому фактору. Несмотря на изученность биохимических процессов, обеспечивающих адаптивные изменения липидного состава мембран отдельных видов культурных растений, таких как арабидопсис, табак, кукуруза, соя, рис и других при действии низких температур, вопросы устойчивости и липидного метаболизма растений остаются мало исследованными. Кроме того, если по химическому составу эфирных масел, алкалоидов, дубильных веществ изучаемых растений имеются некоторые сведения, то их жирнокислотный состав практически не исследовался. По-этому целью нашей работы было изучить состав ЖК пяти видов лекарственных растений Прибайкалья, активность ферментов, влияющих на взаимопревращения жирных кислот, количественно преобладающих в листьях этих растений, и проследить сезонные изменения в метаболизме ЖК растений.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом исследования служили листья 3 видов лекарственных растений из флоры Предбайкалья (вероника дубравная - Veronica chamardrys L., манжетка обыкновенная -Alchemilla vulgaris complex, тысячелистник обыкновенный - Achillea milleforum L., одуванчик лекарственный - Taraxacum officinale, подорожник большой - Plantago major L.), собранных на левом берегу реки Выдринная, 600 м от уреза оз. Байкал, стационар СИФБРа, и в г. Иркутске, на территории СИФИБРа. Образцы для жирнокислотного анализа фиксировали и растирали в жидком азоте до получения гомогенной массы для экстракции липидов. Экстракцию липидов из тканей исследуемых объектов проводили с использованием системы растворителей хлороформ-метанол-вода (1:2:0,8 v/v/v). Для удаления хлороформа из экстракта липидов использовали роторный испаритель RVO-64 (Чехия). Для получения метиловых эфиров жирных кислот к экстракту липидов после удаления растворителя добавляли 1% метанольный раствор Н2SO4 и нагревали на водяной бане при 600С в течение 30 мин. После охлаждения метиловые эфиры жирных кислот трижды экстрагировали гексаном. Анализ полученных метиловых эфиров жирных кислот проводили методом газожидкостной хроматографии с использованием хромато-масс-спектрометра 5973N/6890N MSD/DS Agilent Technology. Капиллярная колонка HP-INNOWAX (30м х 250мк х 0.50 мм), градиент температуры: от 100ºС до 150ºС со скоростью 10ºС в мин от 150ºС до 255ºС со скоростью 3ºС в мин. Для расчета эквивалентной длины цепи использовали изократический режим, температура колонки 200ºС. Газ-носитель гелий, скорость потока газа 1 мл/мин. Масс-спектрометр - квадруполь, способ ионизации -электронный удар (EI) (энергия ионизации: 70эВ). Для идентификации метиловых эфиров жирных кислот липидов использовали значения индекса удерживания Rf (для стандартных насыщенных и ненасыщенных ЖК) и индекса ECL (эквивалентной длины цепи), а также библиотеку масс-спектров NIST05.
Относительное содержание ЖК определяли в весовых процентах от общего их содержания в исследуемом образце. Для оценки ненасыщенности ЖК в тканях листьев использовали индекс двойных связей (ИДС): ИДС=∑ P j n /100, где P j - содержание ЖК (вес. %) и n - количество двойных связей в каждой кислоте. Так же использовали коэффициент ненасыщенности жирных кислот (К) как отношение суммы ненасыщенных ЖК к сумме насыщенных.
Активность ацил-липидных ω9, ω6 и ω3 мембранных десатураз, участвующих в биосинтезе олеиновой, линолевой и α-линоленовой кислот, определялась соответственно по уравнениям (1, 2 и 3):
-
(1) SDR = (%С18:1)/(%С18:0+%С18:1)
-
(2) ODR = (%C18:2 + %C18:3)/(% C18:1+% C18:2 + %C18:3)
-
(3) LDR = (%C18:3)/( % C18:2 + %C18:3)
В таблицах представлены средние данные из трех биологических повторностей и их стандартные отклонения. Достоверность различий сравниваемых средних значений оценивали с помощью t-критерия (Р<0.05).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Впервые исследован жирнокислотный состав пяти видов лекарственных растений, произрастающих на юго-восточном побережье Байкала. Из представленных данных (табл. 1) видно, что высокая степень ненасыщенности ЖК в тканях листьев всех трех видов обусловлена, в основном, двумя кислотами – линолевой (ω6) и ά-линоленовой (ω3). Причем линоленовой ЖК (С18:3ω3) статистически достоверно больше наблюдалось у тысячелистника и одуванчика (58,51 и 59,06 % соответственно), чем у подорожника (52,1%).
