Изменения базовых онтогенетических спектров популяций некоторых редких видов растений Самарской области при антропогенной нагрузке на местообитания

Автор: Ильина В.Н.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 3 т.24, 2015 года.
Бесплатный доступ
В статье приведены многолетние данные по онтогенетической структуре природных популяций редких в самарской флоре видов растений. Указаны основные тенденции динамики структуры 33 представителей ( Ajuga chia, Anthemis trotzkiana, Artemisia salsoloides, Astragalus cornutus, A. helmii, A. henningii, A. macropus, A. physocarpus, A. sulcatus, A. temirеnsis, A. ucrainicus, A. wolgensis, A. zingeri, Atraphaxis frutescens, Chrysocyathus volgensis, Clausia aprica, Ferula caspica, F. tatarica, Galatella angustissima, Gentiana pneumonanthe, Hedysarum gmelinii, H. grandiflorum, H. razoumovianum, Iris pumila, Jurinea ewersmannii, J. ledebourii, J. multiflora, Laser trilobum, Oxytropis floribunda, O. hippolyti, O. spicata, Polygala sibirica, Tanacetum achilleifolium) при возрастании антропогенного пресса на их местообитания.
Популяция, онтогенетический спектр, оптимум, стресс, антропогенные факторы, самарская область
Короткий адрес: https://sciup.org/148314964
IDR: 148314964 | УДК: 581.
Текст краткого сообщения Изменения базовых онтогенетических спектров популяций некоторых редких видов растений Самарской области при антропогенной нагрузке на местообитания
важно в условиях растущего антропогенного пресса и низкой степени соблюдения природоохранного законодательства в Российской Федерации и в Самарской области в частности. Как неоднократно указывается многими авторами, популяционно-онтогенетическое направление исследований играет значимую роль среди методов биологической индикации природных комплексов, выявлении и мониторинге охраняемых природных территорий различного ранга, создании и ведении региональных и федеральной Красных книг, выявлении биологических и экологических характеристик видов растений и т.д. (Уранов, 1975; Полуянова, 1985, 2007; Жукова, 1995, 2001; Хмелев, Скользнева, 1997; Тетерюк, 2000; Абрамова и др., 2001, 2010; Гуреева, Бытотова, 2001; Полуянова, Федорова, 2002, 2004; Ведерникова, Козырева, 2005; Маслова и др., 2005, 2012; Ильина В.Н., 2006, 2010, 2014; Пичугина, Савиных, 2006; Гаврилова, 2007; Жукова и др., 2007; Карнаухова и др., 2008; Полуянова, Любарский, 2008; Дорогова, Жукова, 2009; Ильина И.В., 2009; Мулдашев и др., 2009, 2010, 2011; Татарников, Корчиков, 2009, 2010, 2011; Шамигулова, Суюн-дуков, 2009; Жмудь и др., 2011, 2012; Карнаухова, 2011; Османова, 2011; Федорова, 2011; Атласова, 2012; Глазырина и др., 2012; Мустафина, Абрамова, 2012; Попова и др., 2012; Пузырькина и др., 2012; Савиных, Пересторонина, 2012; Сафонова, 2012; Зубаирова, 2013; Каримова и др., 2013; Карнаухова, Селютина, 2013; Османова, Ведерникова, 2013; Пузырькина, Силаева, 2013; Супрун, 2013 а, б; Родионова, Ильина, 2013; Чукина и др., 2013; Мустафина и др., 2014; Селютина и др., 2006 а, б, 2014; Abramova and other, 2014 и др.). При использовании данных методов становится возможным определение статуса редкости раритетных видов флоры, основных лимитирующих развитие особей и их популяций факторов, вероятных тенденций в развитии и современного состояния ценотических популяций, а также выявление общего состояния природных комплексов.
Одной из задач наших исследований является выявление особенностей онтогенетических спектров природных популяций растений-индикаторов для последующей экспресс оценки состояния степных сообществ и редкости видов в регионе (Ильина, 2006, 2010, 2014).
Онтогенетическая структура популяций растений является высоко информационным показателем состояния вида в сообществах на конкретной территории. Специальные индексы (восстановления, замещения особей и др.) указывают на тип популяции и определяют тенденции ее дальнейшего развития.
Рассматривая совокупность всех особей популяции, можно построить ее онтогенетический спектр. Поскольку последовательные онтогенетические состояния упорядочены во времени, календарный и биологический возраст особей неизбежно сильно скоррелированы, при этом особи одного онтогенетического состояния могут иметь разный календарный возраст (Уранов, 1975; Глотов и др., 2014).
Работы по экспресс-оценке сукцессионного состояния лесных сообществ осуществляются путем сравнения онтогенетических спектров и численности популяций видов древесных растений, входящих в его состав (Восточноевропейские леса, 2004). Исследования М.Н. Гавриловой (2009) ценопопуляций ракитника русского показывают, что в близких к оптимальным условиям местообитаниях онтогенетическая структура популяций имеет сходные показатели. Другими авторами (Щеглова, 2009; Валуйских, Тетерюк, 2013; Бийболатова, Аджиева, 2014) также указывается, что онтогенетическая структура популяций зависит от эколого-фитоценотических условий среды. Л.А. Сергиенко и А.В. Фокусов (2015) отмечают, что в быстро изменяющихся условиях среды значительные изменения претер- певают и онтогенетические спектры (на примере подорожника морского в приморских экосистемах). Относительно некоторых видов исследователи придерживаются мнения, что особенности онтогенетической структуры популяций зависят скорее не от экологических факторов, а обусловлены историей становления и развития конкретных популяций (Жукова и др., 2014).
Разработаны методы статистического анализа онтогенетической структуры ценопопуляций растений и эпифитных лишайников (Иванов, 2014).
Несомненно, у большинства редких представителей флоры онтогенетические спектры популяций в изменяющихся условиях среды будут отличаться от таковых в климаксовых ценозах. Различные изменения в основном связаны с воздействием на растительный покров антропогенных факторов. Работы различных авторов свидетельствуют о формировании «характерных» онтогенетических спектров популяций (Заугольнова, 1976; Валуйских, Тетерюк, 2013), что обусловлено биологией конкретного вида.
Нами с 1998 года осуществляется изучение онтогенетической структуры редких и уязвимых представителей флоры Самарской и сопредельных областей.
