Изменения численно-размерного состава и динамического состояния нервных волокон седалищного нерва при удлинении бедра у собак

Автор: Шевцов В.И., Щудло Н.А., Щудло М.М., Борисова И.В.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 1, 2004 года.
Бесплатный доступ
В опытах на 17 взрослых беспородных собаках, которым осуществляли остеотомию бедренной кости и нейтральный остеосинтез, а также оперативное удлинение бедра спице-стержневым аппаратом оригинальной конструкции, проведено сопоставительное изучение изменений проводниковой части седалищного нерва при костной травме и специфическом влиянии дистракции. Методами световой и трансмиссионной электронной микроскопии с применением компьютерной морфометрии полутонких срезов установлено, что через 42 и 70 дней после операции (в опытах с дистракционным остеосинтезом это - конец дистракции и 30 дней фиксации) изменения численно-размерного состава волокон седалищного нерва обусловлены главным образом увеличением количества новообразованных проводников. Интермиттирующие клинические проявления выпадения функции седалищного нерва связаны прежде всего с нарушениями аксоплазматического тока, что ультраструктурно проявляется изменениями цитоскелета осевых цилиндров мякотных и безмякотных волокон и согласуется с данными морфометрии об изменении аксоплазматического компартмента зрелых мякотных волокон.
Удлинение конечности, периферический нерв, эксперимент
Короткий адрес: https://sciup.org/142120687
IDR: 142120687
The changes of the numerical-and-size composition and dynamic condition of the sciatic nerve fibers during canine femur lengthening
Comparative study of the changes of the sciatic nerve conductive part under bone injury and specific effect of distraction was performed in the experiments on 17 adult mongrel dogs, which were subjected to femoral osteotomy and neutral osteosynthesis and also to surgical lengthening of femur with a device of original design in which wires and half-pins were used. Using the methods of light and transmission electron microscopy with computer morphometry of half-thin sections it was established that the changes of the numerical-and-size composition of the sciatic nerve fibers were mainly caused by increase of newly formed conductors in number in 42 and 70 days after surgery (this is the end of distraction and 30 days of fixation in the experiments with distraction osteosynthesis). Intermittent clinical symptoms of the descent of the sciatic nerve function first of all were associated with axoplasmatic current disorders, that was demonstrated ultrastructurally by the changes of the cytoskeletal axis cylinders of medullary nerve fibers and nonmedullated ones, and was consistent with the morphometry data of the change of the axoplasmatic compartment of mature medullary fibers.
Текст научной статьи Изменения численно-размерного состава и динамического состояния нервных волокон седалищного нерва при удлинении бедра у собак
Большинство морфологических работ, посвящённых влиянию дистракции на нервы конечностей [1, 5-7], не содержат объективных количественных данных. Г.А. Илизаровым и М.М. Щудло [2] в ультраструктурном исследовании нервов удлиняемой конечности рассматривались процессы двух типов: реакция на операционную травму и изменения нерва как органа под влиянием напряжения растяжения. Установлена когерентность изменений структурных характеристик периневрального барьера и реактивных изменений нервных волокон; на протя- жении всего периода дистракции наблюдалось постепенное новообразование тканевых структур.
Изучение морфофункциональных изменений берцовых нервов удлиняемой голени при различной дробности дистракции [3, 4] показало, что в зависимости от режима удлинения конечности в тканях периферических нервов в различной степени выражены явления деструкции, регенерации и вставочного роста. Однако в этих исследованиях не было серии опытов с повреждением костей и нейтральным остеосинтезом, что не позволило четко обозначить специфическое влияние дистракции.
Цель данного исследования – сопостави- тельное изучение адаптивных и патологических изменений проводниковой части нервов при костной травме и удлинении конечности.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование проведено на 17 взрослых беспородных собаках. Под внутривенным барбитуровым наркозом в асептических условиях выполняли открытую чрезнадкостничную поперечную остеотомию бедренной кости в средней трети пилой Джигли и остеосинтез спице-стержневым аппаратом [10]. У 7 собак костные фрагменты фиксировали в течение всего срока эксперимента (6 и 10 недель) в положении контакта (нейтральный остеосинтез). У остальных 10 собак на 7-9 день начинали дистракцию фрагментов бедренной кости по 0,25 мм 3 раза в день; их выводили из опыта через 6 недель (в конце дистракции – 3 собаки), в разные сроки фиксации и после снятия аппарата. Удлинение бедра составляло от 12 до 15% исходной длины.