Содержание линоленовой кислоты (С18:2ω6) у всех видов статистически не отличалось. Известно, что именно биосинтез диеновых и триеновых кислот 18:2ω6 и 18:3ω3 осуществляет биохимическую адаптацию растений к низкой температуре. Судя по вычисленным коэффициентам ORD и LRD (табл. 2), активность соответствующих десатураз (ω6 и ω3) также имела высокий уровень у представленных видов, особенно активность ω6-десатуразы.
Из ω9-жирных кислот в тканях исследуемых растений обнаружены моноеновые кислоты – олеиновая (С18:1ω9) и пальмитолеиновая (С16:1ω9). В сумме содержания этих кислот составили для тысячелистника 1,59 ± 0,33, для подорожника 1,21 ± 0,25 и для одуванчика 0,95
± 0,16 % веса. Причем содержание олеиновой кислоты у всех пяти видов лекарственных растений было на один порядок выше, чем пальмитолеиновой ЖК (табл. 1).
Содержание ω5-жирной кислоты С16:1ω5 не уступало ω9-жирным кислотам и достигало у тысячелистника 2,12, у одуванчика 1,68 и у подорожника 1,38 %.
Так же в исследованных растениях обнаружены ω7-ЖК, но их содержания составляли доли процента, а у одуванчика – десятые доли процента, что может свидетельствовать о меньшей активности ω7-десатуразы у одуванчика по сравнению с остальными исследованными видами. Максимальное содержание ω7-ЖК отмечено у тысячелистника (табл. 1).
Таблица 1. Жирнокислотный состав листьев пяти видов лекарственных растений.
|
Тысячелистник |
Подорожник |
Одуванчик |
|
|
Кислота |
сод. (в % вес.) |
сод. (в % вес.) |
Сод. (в % вес.) |
|
С14:0 |
0,58 ± 0,16 |
0,75 ± 0,09 |
0,42 ± 0,33 |
|
С15:0 |
0,06 ± 0,01 |
0,26 ± 0,04 |
0,2 ± 0,11 |
|
С16:0 |
15,02 ± 0,29 |
16,94 ± 0,94 |
16,74 ± 1,49 |
|
C16:1(n-9) |
0,05 ± 0,03 |
0,2 ± 0,05 |
0,03 ± 0,01 |
|
C16:1(n-7) |
0,18 ± 0,09 |
0,08 ± 0,03 |
0,03 ± 0,01 |
|
C16:1(n-5) |
2,12 ± 0,52 |
1,38 ± 0,16 |
1,68 ± 0,42 |
|
C17:0 |
0,15 ± 0,02 |
0,26 ± 0,01 |
0,15 ± 0,01 |
|
C18:0 |
0,92 ± 0,10 |
2,17 ± 0,12 |
0,92 ± 0,07 |
|
C18:1(n-9) |
1,54 ± 0,30 |
1,01 ± 0,20 |
0,92 ± 0,15 |
|
C18:1(n-7) |
0,13 ± 0,05 |
0,12 ± 0,04 |
0,04 ± 0,01 |
|
C18:2(n-6) |
19,92 ± 0,98 |
21,04 ± 1,27 |
18,82 ±3,01 |
|
C18:3(n-3) |
58,51 ± 0,81 |
52,1 ± 2,59 |
59,06 ± 3,55 |
|
C18:3(n-6) |
0,5 ± 0,21 |
||
|
C18:4(n-3) |
1,25 ± 0,43 |
||
|
C20:0 |
0,18 ± 0,09 |
0,68 ± 0,12 |
0,2 ± 0,06 |
|
C20:3(n-6) |
0,09 ± 0,01 |
||
|
C22:0 |
0,4 ± 0,02 |
0,83 ± 0,07 |
0,46 ± 0,07 |
|
C23:0 |
0,23 ± 0,06 |
0,3 ± 0,02 |
0,32 ± 0,04 |
Таблица 2 Суммы кислот по наличию и числу двойных связей и по длине углеродной цепи, индексы насыщенности и десатуразные соотношения у пяти видов лекарственных растений Прибайкалья.
