Изменения сообществ раковинных амеб, обитающих в эпилитных мхах, вдоль высотного градиента (Прибайкалье, хребет Хамар-Дабан)
")
Автор: Мазей Ю.А., Марфина О.В., Белякова О.И., Чернышов В.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.13, 2011 года.
Бесплатный доступ
Изучены закономерности изменения сообществ раковинных амеб из эпилитных мхов вдоль высотного градиента на склоне пика Черского (хребет Хамар-Дабан, Прибайкалье) в диапазоне высот 1000-1800 м над уровнем моря в соответствии со сменой биогеоценозов от березово-осинового леса через кедрово-пихтовый лес, кедрово-пихтово-кустраниковое лесотундровое редколесье к мохово-лишайниковой тундре. Обнаружено 49 видов и форм раковинных корненожек. Наиболее характерный элемент фауны - убиквитные виды с широким географическим распространением: Centropyxis aerophila, Phryganella acropodia, Trinema enchelys, Corythion dubium, Assulina seminulum. Видовое богатство и численность раковинок выше в поясе мохово-лишайниковой тундры. С высотой возрастает пространственная гетерогенность видового состава и изменяется структура доминирующего комплекса видов. При этом многие формы не проявляют направленных тенденций в изменении, как численности, так и относительной представленности в структуре сообщества. Подчеркивается отсутствие единой тенденции в ценотических изменениях сообществ раковинных амеб вдоль высотных градиентов в Западной Европе и Прибайкалье.
Раковинные амебы, структура сообщества, эпилитные мхи, высотный градиента, прибайкалье, хребет хамар-дабан, пик черского
Короткий адрес: https://sciup.org/148200309
IDR: 148200309 | УДК: 593:1
Changes of testate amoebae communities from epilytic mosses along the altitudinal gradient (Baikal lake area, Khamar-Daban mountain ridge)
Patterns of testate amoebae community structure changes along the latitudinal gradient were studied along the slope of Chersky peak (Khamar-Daban mountain ridge, Baikal Lake region). In the range of altitudes from 1000 till 1800 meters above the sea level several ecosystems replaced named birch-aspen forest, cedar-fir forest, cedar-firhoneysuckle forest-tundra, moss-lichen tundra. Testate amoebae inhabited moss covering large boulders were investigated. 49 species and forms were identified. Most common species are ubiquists with wide geographical distribution: Centropyxis aerophila, Phryganella acropodia, Trinema enchelys, Corythion dubium, Assulina seminulum. Species richness and abundance maximal is in the moss-lichen tundra. Along the altitudinal gradient towards the top spatial heterogeneity grows and community structure changes. However, many species do not show directional patterns along the mountain slope. It is underlined that there are no common patterns of altitudinal community changes of testate amoebae in western Europe and Baikal Lake region.
Текст научной статьи Изменения сообществ раковинных амеб, обитающих в эпилитных мхах, вдоль высотного градиента (Прибайкалье, хребет Хамар-Дабан)
Р аковинные корненожки – полифилетическая группа протистов, включающая лобозных и филоз-ных амеб [10, 17]. Эти организмы объединяет возможность применения одной и той же техники исследования и сходная экология, что обуславливает их изучение в рамках единого методологического подхода – ризоподного анализа [4]. Раковинные амебы широко распространены в почвах, пресных водах и болотах [25]. Возможность обитания этих организмов в наземных мхах лимитируется увлажненностью [24]. При этом раковинки здесь обычно более мелкие и сплющенные, что позволяет им выживать в тонких водных пленках, покрывающих поверхность мхов [9, 29]. Боннэ [24] разделял население более сухих эпифитных мхов с характерными убиквитными и космополитными видами Cory-thion dubium, Assulina muscorum и фауну собственно эпигейных (напочвенных) мхов, в которую значимый вклад вносят почвенно-подстилочные формы Plagiopyxis spp., Centropyxis spp. Об особенностях ризоподного населения эпилитных мхов (растущих обычно в виде моховых подушках на валунах и скалах) данные до настоящего времени отсутствуют.