Животных эвтаназировали передозировкой барбитуратов. С целью исключения аутолитических изменений взятие материала для морфологического исследования производили в течение первых 20 минут после остановки сердцебиения. Отрезки седалищного нерва привязывали к деревянной основе, затем иссекали. Материал фиксировали в охлажденной смеси 2%-ных растворов глютарового и параформальдегидов на фосфатном буфере (pH–7,4), постфиксировали четырехоки-сью осмия, заключали в эпоксидные смолы (арал-дит или эпон-аралдит) и разлагали на срезы на ультратомах фирмы “LKB” (Швеция). Полутонкие срезы окрашивали толуидиновым или метиленовым синим и кислым фуксином для светооптической микроскопии. Ультратонкие срезы контрастировали уранилацетатом, цитратом свинца и исследовали в трансмиссионном электронном микроскопе JEM-100B (“Jeol”, Япония) при ускоряющем напряжении 80 kV.
У 11 собак из 17 выполнено морфометрическое исследование полутонких срезов. Используя большой исследовательский фотомикроскоп фирмы “Opton” (Германия) и аппаратно-программный комплекс “ДиаМорф” (Москва), с каждого уровня оцифровывали от 30 до 80 полей зрения с изображениями не менее 500 мякотных нервных волокон (МНВ). Определяли численную плотность их профилей (NA мнв ), по правилу эквивалентного круга измеряли диаметры мнв (D мнв ), оценивали распределение мякотных волокон по калибрам. Отдельно вычисляли морфометрические параметры крупного калибра: максимальный (Dmaх), средний диаметр мякотных волокон (Dсредн ) и их осевых цилиндров (Axсредн), среднюю толщину миелина (L миел ), среднее число G (G средн ). Для контроля морфометрировали полутонкие срезы седалищных нервов 3 интактных собак. Проверку статистических гипотез проводили с использованием двухвыборочного t-теста, предоставляемого программой Microsoft Excel 97, и непараметрического критерия Манна-Уитни.
РЕЗУЛЬТАТЫ
При клиническом исследовании выявлена существенная разница в функциональном состоянии конечности у животных с нейтральным и дистракционным остеосинтезом. В серии с нейтральным остеосинтезом 2 собаки из 7 с первого дня после операции полноценно нагружали оперированную лапу при ходьбе и опоре в стойке, а у 5 собак опора и походка восстановились только через 2-3 недели после операции.
В опытах с дистракционным остеосинтезом в течение первых 2 недель после операции ослабление опоры отмечалось у 3 животных из 10. К концу дистракции у 6 собак периодически и ещё у 2 собак постоянно наблюдалась патологическая опора на подогнутые пальцы, что свидетельствовало о нарушении функции седалищного нерва. У некоторых собак данная симптоматика исчезла уже в начале периода фиксации, у других это происходило постепенно.
Гистоморфометрические характеристики седалищного нерва интактных собак и человека сопоставимы. Численная плотность нервных волокон в доминантных пучках варьировала от 9 до 12 тыс. на 1 мм2. Распределение их по калибру унимодально, причём мелких волокон (<4 мкм диаметром) не более 25%, средних (4,1-7,0 мкм) – не более 40%, а крупных (>7 мкм) – от 35 до 60%. Самые крупные мякотные волокна имеют диаметр 14-16 мкм, единичные превышают 18 мкм в диаметре, но встречаются они не у всех индивидов. Число G в разных фракциях колеблется от 0,69 до 0,76. Процент мякотных волокон с признаками реактивно-деструктивных изменений составляет от 0,6 до 1,8%, причём среди изменённых волокон преобладают волокна крупного калибра.