В данной статье приводятся базовые онтогенетические спектры популяций 33 видов редких видов растений, включенных в Красную книгу Самарской области (2007) или рекомендованных к внесению в списки охраняемых в регионе, в условиях антропогенного пресса на их местообитания. В скобках указано число учтенных ценопопуляций для определения базового онтогенетического спектра (опти-мум/стресс). Данные собраны в 1998-2014 гг. в Самарской, Ульяновской и Оренбургской областях. Модельными видами явились Ajuga chia Schreb. (6/8), Anthemis trotzkiana Claus (2/4), Artemisia salsoloides Willd. (23/26) , Astragalus cornutus Pall. (8/12), A. helmii Fisch. (14/20), A. henningii ( Stev.) Klok. (12/26), A. macropus Bunge (14/25), A. physocarpus Ledeb . (3/6), A. sulcatus L. (4/9), A. temirеnsis M. Pop. (2/4), A. ucrainicus Klok. et M. Pop. (2/2), A. wolgensis Bunge (13/27), A. zingeri Korsh. (12/26), Atraphaxis frutescens ( L.) C. Koch. (14/45), Chrysocyathus volgensis (DC.) Holub (24/67), Clausia aprica ( Steph.) Korn.-Tr. (6/16), Ferula caspica Bieb. (12/15), F. tata-rica Fisch. ex Spreng. (11/9) , Galatella angustissima ( Tausch) Novopokr. (6/15), Gen-tiana pneumonanthe L. (7/11), Hedysarum gmelinii Ledeb. (56/137), H. grandiflorum Pall. (78/214), H. razoumovianum Fisch. et Helm (66/179), Iris pumila L. (32/68), Jur-inea ewersmannii Bunge (4/12), J. ledebourii Bunge (23/45), J. multiflora (L.) B. Fedtsch. (12/44), Laser trilobum (L.) Borkh. (12/36) , Oxytropis floribunda ( Pall.) DC. (23/57), O. hippolyti Boriss. (12/6), O. spicata ( Pall.) O. et B. Fedtsch. (12/35), Polygala sibirica L. (9/14), Tanacetum achilleifolium (Bieb.) Sch. Bip. (5/11).
Хотя первоначальное деление местообитаний с оптимальными и стрессовыми условиями проводилось нами по флористическим параметрам и визуально, то последующий анализ показывает на существенные различия в онтогенетических спектрах популяций. При проведении биоиндикации экспресс-методами это позволит определить степень воздействия антропогенных факторов (низкая или высокая) на фитоценозы по нескольким индикаторным видам.
Среди основных антропогенных факторов, действующих на фитоценозы с участием модельных представителей, необходимо назвать перевыпас крупного рогатого скота, степные пожоги, высокую рекреацию, сенокошение, отчуждение территории. В некоторых местообитаниях наблюдается комплекс факторов, воздействующих на растительный покров. Однако чаще всего степень воздействия оценивается числом редких видов, либо изменением таксономического состава за некото- рый период времени. Эти показатели не всегда можно считать стопроцентно достоверны в связи с отсутствием мониторинговых исследований и невозможностью сравнения результатов, свойствами флоры как динамической структуры и другими различными причинами.
Базовые онтогенетические спектры популяций растений в составе растительных сообществ имеют свои закономерности в зависимости от жизненной формы представителя, длительности онтогенеза и отдельных его стадий, типа жизненной стратегии.
Типы базовых спектров обычно выделяют по положению абсолютного максимума особей того или иного онтогенетического состояния в этом спектре. В зависимости от способа самоподдержания ценотической популяции в каждом из типов выделяют варианты спектров. Инвазионные спектры (или популяции) - в них представлены прегенеративные и иногда молодые генеративные особи. В нормальном типе спектра выделяют полночленные, вегетативно-полночленные (для популяций, состоящих из особей, образовавшихся вегетативным путем) и прерывистые варианты (присутствуют не все онтогенетические группы особей). Регрессивная популяция состоит только из постгенеративных особей. Фрагментарный спектр характеризуется наличием одной или нескольких онтогенетических групп особей. Основополагающими работами в данном направлении являются труды Т.А. Работного (1950), А.А. Уранова (1975), О.В. Смирновой (2004). Классификация нормальных популяций предложена Л.А. Жуковой (1967), А.А. Урановым и О.В. Смирновой (1969): в молодой нормальной популяции преобладают виргинильные и молодые генеративные растения, в зрелой - средневозрастные генеративные, в стареющей - старая генеративная фракция, в старой - субсенильные особи.
На наш взгляд, для характеристики популяций необходимо учитывать все классификации. Особенности популяций (онтогенетических спектров) приведены в таблице.
Популяции Ajuga chia в близких к оптимальным условиям местообитаниях чаще всего зрелые нормальные неполночленные с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных особях (46%). Доля генеративных особей около 60%. Отсутствуют две группы особей - проростки и сенильные растения. В стрессовых условиях популяции живучки стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (71%). Небольшой пик в спектре приходится на молодые генеративные растения (18%). Доля генеративных особей - 98%. Не зафиксированы проростки, ювенильные, имматур-ные, виргинильные и сенильные растения. Основными видами воздействия на сообщества с участием модельного вода - нерегламентированный выпас скота и степные пожоги.
У Anthemis trotzkiana в оптимуме популяции зрелые нормальные неполночленные с прерывистым одновершинным центрированным спектром с максимумом на зрелой генеративной фракции (46%). Не зафиксированы сенильные растения. Доля генеративных растений составляет 68%. При значительной антропогенной нагрузке они стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (42%). Отсутствуют проростки и сенильные группы особей. Вклад генеративных экземпляров в популяцию - 70%. Значительно увеличивается доля сенильных растений - с 10 до 25%. Снижение численности особей в популяции и из- менение структуры популяций вызывают выпас, степные палы или при отсутствии воздействия, напротив, развитие дернины злаков.
Для Artemisia salsoloides типичным является случай, когда природные популяции характеризуются как зрелые нормальные полночленные с одновершинным центрированным спектром с максимумом на зрелых генеративных растениях (45%). Примерно одинаковые позиции занимают виргинильные, молодые генеративные и старовозрастные генеративные особи (15-17%). Генеративная фракция составляет около 76%. В стрессовых условиях популяции полыни стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных экземплярах (31%). Отсутствуют проростки, ювенильные и имматурные растения. Генеративное ядро популяции представлено 61% зарегистрированных особей. Велика доля растений сенильного периода онтогенеза - 36%. Основными видами воздействия на сообщества с участием модельного вода - нерегламентированный выпас скота, степные пожоги, избыточная рекреация.