|
Тысячелистник |
Подорожник |
Одуванчик |
|
|
Σsfa |
17,54 ± 0,75 |
22,19 ± 1,41 |
19,41 ± 2,18 |
|
Σufa |
82,45 ± 2,78 |
77,77 ± 4,99 |
80,59 ± 7,16 |
|
ИДС |
2,19 ± 0,05 |
2,08 ± 0,13 |
2,18 ± 0,17 |
|
К |
4,70 ± 0,04 |
3,50 ± 0,06 |
4,15 ± 0,10 |
|
SDR |
0,64 ± 0,12 |
0,34 ± 0,07 |
0,5 ± 0,08 |
|
ORD |
0,98 ± 0,03 |
0,99 ± 0,05 |
0,99 ± 0,11 |
|
LRD |
0,66 ± 0,01 |
0,71 ± 0,03 |
0,76 ± 0,04 |
|
∑ С14-15 |
0,64 ± 0,17 |
1,01 ± 0,13 |
0,62 ± 0,44 |
|
∑ С16-17 |
17,52 ± 0,95 |
18,86 ± 1,19 |
18,63± 1,94 |
|
∑ С18-19 |
81,02 ± 2,24 |
78,19 ± 4,86 |
79,76 ± 6,79 |
|
∑ С20-23 |
0,81 ± 0,17 |
1,9 ± 0,22 |
0,98 ± 0,17 |
|
∑ С х:0 |
17,54 ± 0,75 |
22,19 ± 1,41 |
19,41 ± 2,18 |
|
∑ С х:1 |
4,02 ± 0,99 |
2,79 ± 0,48 |
2,7 ± 0,60 |
|
∑ С х:2 |
19,92 ± 0,98 |
21,04 ± 1,27 |
18,82 ± 3,01 |
|
∑ С х:3 |
58,51 ± 0,81 |
52,69 ± 2,81 |
59,06 ± 3,55 |
|
∑ С х:4 |
0 |
1,25 ±0,43 |
0 |
Примечание: Σ SFA – сумма насыщенных ЖК, Σ UFA – сумма ненасыщенных ЖК, ИДС – инжекс двойных связей, К – коэффициент ненасыщенности, SDR – стеароил-десатуразное соотношение, ODR – олеил-десатуразное соотношение, LDR – линолеил-десатуразное соотношение, ∑ С14-15 – сумма короткоцепочечных ЖК, ∑ С16-17 – сумма ЖК с длиной цепи 16 и 17 атомов углерода, ∑ С16-17 – сумма ЖК с длиной цепи 18 и 19 атомов углерода, ∑ С20-23 – сумма длинноцепочечных ЖК, ∑ С х:0 – сумма насыщенных ЖК, ∑ Сх:1 – сумма моноеновых ЖК, ∑ Сх:2 – сумма диеновых ЖК, ∑ Сх:3 – сумма триеновых ЖК, ∑ Сх:4 – сумма тетраеновых ЖК.