Таблица 1
Морфометрические характеристики мякотных нервных волокон в опытах с нейтральным и дистракционным остеосинтезом
|
№ |
Опыт |
NA мнв (M ± m) |
Процент дегенеративно изменённых форм |
Распределение (в процентах) мякотных волокон по калибрам (мкм) |
||
|
1,1-4,0 |
4,1-7,0 |
7,1 и более |
||||
|
1. |
D1 |
17385±1438 |
1,2 |
30,6 |
24,3 |
45,1 |
|
2. |
D2 |
17464±868 |
1,2 |
25,3 |
26,3 |
48,4 |
|
3. |
F30-1 |
22833±1485 |
1,4 |
39,8 |
24,1 |
36,1 |
|
4. |
F30-2 |
18667±941 |
0,98 |
33,6 |
24,0 |
42,4 |
|
5. |
F30-3 |
20600±1196 |
1,8 |
30,4 |
29,7 |
39,9 |
|
6. |
F30-4 |
18933±936 |
0,5 |
28,7 |
30,9 |
40,4 |
|
7. |
F30-5* |
17400±1474 |
7,8 |
29,3 |
40,6 |
30,1 |
|
8. |
Neutr1 |
13167±800 |
1,2 |
18,2 |
21,6 |
60,2 |
|
9. |
Neutr2 |
17793±819 |
0,98 |
24,9 |
25,8 |
45,3 |
|
10. |
Neutr3 |
10829±653 |
2,9 |
16,2 |
22,9 |
60,9 |
|
11. |
Neutr4 |
17267±1154 |
0,2 |
26,5 |
27,6 |
45,9 |
Данные таблицы 1 свидетельствуют, что в экспериментах с остеотомией бедренной кости численная плотность мякотных волокон возрастает в сравнении с интактными нервами, причём если при нейтральном остеосинтезе (Neutr1-4) через 10 недель после операции она увеличена на 40%, то к концу дистракции (D1 и D2 – 6 недель после операции) – на 60%, а через месяц фиксации (F30-1-5 – те же 10 недель после операции) – почти на 90%.
Доля волокон с признаками реактивнодеструктивных изменений оказалась значительно выше, чем в интактных нервах, только в одном эксперименте (F30-5*). Причиной, по-видимому, следует считать ишемическую нейропатию, развившуюся в связи с эмболизацией центральной артерии одного из пучков седалищного нерва.
Распределение мякотных волокон по калибру в опытах с нейтральным остеосинтезом практически не отличается от интактных нервов, а при дистракции бедренной кости доля мелких волокон в седалищном нерве увеличивается, а крупных – уменьшается.
При нейтральном остеосинтезе в большинстве смешанных пучков нерва отношение числа безмякотных волокон к мякотным не достигает единицы или весьма незначительно её превышает, а при дистракции данный показатель увеличивается до 1,3. Среди безмякотных волокон в удлинённом нерве довольно редко встречаются «розетты». Повышено количество незрелых аксо-глиальных комплексов, многие из которых содержат тонкие миелинизирующиеся волокна.
Появление большого количества новообразованных мелких волокон – основная причина изменения распределения мякотных волокон по калибру, то есть увеличение доли мелких волокон обусловлено абсолютным приростом их численности. За счёт прироста численности мелких мякотных волокон процентная доля деструктивно изменённых волокон весьма невелика. Однако вряд ли следует считать, что уменьшение доли крупных волокон только относи- тельно. К абсолютному снижению числа крупных волокон приводит потеря некоторого их количества в результате аксональной дегенерации, а также перекалибровка волокон.
При дистракционном остеосинтезе чаще, чем в норме и в условиях нейтрального остеосинтеза, встречаются осевые цилиндры с конденсацией элементов цитоскелета, выраженной в разной степени (рис. 1), иногда фокальной. Особенно заметна конденсация нейрофиламентов в некоторых крупных двигательных мякотных волокнах. Именно эти волокна имеют уменьшенное число G в результате истончения осевого цилиндра. Однако среди них не обнаружено ни одного, которое имело бы G меньше 0,6, поэтому говорить о развитии аксональной атрофии нет оснований.
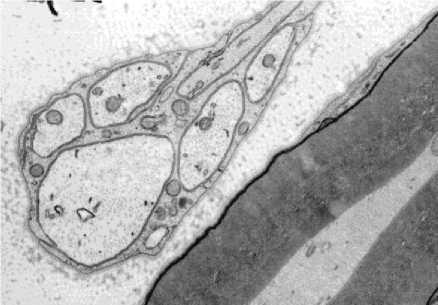
Рис. 1. Различная плотность расположения элементов цитоскелета в осевых цилиндрах нервных волокон удлинённого седалищного нерва собаки. Элек-тронограмма, 100000х
В таблице 2 представлены морфометрические характеристики волокон крупного (более 7 мкм в диаметре) калибра. Через 10 недель после операции эта фракция не содержит новообразованных или регенерировавших волокон, поэтому такое сопоставление даёт возможность судить о влиянии условий эксперимента на зрелые мякотные волокна.