Популяции Astragalus cornutus в условиях, близких к оптимальным, молодые нормальные полночленные с одновершинным центрированным спектром с максимумом на молодых генеративных особях (42%). Генеративные особи в составе популяций занимают ведущее положение (75%). При возрастании нагрузки на местообитания популяции переходят в зрелые нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на зрелых генеративных растениях (45%). В спектре наблюдается небольшой пик на виргинильных особях (14,5%). Генеративное ядро популяций - более 80%. Отсутствуют проростки, ювенильные и имматурные растения. Факторами, вызывающими изменения структуры популяций, являются сенокошение, пожоги, перевыпас и рекреация.
У Astragalus helmii в оптимуме популяции зрелые нормальные полночленные с одновершинным центрированным спектром с максимумом на зрелых генеративных растениях (23%). Однако высоко значение в популяциях и других групп генеративных особей - по 20-21%. Число генеративных особей достигает 75%. Стареющие нормальные неполночленные популяции с прерывистым одновершинным правосторонним онтогенетическим спектром с максимумом на старовозрастных генеративных экземплярах (37%) отмечаются при нерегулируемой пастьбе скота и проводимых палах. В популяциях не зафиксированы проростки, ювенильные и сенильные группы особей. Генеративные растения в сумме составляют 83%. Сенильные особи составляют примерно 6-10% во всех популяциях.
Astragalus henningii в оптимальных условиях существования отмечается в составе зрелых нормальных полночленных популяций с одновершинным центрированным спектром с примерно равным распределением между молодыми, зрелыми и старовозрастными генеративными растениями (по 20-23%). В этих популяциях астрагала генеративные растения составляют 64% от общего числа особей. Возрастающий антропогенный пресс приводит к изменениям, и популяции являются стареющими нормальными неполночленными с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (37%). Отсутствуют проростки, ювенильные, имматурные и сенильные экземпляры. Генеративное ядро насчитывает 77% особей. К этому так же приводят перевы-пас и степные пожары. Однако высока доля виргинильных и молодых генеративных особей (15-16%), что свидетельствует о возможности восстановления популяций при снижении нагрузки на местообитания.
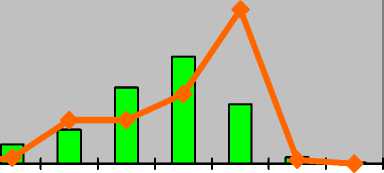
Для Astragalus macropus характерны молодые нормальные полночленные популяции с двувершинным левосторонним спектром с максимумом на виргиниль-ных особях (32%). Второй пик образуют зрелые генеративные растения (26%). В некоторых случаях популяции следует считать зрелыми нормальными. Генеративные особи составляют около 41% зарегистрированных. Стрессовые факторы (пере-выпас, пожоги) обусловливают переход популяций астрагала в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных экземплярах (около 36%). В спектрах отсутствуют проростки, ювенильные и имматурные растения. Возрастает генеративная часть популяций – 73%. В половину уменьшается число виргиниль-ных растений, однако в среднем их насчитывается около 16%.
Другой представитель этого рода, Astragalus physocarpus, характеризуется зрелыми нормальными полночленными популяциями с одновершинным центрированным спектром с максимумом на зрелых генеративных растениях (32%). Субдо-минируют старовозрастные генеративные растения – 30%. Популяции отличаются высоким содержанием генеративных особей – 82%. Однако даже в оптимальных условиях популяции не способны к эффективному самовосстановлению. Возросшая нагрузка на местообитания приводит к формированию стареющих нормальных неполночленных популяций с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных экземплярах (41%). Этому также способствуют перевыпас и проводимые нерегулируемые палы. При этом отсутствуют все особи предгенеративного периода (проростки, ювенильные, имматурные, виргинильные) – наблюдается их катастрофическая гибель. В малочисленных популяциях фиксируются только единичные генеративные (86%) и сенильные (14%) растения.
Astragalus sulcatus в условиях низкой нагрузки или ее отсутствии отмечается в зрелых нормальных неполночленных популяциях с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных растениях (31%). В популяциях не зафиксированы сенильные особи. Генеративное ядро представлено 76% особей. Антропогенная трансформация почвенно-растительного покрова способствует переходу популяций в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (35%). Также, как и в предыдущем случае, полностью выпадает левая (от проростков до виргинильного состояния) часть онтогенетического спектра. Генеративные особи представлены 86% экземпляров.
Очень редкий в регионе Astragalus temirеnsis отмечен в составе стареющих нормальных неполночленных популяций с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных экземплярах (40%). В связи с малой выборкой тип популяций в оптимуме и стрессовых условиях мало отличается. Популяции изучались в Сыртовом Заволжье. В обоих случаях отсутствуют проростки и ювенильные растения, при возрастании нагрузки также не фиксируются имматурные особи. Генеративное ядро в целом представлено 75% особей. Их соотношение изменяется по сезонам, но всегда преобладают старовозрастные генеративные растения (до 28%).
Таблица
Особенности базовых спектров популяций редких растений
|
Таксон |
Базовый онтогенетический спектр популяций |
Тип популяций |
|||||||||||||
|
Классификация ЦП А.А. Уранова (1975) |
Классификация ЦП О.В. Смирновой (2004) |
Классификация ЦП Л.А. Жуковой (1967) и А.А. Уранова и О.В. Смирновой (1969) |
|||||||||||||
|
Оптимум |
Стресс |
Оптимум |
Стресс |
Оптимум |
Стресс |
||||||||||
|
Ajuga chia Schreb. Живучка хиосская |
80 60 40 20 0 |
Неполночленные (отсутствуют p, s) |
Непол-ночлен-ные (от-сутст-вуют p, j, im, v, s) |
нормальные с прерывистым одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
||||||||
|
1_____1 оптимум ^^стресс |
|||||||||||||||
|
г |
|||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||
|
Anthemis trotzkiana Claus Пупавка Корнух-Троцкого |
50 40 30 20 10 0 |
Неполночленные (отсутствуют s) |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с прерывистым одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
||||||||
|
m . П . П . |
|||||||||||||||
|
1_____1 оптимум ^^стресс |
|||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||
|
Artemisia sal-soloides Willd. Полынь солянковидна я |
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j, im) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
|||||||||
|
40 30 20 10 0 |
|||||||||||||||
|
оптимум стресс |
|||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||
Astragalus cornutus Pall.
Астрагал рого-плодный
Astragalus helmii Fisch.