Таблица 3 Температуры воздуха и почвы в день отбора проб растений (Выдринная)
|
Месяц/температура |
июль |
август |
сентябрь |
|
Минимальная температура воздуха, °С |
+8.1 |
+11.5 |
+2.7 |
|
Максимальная температура воздуха, °С |
+20.7 |
+18.9 |
+5.4 |
|
Средне-суточная температура воздуха,°С |
+15.1 |
+14.8 |
+4.1 |
|
Температура почвы (глубина 5 см) |
+17.8 |
+15.1 |
+5.2 |
Таблица 4. Жирнокислотный состав и показатели активности десатураз во второй половине лета и начале осени
А
Б
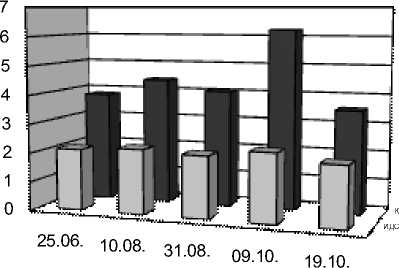
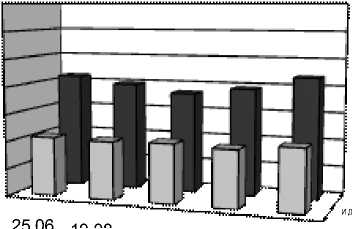
К
10.08. 31.08. 09.10. 19.10.
В
Г
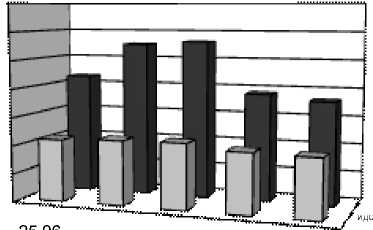
10.08. 31.08. 09.10. 19.10.
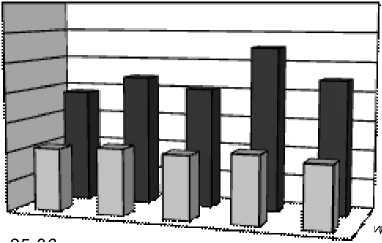
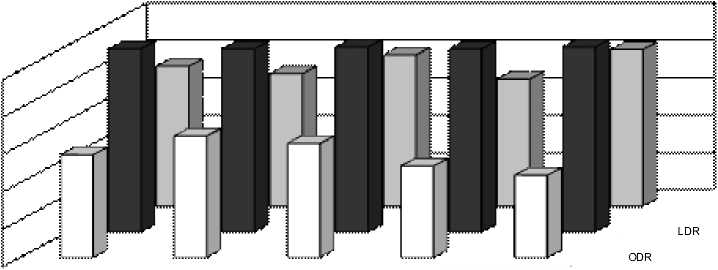
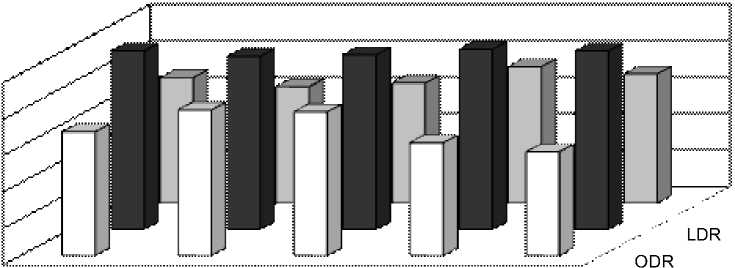
Индексы LDR, ODR и SDR (табл. 2), отражающие активность ω3-, ω6- и ω9-десатураз для ЖК с 18-ю атомами углерода, показали одинаково высокую активность ω6-десатураз у исследованных видов, соответствующую значениям ODR 0,98-0,99. Активность ω3-десатураз у изученных растений ниже и индекс LDR составляет для одуванчика 0,75, для подорожника 0,71, а у тысячелистника всего 0,66. Индекс SDR, напротив, у тысячелистника максимален (0,64), что свидетельствует об
относительно высокой активности ω9-десатураз. У остальных изученных растений этот индекс составил 0,50 у одуванчика, а у подорожника и он еще ниже (0,34). По сумме трех индексов (LDR+ODR+SDR) можно сделать вывод, что суммарная активность ω3-, ω6- и ω9-десатураз наибольшая у тысячелистника и относительно низкая у подорожника. В то же время, у подорожника обнаружены ω3-тетраеновые (1,25%) кислоты. Четыре ненасыщенных связи в этих ЖК могут существенно повышать
Д
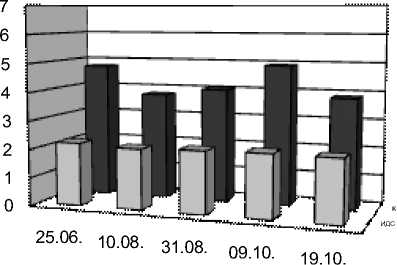
25.06. 10.08. 31.08. 09.10. 19.10.