Неравномерно исследована также почвенная протистофауна на территории Российской Федерации. Основные работы были проведены в пределах
Восточно-Европейской равнины [1–3, 5, 7, 8, 11–16, 18,19]. Значительно реже приводятся данные по Западно-Сибирской равнине [6, 20]. Что касается территории Прибайкалья, то она исследована крайне фрагментарно. Существует только одна работа [23], в которой приводятся список из 53 видов и описания некоторых форм раковинных корненожек из пяти точек (проб), отобранных в районе устья реки Ангары и в Байкальском заповеднике. Таким образом, байкальский регион до сих пор остается «протозоологическим белым пятном», что определяет актуальность любой информации о населении простейших, полученной здесь.
Наконец, помимо сугубо протозоологических задач выявления видового состава и структуры сообществ в пределах слабо изученной территории (Прибайкалье) и на необычных субстратах (эпи-литные мхи), настоящая работа нацелена на выявление закономерностей изменения сообществ вдоль высотного градиента. В последнее время становится очевидным, что широко известные макроэколо-гические закономерности (например, широтная зональность, высотная поясность и т.п.), выявленные на организмах крупного размера, не всегда проявляются на уровне микроскопических объектов [22]. Для раковинных корненожек этот феномен достаточно хорошо понятен в случае широтных градиентов [28, 30, 32], однако информация о роли высотного градиента крайне незначительна, чтобы делать определенные заключения. Так, существуют лишь две работы, в которых анализировался подобный вопрос. Тодоров [31] установил, что видовое разнообразие почвообитающих раковинных амеб уменьшается в субальпийской зоне (2000–2500 м н.у.м.) по сравнению с более низкими высотами (400–2000 м н.у.м.) на горе Пирин в Болгарии. В другой работе [27] было отмечено отсут- ствие существенных изменений разнообразия и структуры сообщества раковинных амеб, населяющих эпигейные мхи Hylocomium splendens, в пределах высот 1000-2200 м н.у.м. в юго-восточных Альпах (Италия). В задачи настоящей работы входило выяснение того, как меняются сообщества раковинных корненожек, населяющих подушки эпилитных мхов на склоне пика Черского (хребет Хамар-Дабан) на высотах от 1000 до 1800 м н.у.м.
МАТЕРИАЛ И МЕТОДЫ
Материал был собран в июле 2008 г. в разнотипных биогеоценозах, сменяющих друг друга вдоль высотного градиента (1000-1800 м н.у.м.) вдоль пика Черского (хребет Хамар-Дабан, Прибайкалье): 1) осинник разнотравный с примесью березы (7С3Б) на высоте 990 м н.у.м; 2) участок темнохвойной тайги - кедрово-пихтовый лес зеле-номошник (6П4К) на высоте 1340 м н.у.м.; 3) участок разреженного лесотундрового биогеоценоза с преобладанием невысоких деревьев пихты, кедра, а также кустарников жимолости на высоте 1570 м н.у.м.; 4) мохово-лишайниковая тундра на высоте 1830 м н.у.м. В каждом биогеоценозе были выбраны по три крупных валуна (высотой 2-3 м), на которых произрастали мхи. В пределах каждого валуна пробы были отобраны из моховых подушек на уровне 10-20 см, 100-120 см и 200-220 см над поверхностью земли. Таким образом, из каждого биогеоценоза проанализировано 9 образцов. Пробы отбирались и просматривались по стандартной методике [14]. Для выявления связи между различия -ми локальных ценозов и видовой структурой проводили ординацию сообществ методом анализа главных компонент на основе величин относительных обилий доминирующих (более 5 % от общей численности) видов. Для классификации локальных сообществ по видовому составу осуществляли кластерный анализ методом среднего присоединения на основе матрицы индексов сходства Раупа-Крика. Достоверность различий в величинах видового богатства и численности между биотопами оценивали при помощи критерия Манна-Уитни с поправкой Бонферрони для множественных сравнений. Расчеты вели при помощи пакета программ PAST 1.89.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В изученных биотопах обнаружено 49 видов и форм раковинных амеб: Arcella arenaria, A. a. compressa, A. a. sphagnicola, Assulina muscorum, A. seminulum, Centropyxis aculeata, C. aerophila, C. a. shagnicola, C. constricta, C. orbicularis, C. platystoma, C. sylvatica, Corythion dubium, C. orbicularis, Difflugia penardi, Euglypha acantophora, E. anodonta magna, E. capsiosa, E. ciliata, E. c. glabra, Euglypha compressa, E. cristata decora, E. c. major, E.denticulata, E. laevis, E. marginata, E.rotunda, E. simplex, E. strigosa, E. s. glabra, E. tuberculata,
Heleopera sphagni, H. sylvatica, Nebela collaris, N. galeata, N. militaris, N. parvula, N. tincta, Phryganella acropodia, Ph. hemisphaerica, Plagiopyxis declivis, Tracheleuglypha dentata, Trigonopyxis arcula, Trinema complanatum, T. enchelys, T. e. biconvexa, T. galeata, T. lineare, T. penardi .