Астрагал
Гельма
Astragalus henningii ( Stev.) Klok. Астрагал Геннинга
Полно-
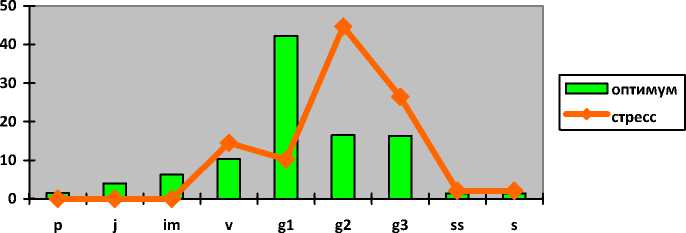
I-----1 оптимум ^^стресс p j im v g1 g2 g3 ss s
] оптимум стресс im
v
s g1 g2 g3 ss
p
членные
Полночленные
Полночленные
Непол-ночлен-ные (от-сутст-вуют p, j, im)
нормальные с полночленным одновершинным центрированным спектром
нормальные с прерывистым одновершинным правосторонним спектром
молодые нормальные с максимумом на g 1
зрелые нормальные с максимумом на g 2
|
Непол-ночлен-ные (от-сутст-вуют p, j, s) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
|
Непол- |
нормаль- |
нормаль- |
зрелые |
старею- |
|
ночлен- |
ные с |
ные с пре- |
нормаль- |
щие нор- |
|
ные (от- |
полно- |
рывистым |
ные с |
мальные |
|
сутст- |
членным |
одновер- |
максиму- |
с макси- |
|
вуют p, |
одновер- |
шинным |
мом на g 1, |
мумом |
|
j, im, s) |
шинным центрированным спектром |
правосторонним спектром |
g 2, g 3 |
на g 3 |
Astragalus macropus Bunge Астрагал длиннонож-ковый
Astragalus physocarpus Ledeb . [A. physodes auct .] Астрагал пузырчатоплодный
Astragalus sulcatus L.
Астрагал бороздчатый
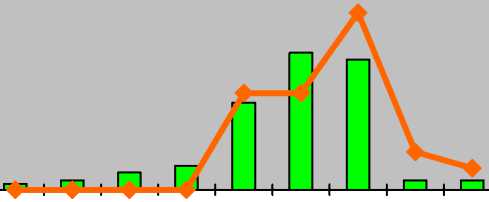
p j im v g1 g2 g3 ss s
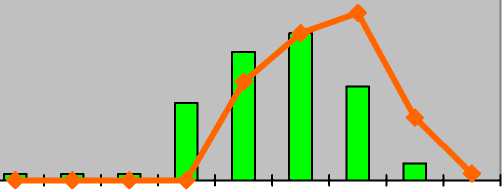
I-----1 оптимум ^^стресс p j im v g1 g2 g3 ss s
I-----1 оптимум ^^стресс
|
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j, im) |
нормальные с полночленным двувершинным левосторонним спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
молодые нормальные с максимумом на v |
стареющие нормальные с максимумом на g 3 |
|
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j, im, v) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
|
Неполночленные (отсутствуют s) |
Непол-ночлен-ные (от-сутст-вуют p, j, im, v) |
нормальные с прерывистым одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
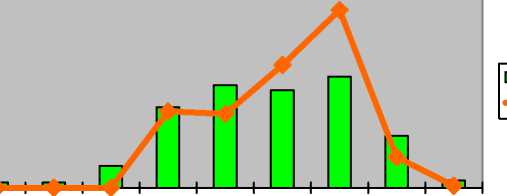
Astragalus temirеnsis M.
Pop. Астрагал темирский
Astragalus ucrainicus Klok. et M. Pop.
Астрагал украинский
Astragalus wolgensis Bunge Астрагал волжский
|
. П . _ |
|||||||||
I-----1 оптимум ^^стресс p j im v g1 g2 g3 ss s
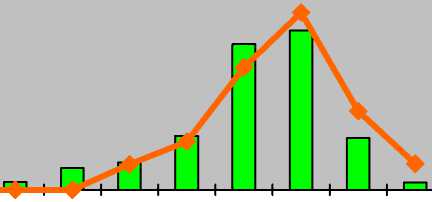
p j im v g1 g2 g3 ss s
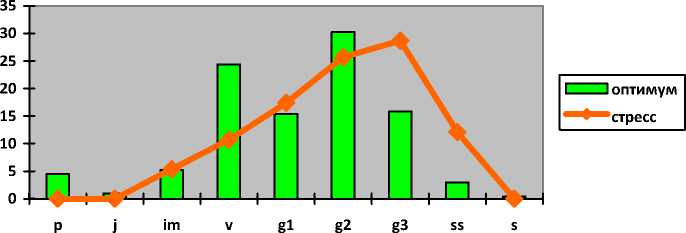
I ----- 1 оптимум ^^стресс
|
Неполночленные (отсутствуют p, j) |
Непол-ночлен-ные (от-сутст-вуют p, j, im) |
нормальные с прерывистым одновершинным правосторонним спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
стареющие нормальные с максимумом на g 3 |
стареющие нормальные с максимумом на g 3 |
|
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j, im) |
нормальные с полночленным одновершинным правосторонним спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
стареющие нормальные с максимумом на g 3 |
стареющие нормальные с максимумом на g 3 |
|
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j, s) |
нормальные с полночленным двувершинным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
|
Astragalus zingeri Korsh. Астрагал Цингера |
45 |
Неполночленные (отсутствуют s) |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
||||||||||||||
|
40 35 30 25 20 15 10 5 0 |
—1 |
||||||||||||||||||||
|
—1 |
1_____1 оптимум ^^стресс |
||||||||||||||||||||
|
' . гп . П . |
|||||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||||
|
Atraphaxis frutescens ( L.) C. Koch. Курчавка кустарникова я |
40 |
Неполночленные (отсутствуют s) |
Непол-ночлен-ные (от-сутст-вуют p, j, im) |
нормальные с прерывистым одновершинным правосторонним спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
стареющие нормальные с максимумом на g 3 |
зрелые нормальные с максимумом на g 2 |
||||||||||||||
|
35 30 25 20 15 10 5 0 |
|||||||||||||||||||||
|
1_____1 оптимум —•—стресс |
|||||||||||||||||||||
|
г-. . П . |
"1 |
||||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||||
|
Chrysocyathus volgensis (DC.) Holub (Adonis volgensis DC. ). Желтоцвет волжский |