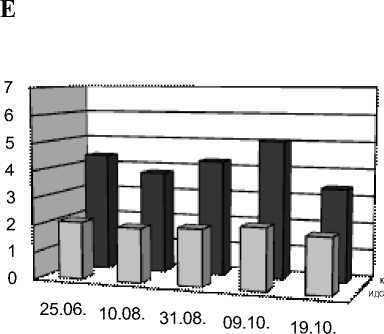
Рис. 1. Индексы двойных связей (ИДС) и коэффициенты ненасыщенности (К) у растений в г. Иркутске и Выдрино в течение периода вегетации
А – одуванчик г. Иркутск, Б – одуванчик Выдрино,
В – подорожник г. Иркутск, Г – подорожник Выдрино,
Д – тысячелистник Иркутск, Е – тысячелистник Выдрино
А
0,8
0,6
0,4
0,2
25.06.
10.08. 31.08. 09.10.
19.10. SDR
Б
0,8
0,6
0,4
0,2
25.06.
10.08.
31.08. 09.10.
19.10.
SDR
В
0,8
0,6
0,4
0,2

25.06. 1 0.08. 31.08. 09.1 0. 1 9.1 0. SDR
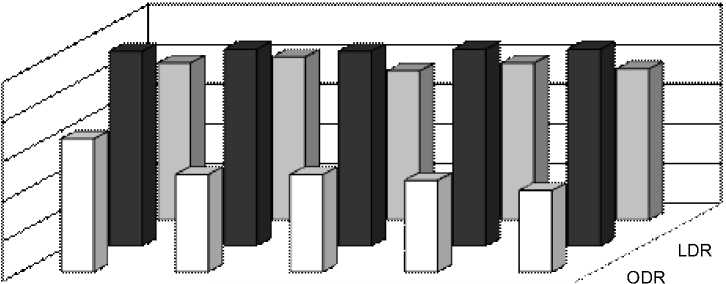
Рис. 2 Индексы SDR, ODR и LDR у растений в Иркутске в течение периода вегетации
А – одуванчик, Б – подорожник, В – тысячелистник
А
0,8
0,6
0,4
0,2
25.06.
10.08.
31.08.
09.10.
19.10.
SDR
Б
0,8
0,6
0,4
0,2

25.06.
10.08.
31.08.
09.10.
19.10.
SDR
В
0,8
0,6
0,4
0,2
25.06.
10.08. 31.08. 09.10.