Наиболее часто встречались убиквитные виды с широким географическим распространением Centropyxis aerophila (обнаружен в 72% микробиотопов), Phryganella acropodia (69%), Trinema enchelys (63%), Corythion dubium (56%), Assulina seminulum (50%), Phryganella hemisphaerica (43%), а также броибионт Nebela tincta (43%). В предыдущей работе, проведенной в Прибайкалье [23], 4 вида были встречены более чем в половине проанализированных проб: Trinema lineare (100%), Trinema com-planatum (60%), Phryganella acropodia (60%), Assu-lina muscorum (60%). Все они также широко распространенные эврибионты. В эпигейных мхах в Альпах [27] также наиболее обычными формами были Assulina muscorum (100%), Corythion dubium (100%), Euglypha ciliata (100%), Phryganella acro-podia (95%), Euglypha laevis (95%), Centropyxis aerophila (86%), Nebela tincta (81%). Подобная же ситуация складывается и в горах Пирин в Болгарии [31]. Следовательно, преобладание убиквистов, вероятно, является характерной чертой горных тер-
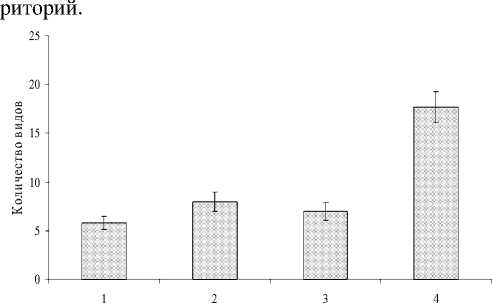
Рис. 1. Изменение среднего видового богатства в локальных сообществах раковинных амеб с высотой:
1 - березово-осиновый лес, 2 - темнохвоная тайга, 3 -лесотундра, 4 - тундра. Планки погрешностей -ошибка средней
Видовое богатство в локальных сообществах варьирует в широких пределах (от 2 до 25 видов в пробе). При этом в мохово-лишайниковой тундре этот показатель достоверно (р<0.05) выше, чем в остальных биотопах (рис. 1). Полученные данные не согласуются с имевшимися ранее представлениями. Так, в горах Пирин [31] отмечена противоположная тенденция уменьшения разнообразия с высотой, а в Альпах [27] достоверных направленных изменения выявлено не было. Все это указывает на то, что закономерности изменений связаны не столько с высотой над уровнем моря как таковой, сколько с конкретными биогеоценотическими фак- 135
торами, определяющими специфику высотного градиента структуры протозойных сообществ в каждом из этих вариантов.
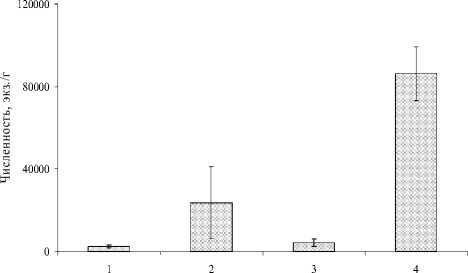
Рис. 2. Изменение обилия раковинных амеб с высотой:
1 – березово-осиновый лес, 2 – темнохвоная тайга, 3 – лесотундра, 4 – тундра.