Неполночленные (отсутствуют s) |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с прерывистым двувершинным левосторонним спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
молодые нормальные с максимумом на v |
стареющие нормальные с максимумом на g 3 |
|||||||||||||||
|
40 35 30 25 20 15 10 5 0 |
1 |
||||||||||||||||||||
|
— |
1_____1 оптимум ^^стресс |
||||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||||
|
Clausia aprica ( Steph.) Korn.-Tr. Клаусия солнцелюбив ая |
45 |
Неполночленные (отсутствуют j, s) |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с прерывистым одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
зрелые нормальные с максимумом на g 2 |
||||||||||||
|
40 35 30 25 20 15 10 5 0 |
|||||||||||||||||||
|
1_____1 оптимум —•—стресс |
|||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||
|
Ferula cas-pica Bieb. Смолоносица каспийская |
40 |
Неполночленные (отсутствуют j) |
Непол-ночлен-ные (от-сутст-вуют p, j, s) |
нормальные с прерывистым двувершинным левосторонним спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
молодые нормальные с максимумом на v |
стареющие нормальные с максимумом на g 3 |
||||||||||||
|
35 30 25 20 15 10 5 0 |
|||||||||||||||||||
|
1_____1 оптимум —•—стресс |
|||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||
|
Ferula tatari-ca Fisch. ex Spreng. Смолоносица татарская |
40 |
Неполночленные (отсутствуют p, s) |
Непол-ночлен-ные (от-сутст-вуют p, j, im) |
нормальные с прерывистым двувершинным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
молодые нормальные с максимумом на v |
стареющие нормальные с максимумом на g 3 |
||||||||||||
|
35 30 25 20 15 10 5 0 |
|||||||||||||||||||
|
1_____1 оптимум ^^стресс |
|||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||
|
Galatella an-gustissima ( Tausch) No-vopokr. Солонечник узколистный |
50 |
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
||||||||||||
|
40 30 20 10 0 |
1_____1 оптимум —•—стресс |
||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||
|
Gentiana pneumonanthe L. Горечавка легочная |
45 |
Неполночленные (отсутствуют p, j) |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с прерывистым одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
||||||||||||
|
40 35 30 25 20 15 10 5 0 |
|||||||||||||||||||
|
1_____1 оптимум ^^стресс |
|||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||
|
Hedysarum gmelinii Le-deb. Копеечник Гмелина |
50 |
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
||||||||||||
|
40 30 20 |
|||||||||||||||||||
|
1_____1 оптимум ^^стресс |
|||||||||||||||||||
|
10 0 |
_Г---1 ■ 1— ■ . Г 1 . |
. ГП . Г1 |
|||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||
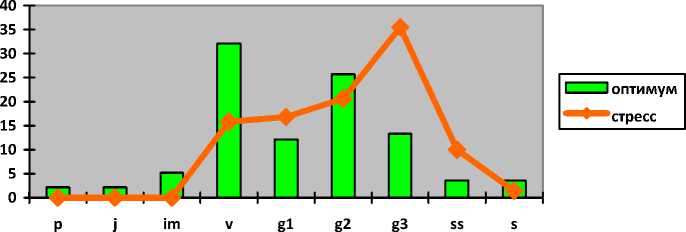
Hedysarum grandiflorum Pall.
Копеечник крупноцветковый
Hedysarum razoumovi-anum Fisch. et Helm Копеечник Разумовского
Iris pumila L. Касатик низкий
Полно-
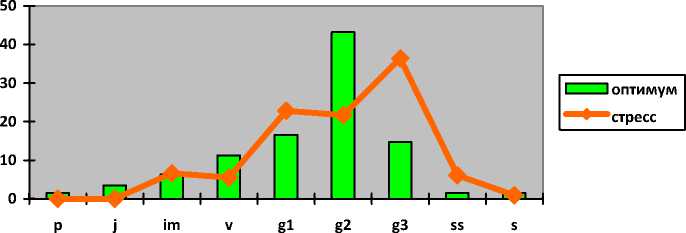
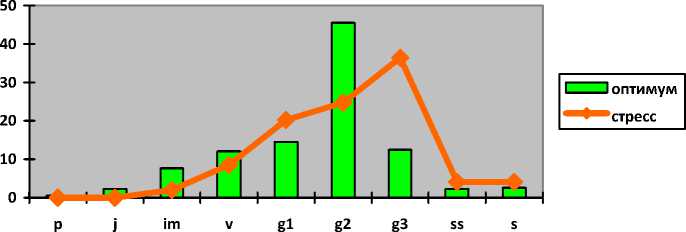
I-----1 оптимум ^^стресс
p j im v g1 g2 g3 ss s
членные
Полночленные
Неполночленные (отсутствуют p, j, im, s)
Непол-ночлен-ные (от-сутст-вуют p, j, s)
Непол-ночлен-ные (от-сутст-вуют p, j)
нормальные с полночленным одновершинным центрированным спектром
нормальные с полночленным одновершинным центрированным спектром
нормальные с прерывистым одновершинным правосторонним спектром
нормальные с прерывистым одновершинным правосторонним спектром
зрелые нормальные с максимумом на g 2
зрелые нормальные с максимумом на g 2
стареющие нормальные с максимумом на g3
стареющие нормальные с максимумом на g3
|
Непол- |
фрагмен- |
фрагмен- |
зрелые |
старею- |
|
ночлен- |
тарные с |
тарные с |
нормаль- |
щие нор- |
|
ные (от- |
одновер- |
одновер- |
ные с |
мальные |
|
сутст- |
шинным |
шинным |
максиму- |
с макси- |
|
вуют p, |
центриро- |
правосто- |
мом на g 2 |
мумом |
|
j, im, v) |
ванным |
ронним |
на g 3 |
|
|
спектром |
спектром |
Jurinea ew-ersmannii Bunge Наголоватка Эверсманна
Jurinea lede-bourii Bunge Наголоватка Ледебура
Jurinea multiflora ( L.) B. Fedtsch.