19.10. SDR
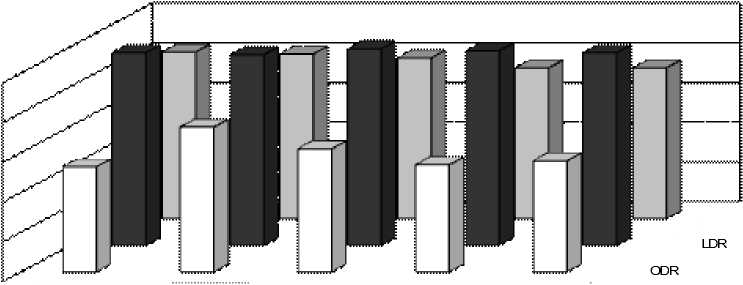
Рис. 3 Индексы SDR, ODR и LDR у растений в Выдрино в течение периода вегетации А – одуванчик, Б – подорожник, В – тысячелистник
А
Б
В
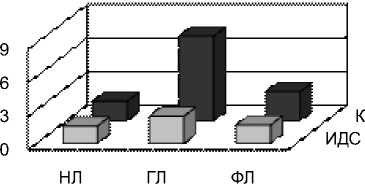
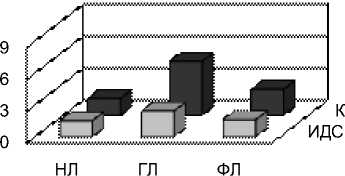
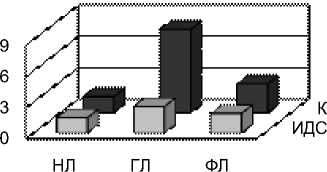
Рис. 4 Индексы ИДС и коэффициенты К одуванчика, тысячелистника и подорожника в сентябре у разных классов липидов: нейтральных липидов (НЛ), гликолипидов (ГЛ), фосфолипидов (ФЛ)
А – одуванчик, Б – подорожник, В – тысячелистник
А
Б

НЛ ГЛ ФЛ
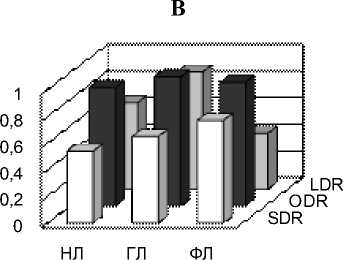
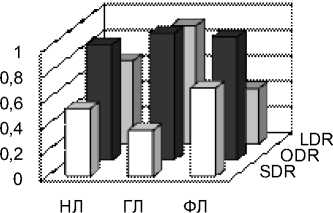
Рис. 5 SDR, ODR и LDR одуванчика, подорожника и тысячелистника в сентябре у разных классов липидов: нейтральных липидов (НЛ), гликолипидов (ГЛ), фосфолипидов (ФЛ) А – одуванчик, Б – подорожник, В – тысячелистник
Индекс ИДС, интегральная величина, характеризующая степень ненасыщенности жирных кислот, и, по-видимому, связанный напрямую с устойчивостью растений к холоду, был максимальным (2,19) у тысячелистника, как и коэффициент ненасыщенности (4,7 соответственно). Таким образом, у тысячелистника содержание ненасыщенных ЖК в 4,7 раз больше, чем насыщенных. Для сравнения, у одуванчика коэффициент ненасыщенности составил 4,2, а у подорожника – 3,5.
Длина углеродной цепи ЖК также может влиять на свойства клеточных мембран, в том числе и на их холодостойкость (Лось Д.А., 2005). В таблице 2 также представлены суммарные содержания короткоцепочечных ЖК (С14-15), ЖК с длинами цепей в 16-17 и 18-19 атомов углерода, а также длинноцепочечных ЖК с углеродными цепочками более 20 атомов (С2023). Обращает на себя внимание высокое содержание длинноцепочечных ЖК у подорожника, различие с двумя другими видами статистически достоверно. У всех исследованных видов преобладали16- и 18-углеродные ЖК, что в целом характерно для зеленых листьев растений, причем сумма 16С- и 18С-ЖК составила 97–98,5 % веса всех ЖК.
Также в таблице 2 представлены суммы ЖК не по длине цепи, а по числу двойных связей в молекуле. У всех растений преобладали триеновые ЖК. Их содержание было достоверно выше у тысячелистника и одуванчика, чем у подорожника. Содержание диеновых кислот у исследованных растений статистически не отличалось. Моноеновых кислот у всех трех видов растений мало и их содержания достоверно не отличаются. Тетраеновые кислоты встречены в небольшом количестве у подорожника, и у остальных двух видов пока не обнаружены.
Далее было изучено изменение жирнокислотного состава лекарственных растений в связи с наступлением осенних холодов (табл 3).
В таблице 4 представлены результаты анализа сезонной динамики содержания насыщенных и ненасыщенных жирных кислот в тканях листьев и активности десатураз для тысячелистника, одуванчика и подорожника.