Таблица 2
Морфометрические характеристики фракции крупных волокон (7 и более мкм)
|
№ |
Опыт |
D maх |
D средн |
Ах x средн |
L миел |
G средн |
|
1. |
Neutr1 |
15,3 |
10,62±0,10 |
5,50±0,08 |
2,56±0,02 |
0,756±0,002 |
|
2. |
Neutr2 |
16,2 |
10,43±0,13 |
4,94±0,09 |
2,75±0,02 |
0,734±0,002 |
|
3. |
Neutr3 |
17,9 |
11,24±0,14 |
5,23±0,09 |
3,00±0,02 |
0,731±0,002 |
|
4. |
Neutr4 |
16,7 |
10,68±0,14 |
5,59±0,10 |
2,55±0,03 |
0,759±0,002 |
|
5. |
Intact1 |
14,5 |
10,00±0,12 |
5,28±0,09 |
2,36±0,02 |
0,760±0,003 |
|
6. |
Intact2 |
18,9 |
11,18±0,13 |
5,69±0,11 |
2,79±0,02 |
0,734±0,003 |
|
7. |
Intact3 |
15,8 |
9,99±0,14 |
4,74±0,11 |
2,63±0,02 |
0,730±0,003 |
|
8. |
D-1 |
15,4 |
10,63±0,14 |
5,98±0,11 |
2,33±0,03 |
0,779±0,002 |
|
9. |
D-2 |
13,5 |
9,32±0,09 |
4,15±0,11 |
2,58±0,02 |
0,721±0,002 |
|
5. |
F30-1 |
16,0 |
9,85±0,13 |
4,54±0,11 |
2,65±0,02 |
0,727±0,003 |
|
6. |
F30-2 |
16,6 |
10,68±0,14 |
5,25±0,10 |
2,71±0,03 |
0,742±0,002 |
|
7. |
F30-3 |
15,2 |
9,33±0,12 |
4,45±0,09 |
2,44±0,02 |
0,736±0,002 |
|
8. |
F30-4 |
15,4 |
9,59±0,12 |
4,38±0,09 |
2,61±0,02 |
0,726±0,002 |
|
9. |
F30-5* |
17,3 |
9,60±0,12 |
4,44±0,11 |
2,58±0,02 |
0,729±0,003 |
Сравнение максимальных диаметров позволяет утверждать, что вариативность крупных мякотных волокон в условиях опытов не меняется, популяция в целом сохраняет свою гетерогенность. В конце дистракции при практически нормальной средней толщине миелина отмечаются отклонения параметра «средний диаметр осевого цилиндра» от интактной нормы как в сторону увеличения (D-1), так и уменьшения (D-2). Через 30 дней фиксации (F30-1-5) в серии с дистракционным остеосинтезом по сравнению с соответствующим сроком нейтрального остео- синтеза статистически незначимо уменьшены средняя толщина миелина и среднее G, но достоверно ниже средние диаметры крупных мякотных волокон и их осевых цилиндров.
По среднему диаметру мякотных волокон крупного калибра и среднему диаметру их осевых цилиндров все 4 варианты серии с нейтральным остеосинтезом попадают в диапазон интактных нервов (Intact 1-3). В серии с дистракционным остеосинтезом эти параметры снижены по сравнению с интактными в 4 опытах из 5.
ОБСУЖДЕНИЕ
Результаты выполненного исследования свидетельствуют, что в популяции нервных волокон седалищного нерва собак протекают интенсивные перестроечные процессы не только при удлинении бедра на 12-15%, но и в ответ на остеотомию бедренной кости. Доля деструктивно изменённых мякотных волокон варьирует от 0,2 до 2,9%, однако происходит интенсивное, по принципу избыточности, новообразование нервных проводников. Выяснение биологического значения и механизма этого феномена должно стать предметом специальных исследований.
Появление незрелых аксо-глиальных комплексов и увеличение численной плотности безмякотных и мякотных волокон в большей мере характерно для экспериментов с дистракционным остеосинтезом. В частности, численная плотность мякотных волокон по сравнению с интактными нервами при дистракционном остеосинтезе увеличивается почти вдвое. Это свидетельствует прежде всего о высокой биосинтетической и секреторной активности органелл в клеточных телах нейронов, которые увеличивают количество ветвлений своих отростков, а возможно, образуют и новые отростки.