Планки погрешностей – ошибка средней

0,8
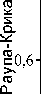
ф ct
0,2
Рис. 3. Результаты классификации локальных сообществ раковинных амеб по видовому составу:
1–4 – различные биогеоценозы (см. рис. 1), a–c – разные валуны в пределах биогеоценоза
Обилие организмов также достоверно выше в эпилитных мхах в условиях тундры на высоте 1800 м н.у.м. (рис. 2), где в отдельных образцах может достигать 150 тыс. экз./г абсолютно сухого субстрата. Минимальные показатели численности (1 тыс. экз./г) отмечены на верхушках валунов в поясе лиственных лесов и лесотундры. В темнохвойной тайге обилие варьирует в широких пределах – от 4 тыс. до 120 тыс. экз/г. В разнотипных почвенных и моховых биотопах Прибайкалья [23] численность раковинок находилась в пределах 9–30 тыс. экз./г и не столь сильно различалась от пробы к пробе как в случае эпилитных мхов. Средние величины обилий корненожек в Альпах [27] варьировали еще меньше и находились в пределах 24–31 тыс. экз./г. По всей видимости, эпилитные биотопы более гетерогенны, чем напочвенные, что и приводит к существенной разности в показателях численности формирующихся локальных сообществ в разных микробиотопах.
Локальные сообщества из тундры также наиболее значительно отличаются от других по видовому составу (рис. 3). Здесь обнаружено 13 видов, не встречающихся на более низких высотах. Среди них значительное количество мелких эврибионт-ных видов и форм из рода Euglypha , а также гигрофильные бриобионты Nebela parvula, N. galeata, Heleopera sphagni, Difflugia penardi, Centropyxis constricta . Видовой состав в наиболее низко расположенных экосистемах (осиннике и пихтарнике) весьма гомогенен. Таким образом, гетерогенность видового состава локальных сообществ возрастает с увеличением высоты над уровнем моря.
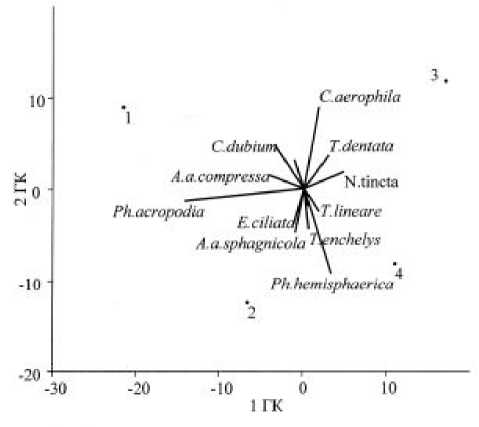
Рис. 4. Результаты ординации видов методом главных компонент на основе относительных обилий доминирующих видов:
1ГК – первая главная компонента (объясняет 55,5% общей дисперсии видовой структуры), 2ГК – вторая главная компонента (26,8%)
Структурно сообщества разделяются на два варианта (рис. 4): из более низко расположенных осинника и пихтарника с преобладанием в доминантном комплексе (около 30% от общей численности) мелкого вида лобозных амеб Phryganella acropodia и формирующихся в более суровых условиях лесотундры и тундры бриофилов Nebela tincta и Tracheleuglypha dentata. Вместе с тем, во многих случаях изменения представленности в сообществах отдельных видов не проявляют направленных тенденций. Так, Phryganella hemisphaerica, Trinema enchelys массово развиваются в пихтарнике и тундре, Centropyxis aerophila – в осиннике и лесотундре, Corythion dubium – в осиннике и тундре.