Как видно из представленных данных (табл. 4), степень ненасыщенности жирных кислот в тканях листьев изучаемых растений меняется в процессе вегетации. Во всех случаях по мере снижения среднесуточной температуры (табл. 3) наблюдали снижение содержания насыщенных и увеличение содержания ненасыщенных ЖК, что было наиболее выражено у тысячелистника (табл. 4). Индексы содержания двойных связей К и ИДС постепенно увеличивались по мере снижения среднесуточной температуры, причем у тысячелистника отмечается наиболее сильная вариабельность этих показателей. Возможно, это связано с более широкой нормой реакции этого растения и свидетельствует о высокой способности тысячелистника к низкотемпературной адаптации.
Установлены также изменения в активности десатураз, в ответ на снижение среднесуточной температуры (табл. 4). Адаптация мембранных липидов к понижению среднесуточной температуры воздуха у исследованных растений носила видоспецифичный характер и была связана с активностью разных десатураз. Так, у одуванчика по мере снижения температуры возрастала активность ω3-десатуразы (LDR менялся с 0,69 до 0,80) при понижении активности ω9-десатуразы, а высокий уровень ненасыщенности в тканях листьев тысячелистника поддерживается, по всей видимости, активностью и ω9- (SDR меняется от 0,46 до 0,63) и ω3-десатуразы. У подорожника наблюдается снижение активности и ω9- и ω3-десатуразы, но активируется ω6-десатураза. В целом, активность ω6-десатуразы оставалась неизменно высокой у всех изучаемых видов.
На третьем этапе исследований кроме растений, произрастающих на юго-восточном побережье Байкала, анализировался состав растений, собранных в Иркутске. Это позволяет сравнить адаптационные особенности изучаемых растений в разных климатических условиях произрастания.
Среднемесячные температуры мест отбора проб приведены в таблице 5, а данные по ЖК и активности десатураз – на рисунках 1,2 и 3.
Полученные данные (рис. 1) показывают сильное увеличение индекса ненасыщенности ИДС у одуванчика в августе и особенно в начале октября в Иркутске, что скорее всего связано с адаптацией к понижению температур. Сильное снижение К во второй половине октября может быть связано с истощением защитных механизмов растения. У подорожника так же к концу августа увеличивается адаптация к пониженным температурам за счет НН ЖК, но уже даже в первых числах октября наблюдается сильное истощение защитных ресурсов растительного организма. В Выдрино, наоборот, в середине лета наблюдали относительно более высокие значения обоих индексов, которые к августу снижались, а осенний пик был смещен на конец октября у одуванчика и на начало октября у подорожника. У тысячелистника в Иркутске отмечалось высокое содержание НН ЖК и высокие значения ИДС и К, которое к концу месяца значительно снижалось. В Выдрино наблюдались высокие показатели ненасыщенности ЖК тысячелистника в июне, спад к началу августа и постепенный рост к началу октября с минимумом в конце месяца. Таким образом, у растений, произрастающих в непосредственной близости от Байкала, в июне наблюдалась более сильная жирнокислотная адаптация к низким температурам по сравнению с иркутскими растениями, в августе, наоборот, растения из Выдрино были приспособлены к более теплому климату, а в октябре дольше сохраняли устойчивость к морозам.
Коэффициенты, отражающие активность десатураз в изучаемых растениях (рис. 2, 3), показали для одуванчика пик активности ω3-десатуразы в июле для Иркутска и в августе для Выдрино, а пики активности ω9-десатуразы в Иркутске шли в противофазе с Иркутскими. Для подорожника в Иркутске отмечалось закономерное уменьшение активности ω6-десатуразы с приходом осенних морозов, в то время как для Выдрино было характерно, чередование пиков активности этого фермента, причем в конце октября также наблюдалось увеличение активности этой десатуразы. В свою очередь, активность ω3-десатуразы подорожника в Иркутске была максимальной в начале августа, после чего ее значения постепенно снижались, в то время как в Выдрино наблюдалось два максимума – в июле и начале октября. У тысячелистника в Выдрино наблюдали постепенное увеличение LDR с июня по октябрь, а в Иркутске, наоборот, его уменьшение по мере уменьшения среднесуточных температур.