Наголоватка многоцветковая
I-----1 оптимум ^^стресс p j im v g1 g2 g3 ss s
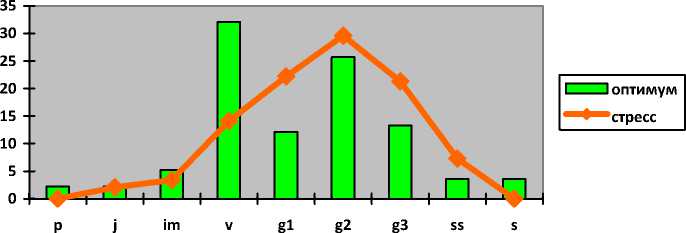

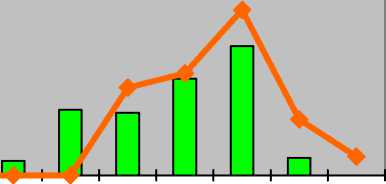
I-----1 оптимум ^^стресс p j im v g1 g2 g3 ss s
Неполночленные (отсутствуют p)
Полночленные
Неполночленные (отсутствуют s)
Непол-ночлен-ные (от-сутст-вуют p, j, im, v)
нормальные с прерывистым одновершинным центрированным спектром
нормальные с прерывистым одновершинным правосторонним спектром
зрелые нормальные с максимумом на g 2
стареющие нормальные с максимумом на g3
|
Непол-ночлен-ные (от-сутст-вуют p, s) |
нормальные с полночленным двувершинным левосторонним спектром |
нормальные с прерывистым одновершинным центрированным спектром |
молодые нормальные с максимумом на v |
зрелые нормальные с максимумом на g 2 |
|
Непол- |
нормаль- |
нормаль- |
старею- |
старею- |
|
ночлен- |
ные с пре- |
ные с пре- |
щие нор- |
щие нор- |
|
ные (от- |
рывистым |
рывистым |
мальные с |
мальные |
|
сутст- |
одновер- |
одновер- |
максиму- |
с макси- |
|
вуют p, |
шинным |
шинным |
мом на g 3 |
мумом |
|
j, im, v) |
правосторонним спектром |
правосторонним спектром |
на g 3 |
|
Laser trilobum (L.) Borkh. Лазурник трехлопастной |
35 |
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с полночленным одновершинным правосторонним спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
стареющие нормальные с максимумом на g 3 |
стареющие нормальные с максимумом на g 3 |
||||||||||||||
|
30 25 20 15 10 5 0 |
. п . „ |
||||||||||||||||||||
|
1_____1 оптимум —•—стресс |
|||||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||||
|
Oxytropis flo-ribunda ( Pall.) DC. Остролодочник яркоцветный |
45 |
Неполночленные (отсутствуют s) |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с прерывистым одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
||||||||||||||
|
40 35 30 25 20 15 10 5 0 |
|||||||||||||||||||||
|
1_____1 оптимум —•—стресс |
|||||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||||
|
Oxytropis hip-polyti Boriss. Остролодочник Ипполи та |
35 |
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с полночленным двувершинным правосторонним спектром |
нормальные с прерывистым одновершинным центрированным спектром |
стареющие нормальные с максимумом на g 3 |
зрелые нормальные с максимумом на g 2 |
||||||||||||||
|
30 25 20 15 10 5 0 |
|||||||||||||||||||||
|
1_____1 оптимум ^^стресс |
|||||||||||||||||||||
|
p j im v g1 g2 g3 ss s |
|||||||||||||||||||||
|
Oxytropis spi-cata ( Pall.) O. et B. Fedtsch. Остролодочник колосистый |
40 |
Неполночленные (отсутствуют s) |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с прерывистым одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
|||||||||||||
|
35 30 25 20 15 10 5 0 |
||||||||||||||||||||
|
। _____ । оптимум —•—стресс |
||||||||||||||||||||
|
p j im v g1 g2 g3 |
ss s |
|||||||||||||||||||
|
Polygala si-birica L. Истод сибирский |
45 |
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
|||||||||||||
|
40 35 30 25 20 15 10 5 0 |
||||||||||||||||||||
|
। _____ । оптимум стресс |
||||||||||||||||||||
|
" IP™ |
||||||||||||||||||||
|
p j im v g1 g2 g3 |
ss s |
|||||||||||||||||||
|
Tanacetum achilleifolium (Bieb.) Sch. Bip. Пижма тысяче-листниковая |
40 |
Полночленные |
Непол-ночлен-ные (от-сутст-вуют p, j, v, s) |
нормальные с полночленным одновершинным центрированным спектром |
нормальные с прерывистым одновершинным правосторонним спектром |
зрелые нормальные с максимумом на g 2 |
стареющие нормальные с максимумом на g 3 |
|||||||||||||
|
35 30 25 20 15 10 5 0 |
||||||||||||||||||||
|
। — । оптимум —ф— стресс |
||||||||||||||||||||
|
। i.i—1.1 ;, |
||||||||||||||||||||
|
p j im v g1 g2 g3 |
ss s |
|||||||||||||||||||
Популяции Astragalus ucrainicus стареющие нормальные полночленные с одновершинным правосторонним спектром с максимумом на старовозрастных генеративных экземплярах (33%). Доля генеративной фракции - примерно 74%. В стрессовых условиях они также стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (37%). Отсутствуют проростки, ювенильные и имматурные группы особей. Генеративная часть популяции составляет 70-75%. Велико содержание сенильных растений - их число увеличивается с 12 до 20%. Число обследованных популяций также невелико, все они расположены близко территориально (Сыртовое Заволжье).
Для Astragalus wolgensis в оптимальных условиях популяции зрелые нормальные полночленные с двувершинным спектром с максимумом на зрелых генеративных экземплярах (30%). Второй пик отмечается на виргинильных особях (24%). Генеративное ядро представлено 61% зарегистрированных экземпляров. При экстремальном воздействии антропогенных факторов (перевыпас) популяции характеризуются как стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (29%). Отсутствуют проростки, ювенильные и сенильные растения. Доля виргинильных особей снижается в два раза (11%). Также в два раза уменьшается число всех особей предгенеративного периода. Доля генеративных, напротив, закономерно возрастает до 71%.
У Astragalus zingeri в оптимальных условиях отмечены зрелые нормальные неполночленные популяции с одновершинным центрированным спектром с максимумом на зрелой генеративной группе особей (34%). Отсутствуют сенильные растения. Доля генеративной группы превышает 70%. С возрастанием нагрузки на местообитания (перевыпас, пожоги) популяции переходят в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных экземплярах (39%). Онтогенетический спектр значительно смещается в правую сторону, отсутствуют проростки и ювенильные экземпляры. Генеративное ядро популяций составляет 77%. Субсенильные и сенильные растения в составе популяций в сумме превышают 18%.
Популяции Atraphaxis frutescens в климаксовых сообществах каменистых степей характеризуются как стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (34%). В них отсутствуют сенильные растения. Генеративное ядро составляет 68%. Незначительный пик в спектре отмечается на виргинильных особях (12%). При действии частых пожаров и перевыпаса популяции зрелые нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на зрелых генеративных экземплярах (36%). В них отсутствуют проростки, ювенильные и имматурные экземпляры. Генеративные особи в сумме составляют почти 72%.