Сохранение ультраструктуры большинства зрелых нервных волокон в процессе дробного удлинения голени собак с суточным темпом 1,0 мм на 15 и 30% исходной длины отмечено в исследовании М.М. Щудло [9]. K. Ikeda et al. [14] удлиняли бедро кроликов также на 30%, но в других режимах: по 0,8; 2 и 4 мм в сутки за один приём. При дистракции по 0,8 мм в день авторы не выявили функциональных нарушений. При удлинении по 2 мм в день регистрировалось снижение амплитуды SSAP и увеличение латентного периода, а при темпе 4 мм – полный блок проведения и ретроградного аксонального транспорта, исследованного с помощью реакции агглютинина зародышей пшеницы и пероксидазы хрена (WGA-HRP). Однако даже при таком глубоком нарушении функции авторы не обнаружили дегенеративных изменений нервных волокон при электронномикроскопическом исследовании, поэтому предложили обозначить данный вид повреждения нерва как «аксонолептостазис», понимая под этим термином стагнацию аксонального транспорта.
Известно, что благодаря аксональному транспорту нейроны поддерживают свою жизнедеятельность, а также выживание и функционирование периферических отростков, не имеющих ни рибосом, ни комплекса Гольджи, но простирающихся на большие расстояния (метр и более), в сотни и тысячи раз превышающие размеры клеточных тел.
Отмеченные в нашем исследовании интермиттирующие клинические проявления нарушения функции седалищного нерва при удлинении бедра собак возможно также вызваны изменениями аксонального транспорта, о чём свидетельствуют фокальные аккумуляции и разрежения элементов цитоскелета в некоторых осевых цилиндрах, а также выраженная конденсация нейрофиламентов в части крупных двигательных волокон. Эти ульт-раструктурные изменения детерминируют регистрируемые при морфометрии полутонких срезов изменения среднего диаметра нервных волокон и их осевых цилиндров. То, что именно состояние нейрофиламентных протеинов определяет калибр аксонов, в литературе считается чётко установленным [15]. С другой стороны, эти же авторы при исследовании морфологически изменённых и интактных волокон седалищного нерва трансгенных мышей интрааксональными электродами обнаружили, что при повреждении генов NF-H и NF-M-протеинов нейрофиламентов даже в волокнах сохранённого крупного калибра скорость проведения снижалась за счёт удлинения рефрактерного периода потенциала действия. Эти результаты позволяют предположить, что нейрофиламенты играют специфическую роль в модуляции функций ионных каналов.
Известно, что в норме средняя суточная скорость транспорта цитоскелетных полимеров и ассоциированных протеинов варьирует от 0,3 до 3 мм в день [11]; в хрестоматийных схемах это перемещение обозначается как самый медленный компонент антероградного транспорта. Однако исследования последних лет позволили уточнить, что оно включает быстрые, нечастые и высокоасинхронные антероградные движения, но 20-30% нейрофиламентов подвергается ретроградным забросам. В связи с этим представляется обоснованным предположение, что не только при развитии нейропатий, но и в условиях дистракционного остеосинтеза (то есть, при механическом, хотя и дозированном, растяжении мягких тканей и разнообразных гомеостатических сдвигах, сопровождающих повреждение и новообразование кости) именно этот сложный процесс оказывается самым уязвимым.
ЗАКЛЮЧЕНИЕ
Проведённые эксперименты позволили получить объективные количественные данные, свидетельствующие о преобладании процесса новообразования нервных волокон над деструктивными изменениями при удлинении бедра собак с суточным темпом 0,75 мм за 3 приёма. Наблюдавшиеся в ходе эксперимента обратимые и эпизодические расстройства функции седалищного нерва, по-видимому, вызваны изменениями динамического состояния нервных волокон – нарушениями аксонального транспорта и ионной проницаемости. В этой связи представляются вполне оправданными планомерное изучение и внедрение в практику высокодробных режимов дистракции [8]. При разработке методов профилактики и коррекции подобных нарушений следует также иметь в виду, что аксональный транспорт сопровождается разрушением АТФ, является кальций- и кислород-зависимым процессом, то есть может страдать при соответствующих дефицитных состояниях.