Также в сентябрьских пробах были исследованы отдельные классы липидов в изучаемых растениях: нейтральные липиды (НЛ)
и глико- (ГЛ) и фосфолипиды (ФЛ) мембран. Для каждой фракции липидов были определены коэффициенты ИДС, К, SDR, ODR и LDR (рис. 3, 4). Видно, что степень ненасыщенности ЖК распределена неравномерно как по классам липидов, так и по видам растений. В целом, ИДС и К наиболее высоки у гликолипидов, что вполне закономерно, так как эти липиды являются основными состовляющими мембран хлоропластов, которым присуще высокое содержание ά-линоленовой кислоты. При этом у подорожника наибольший ИДС, составивший 2,56 (у одуванчика и тысячелистника 2,41 и 2,43 соответственно). Менее показательный, но более вариабельный коэффициент К наибольший также у подорожника (8,04), чуть меньше он у тысячелистника (7,59) и наименьший у одуванчика (5,07).
Из нейтральных липидов и фосфолипидов преобладает ненасыщенность ФЛ. Ее значение также максимально у подорожника, а минимально у одуванчика.
В то же время, индексы SDR были максимальными для всех трех классов у тысячелистника. Из данных (рис. 4) видно, что из трех индексов, отражающих активность десатураз, SDR является самым вариабельным и видоспецифичным, в то время как остальные два почти одинаковы у всех трех видов и максимальны для ГЛ.
В контексте современных представлений о вкладе липидов биологических мембран клетки в холодо- и морозоустойчивость (Макаренко С.П. и др., 2003) полученные данные дают основание полагать, что изученные виды обладают высоким потенциалом холодоустойчивости, связанным с особенностями жирнокислотного состава мембранных липидов.
ВЫВОДЫ
Впервые определен жирнокислотный состав тысячелистника, одуванчика и подорожника. В связи со снижением среднесуточной температуры, выявлено снижение содержания насыщенных и увеличение содержания ненасыщенных жирных кислот. Низкотемпературная адаптация мембранных липидов изучаемых растений оказалась видоспецифичной и была связана с увеличением активности разных десатураз. На побережье Байкала отмечалась более высокая насыщенность мембранных липидов растений по сравнению с Иркутском, в то время как поздним летом и осенью в Иркутске жирнокислотная термоадаптация развивалась сильнее и раньше, чем в Выдрино. Показано, что из трех классов липидов, максимальный вклад в холодовую адаптацию вносят гликолипиды и заметно меньшую – фосфолипиды.
Работа выполнена при финансовой поддержке гранта РФФИ 07-04-01055-а, РФФИ р Сибирь а 08-04-98040.
Список литературы Изменение жирнокислотного состава в растениях при гипотермической адаптации
- Ильинская Л.И., Озерецковская О.Л. (1998) Продукты липоксигеназного окисления жирных кислот как сигнальные молекулы в индуцированной устойчивости растений (обзор) Прикл. биохимия и микробиология. 34(5) 467-479.
- Колесниченко А.В., Войников В.К. (2003) Белки низкотемпературного стресса растений. Иркутск: Арт-Пресс,. 196.
- Лось Д.А. (2001) Структура, регуляция экспрессии и функционирование десатураз жирных кислот Успехи биологической химии. 41. 163-198.
- Лось Д.А. (2005) Молекулярные механизмы холодоустойчивости растений Вестник РАН. 75(4) 338-345.
- Макаренко С.П., Константинов Ю.М., Хотимченко С.В., Коненкина Т.А., Арзиев А.Ш. (2003) Жирнокислотный состав липидов митохондриальных мембран у представителей культурных (Zea mays) и дикорастущих (Elymus sibiricus) злаков Физиология растений. 50(4) 548-553.