Chrysocyathus volgensis в условиях низкой нагрузки или ее отсутствии отмечается в молодых нормальных неполночленных с прерывистым двувершинным левосторонним спектром с максимумом на виргинильной фракции (33%). Субдоминируют зрелые генеративные особи (30%). Отсутствуют сенильные астения. Вклад генеративной фракции в состав популяции адониса - чуть более 60%. С возрастанием нагрузки на местообитания (перевыпас, пожоги) популяции переходят в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спек- тром с максимумом на старовозрастных генеративных экземплярах. Отсутствуют проростки и ювенильные растения. Генеративные особи насчитывают почти 70% от общего числа зарегистрированных экземпляров.
Популяции Clausia aprica в оптимальных условиях являются зрелыми нормальными неполночленными с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (33%). Значительную роль в составе популяций играют молодые генеративные растения, занимающие субдоминирующее положение (около 30%). В спектре отсутствуют ювенильные и сенильные особи. Доля генеративных экземпляров превышает 80%. При антропогенном прессе популяции переходят в зрелые нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на зрелых генеративных особях (42%). Спектр смещается вправо, в нем отсутствуют проростки и ювенильные растения, доля виргинильных особей снижается с 17 до 6%, на субдоминирующие позиции выходят старовозрастные генеративные особи (почти 32%), значительно увеличивается число сенильных особей (с 1 до 10%). Общее число генеративных растений в популяциях достигает 82%, что мало отличается от первого случая, однако соотношение фракций здесь меняется в сторону стареющих особей.
Для Ferula caspica в оптимальных условиях существования свойственны молодые нормальные неполночленные популяции с прерывистым двувершинным левосторонним спектром с максимумом на виргинильной группе (37%). В спектре отсутствуют ювенильные экземпляры. Субдоминируют зрелые генеративные особи (29%), доля же всех генеративных растений составляет 52%. Стрессовые факторы вызывают переход к стареющим нормальным неполночленным популяциям с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (35%). В спектре нет проростков, ювенильных и сенильных особей. Значителен вклад в состав популяций генеративной фракции особей (83%).
Популяции Ferula tatarica в оптимальных условиях характеризуются как молодые нормальные неполночленные с прерывистым двувершинным спектром с максимумом на виргинильных особях (28%). Субдоминируют в составе популяций старовозрастные генеративные растения (26%), а вся генеративная фракция насчитывает 62% особей. Отсутствуют проростки и сенильные растения. При действии антропогенных факторов (пожоги) они переходят в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (36%), на втором месте также находятся зрелые генеративные особи (34%). Доля генеративной группы особей около 81%. В спектре отсутствуют проростки, ювенильные и имматурные экземпляры. Почти в два раза возрастает доля особей сенильного периода.
Galatella angustissima в условиях низкой нагрузки или ее отсутствии отмечается в составе зрелых нормальных полночленных популяциях с одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (46%). Генеративные особи составляют около 71% от численности популяции. В стрессовых условиях (перевыпас, пожоги) популяции стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (27%). Значителен вклад зрелых генеративных растений (25%), особи генеративного периода онтогенеза составляют 68% от общей численности. В спектре отсутствуют проростки и ювенильные растения. Более чем в три раза увеличивается процент сенильных особей - с 3 до 11%.
У Gentiana pneumonanthe в оптимуме популяции характеризуются как зрелые нормальные неполночленные с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных особях (42%). Примерно по 20% приходятся на виргинильные и старовозрастные генеративные особи. Доля генеративной фракции 75%. Отсутствуют в спектрах проростки и ювенильные растения. В стрессовых условиях (перевыпас, сенокошение) популяции горечавки переходят в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (около 40%). Генеративные особи в сумме составляют также около 75% популяции. Не зарегистрированы проростки и ювенильные экземпляры.
У Hedysarum gmelinii в условиях низкой нагрузки или ее отсутствии отмечаются зрелые нормальные полночленные популяции с одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (46%). Доля генеративных особей составляет 71%. При возрастании нагрузки на местообитания (степные палы, перевыпас) популяции переходят в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (31%). Субдоминируют зрелые генеративные экземпляры (29%), генеративное ядро популяции сложено примерно 73% особей. В спектре отсутствуют проростки и ювенильные растения.
В оптимуме популяции Hedysarum grandiflorum являются зрелыми нормальными полночленными с одновершинным центрированным спектром с максимумом на зрелых генеративных особях (43%). Генеративные фракции представлены 64% зарегистрированных особей. При значительной антропогенной нагрузке (перевыпас, степные палы, рекреация) они переходят в разряд стареющих нормальных неполночленных с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (36%). В популяциях отсутствуют проростки, ювенильные и сенильные растения. Доля генеративных особей составляет более 80%. Сенильная группа возрастает более чем в два раза с 3 до 7%.
Для Hedysarum razoumovianum в оптимальных условиях свойственны зрелые нормальные полночленные популяции с одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (46%). Вклад генеративных особей в состав популяций значителен – около 73%. Стареющие нормальные неполночленные популяции с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (36%) отмечаются в стрессовых условиях среды (пожоги, перевыпас). Субдоминируют зрелые генеративные особи (25%), а доля генеративной фракции в составе популяции более 81%. В таких популяциях отсутствуют проростки и ювенильные растения. Почти вдвое увеличивается число сенильных особей – с 5 до 8%.
У Iris pumila в условиях низкой нагрузки или ее отсутствии отмечаются зрелые нормальные фрагментарные популяции с одновершинным центрированным спектром с максимумом на зрелых генеративных особях (53%). Генеративная составляющая популяции – около 87%. В спектре отсутствуют проростки, ювенильные, имматурные и сенильные растения. В стрессовых условиях популяции касатика характеризуются как стареющие нормальные фрагментарные с одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (36%). Немногим меньше в популяции зрелых генеративных (35%) и молодых генеративных (25%) особей. Онтогенетический спектр еще больше смещается вправо за счет отсутствия также и виргинильных экземпляров, кроме того в два раза увеличивается число сениль- ных растений с 2 до 4%. Фрагментарность спектра популяций касатика обусловлена особенностями жизненной формы и размножения.
Jurinea ewersmannii в условиях низкой нагрузки или ее отсутствии отмечается в составе зрелых нормальных неполночленных популяций с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (32%). Доля генеративных особей в популяциях почти 84%. Нередко в популяциях на момент изучения отсутствуют проростки. При возрастании нагрузки на местообитания популяции переходят в стареющие нормальные неполночленные с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (41%). Генеративная фракция особей насчитывает около 86%. Спектр резко смещается вправо, в нем отсутствуют все группы предгене-ративного периода, а сенильная фракция возрастает с 5,5 до 14%.
Для Jurinea ledebourii в условиях низкой нагрузки или ее отсутствии свойственны молодые нормальные полночленные популяции с двувершинным левосторонним спектром с максимумом на виргинильных особях (32%). Субдоминируют зрелые генеративные растения (26%). Генеративное ядро популяции представлено 51% зарегистрированных экземпляров. При экстремальном воздействии антропогенных факторов (также степные пожары и перевыпас) популяции наголоватки претерпевают изменения и переходят в зрелые нормальные неполночленные с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (30%). В составе этих популяций насчитывается до 73% генеративных особей. В спектре отсутствуют проростки и сенильные особи.
У Jurinea multiflora даже в оптимальных условиях отмечены стареющие нормальные неполночленные популяции с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (30%). Дол генеративных растений составляет 67%. Отсутствует сенильная онтогенетическая группа особей. При существенном воздействии антропогенных факторов они являются также стареющими нормальными неполночленными с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (около 39%). При полном отсутствии возобновления популяциях (нет особей предгенеративного периода), ее составляет в основном генеративная группа растений – 83%. Сенильная фракция возрастает с 4 до 17%.
Популяции Laser trilobum в оптимальных условиях характеризуются как стареющие нормальные полночленные с одновершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (30%). Доля генеративные особей в популяциях составляет около 62%. Стареющие нормальные неполночленные популяции вида с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (28%) отмечаются в трансформированных растительных сообществах. В популяциях нередко отсутствуют проростки и ювенильные особи. Генеративная фракция представлена 68% зарегистрированных экземпляров. Более чем в три раза увеличивается число сенильных растений с 5 до 17%.
Природные популяции Oxytropis floribunda в условиях низкой нагрузки или ее отсутствии являются зрелыми нормальными неполночленными с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (40%). В спектрах отсутствуют сенильные растения. Доля генеративных особей составляет почти 85%. При перевыпасе и нерегулируемых палах степного травостоя у остролодочника регистрируются стареющие нормальные неполночленные популяции с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (около 34%). Обычно не фиксируются особи на ранних этапах развития – проростки и ювенильные – в связи с катастрофической гибелью. Генеративная фракция насчитывает более 75% особей обследованных популяций. В два раза и более увеличивается число сенильных экземпляров – с 3 до 7%.
Oxytropis hippolyti в оптимальных условиях отмечен в стареющих нормальных полночленных популяциях с двувершинным правосторонним спектром с максимумом на старовозрастных генеративных особях (30%). Субдоминирующее положение занимают молодые генеративные особи (около 25%). Генеративное ядро – 74%. Зрелые нормальные неполночленные популяции с прерывистым одновершинным центрированным спектром с максимумом на особях зрелой генеративной группы (29%) фиксируются при повышении антропогенного пресса. Незначительно уступают им по численности старовозрастные генеративные особи (28%), доля всех генеративных групп – более 76%. Зачастую в популяциях не фиксируются проростки и ювенильные растения. Сенильная группа увеличивается с 3 до 11%.
Популяции Oxytropis spicata в условиях низкой нагрузки или ее отсутствии характеризуются как зрелые нормальные неполночленные с прерывистым одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (30%). Почти не уступает им молодая генеративная группа особей – 27%. Генеративная фракция составляет до 76% зарегистрированных экземпляров. В спектре отсутствуют сенильные растения. В стрессовых условиях популяции являются стареющими нормальными неполночленными с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (36%). Генеративная фракция – 77%. В спектре отсутствуют проростки и ювенильные особи. Сенильная группа увеличивается с 1,5 до 7%.
Polygala sibirica в оптимальных условиях существует в зрелых нормальных полночленных популяциях с одновершинным центрированным спектром с максимумом на зрелых генеративных особях (30%). Генеративное ядро насчитывает более 70% особей. При значительной антропогенной нагрузке популяции истода становятся стареющими нормальными неполночленными с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастной генеративной группе особей (40%). Генеративная фракция составляет 81%. В спектре отсутствуют проростки и ювенильные особи. Сенильные растения не играют значимой роли, их число остается примерно на одном уровне – 4-6%.
Популяции Tanacetum achilleifolium в условиях низкой нагрузки или ее отсутствии следует считать зрелыми нормальными полночленными с одновершинным центрированным спектром с максимумом на зрелых генеративных экземплярах (36%). Генеративная фракция представлена 70% особей. Стареющие нормальные неполночленные популяции пижмы с прерывистым одновершинным правосторонним спектром с максимумом на старовозрастных генеративных растениях (31%) зарегистрированы в условиях стрессовой нагрузки на местообитания. Генеративное ядро составляют почти 79% зафиксированных особей. В спектрах отсутствуют проростки, ювенильные, имматурные и сенильные растения.
Установлено, что у большинства изученных видов растений при возрастании антропогенного воздействия на местообитания изменения в популяциях заключаются в смещении онтогенетических спектров вправо, увеличении доли генеративных и сенильных особей, почти полном отсутствии особей начальных периодов онтогенеза в связи с их катастрофической гибелью. В основном существование популяций этих представителей обусловлено не стабильностью или способностью к самовосстановлению, а лишь длительностью существования генеративных экземпляров.
Список литературы Изменения базовых онтогенетических спектров популяций некоторых редких видов растений Самарской области при антропогенной нагрузке на местообитания
- Абрамова Л.М., Баширова Р.М., Муртазина Ф.К., Усманов И.Ю. Характеристика ценопопуляций Glycyrrhiza korchinskyi Grig. на юго-востоке Республики Башкортостан//Растительные ресурсы. 2001. Т. 37, № 2. С. 24-29.
- Абрамова Л.М., Каримова О.А., Андреева И.З. Структура и состояние популяций Althaea officinalis (Malvaceae) на юге Предуралья (Республика Башкортостан)//Растительные ресурсы. 2010. Т. 46, № 4. С. 47-54.
- Атласова Л.Г. Онтогенетическое состояние и возрастная структура Oxytropis candicans и O. strobilacea в условиях Центральной Якутии//Изв. Самар. НЦ РАН. 2012. Т. 14, № 1-9. С. 2174-2177.
- Бийболатова З.А., Аджиева А.И. Онтогенетическая структура ценопопуляций эндемичного дагестансткого вида Scabiosa gumbetica Boiss.//Междунар. журн. прикладных и фундаментальных исследований. 2014. № 10-1. С. 43-47.
- Валуйских О.Е., Тетерюк Л.В. Структура и динамика краевых популяций Gymnadenia conopsea (L.) R. Br. (Orchidaceae) на известняках европейского северо-запада России//Экология. 2013. № 6. С. 420-427.