Таким образом, полученные данные дополняют имеющуюся информацию по составу, структуре и закономерностям изменений сообществ нанофауны, населяющей мхи, вдоль высотного градиента. Несмотря на ярко выраженную высотную поясность, выражающуюся в смене типов биогеоценозов от широколиственных лесов, через темнохвойную тайгу, лесотундру к тундре, изменения сообществ раковинных корненожек не столь направлены. В целом, видовой состав и структура доминирующего комплекса видов весьма однородны и сходы с таковыми в Западной Европе [27, 31]. Специфика Прибайкальского варианта высотного градиента в ризоподном сообществе – возрастание видового богатства, обилия, видовой специфики и пространственной гетерогенности сообществ в поясе мохово-лишайниковой тундры, что может быть связано с более медленными процессами трансформации органики в более суровых условиях, что дает конкурентные преимущества относительно медленно размножающимся раковинным корненожкам [4]. Отсутствие единой тенденции в ценотических изменениях вдоль высотных градиентов характерно не только для населения одноклеточных организмов, но и широко известно для арт-ропод [21]. По всей видимости, на структуру формирующихся сообществ влияет не сам по себе фактор высоты над уровнем моря, а причины, действующие либо в меньшем (увлажненность локального биотопа, наличие пищи, хищников, особенности субстрата), либо в большем (особенности климата, исторические причины) масштабах [21, 26].
ЗАКЛЮЧЕНИЕ
Таким образом, проведенное исследование позволило выявить 49 видов и форм раковинных корненожек, что существенно дополняет известные данные [Балик] о ризоподном населении Прибайкалья. Наиболее характерный элемент фауны – убиквитные виды с широким географическим распространением (Centropyxis aerophila, Phryganella acropodia, Trinema enchelys, Corythion dubium, Assu-lina seminulum), что соответствует известной информации о специфике почвенно-мохового населения раковинных амеб горных территорий [Тодоров, Митчелл, Балик]. Видовое богатство и численность раковинок возрастают в наиболее высоко расположенном (1800 м н.у.м.) поясе моховолишайниковой тундры. Это противоречит данным о убывании [Тодоров] или сохранении на одном уровне [Mit] видового разнообразия с высотой и позволяет предположить, что высота над уровнем моря, как фактор, непосредственно не оказывает влияния на раковинных корненожек, а действует опосредованно через изменения характеристик биогеоценозов как в локальном, так и в биогеографическом масштабах. С высотой возрастает гетеро- генность видового состава локальных сообществ в пределах высотных поясов и изменяется структура доминирующего комплекса видов. При этом многие формы не проявляют направленных тенденций в изменении, как численности, так и относительной представленности в структуре сообщества.
Список литературы Изменения сообществ раковинных амеб, обитающих в эпилитных мхах, вдоль высотного градиента (Прибайкалье, хребет Хамар-Дабан)
- Алексеев Д.В. Раковинные амёбы почв болотных лесов северной подзоны европейской тайги. Автореф. дис. … канд. биол. наук. М.: МПГИ, 1984. 16 с.
- Бобров А.А. Эколого-географические закономерности распространения и структуры сообществ раковинных амёб (Protozoa: Testacea). Дис. … докт. биол. наук. М.: МГУ, 1999. 341 с.
- Бобров А.А. Раковинные амебы и закономерности их распределения в почвах//Почвоведение. 2005. №9. С. 1130-1137.
- Гельцер Ю.Г., Корганова Г.А., Алексеев Д.А. Почвенные раковинные амебы и методы их изучения. М.: Изд-во МГУ, 1985. 79 с.
- Гельцер Ю.Г., Корганова Г.А., Яковлев А.C., Алексеев Д.А. Раковинные корненожки (Testacida) почв//Почвенные простейшие. Л.: Наука, 1980. С. 108-142.
- Карташев А.Г., Смолина Т.В. Влияние нефти на почвенных раковинных амеб (Arcellinida, Euglyphida) в условиях полевого эксперимента//Зоол. журн. 2008. Т. 87. № 9. С. 1027-1033.
- Корганова Г.А. Раковинные амебы (Testacida) некоторых почв Европейской части СССР//Pedobiologia. 1975. Bd. 15. S. 125-431.
- Корганова Г.А. Организация почвенных сообществ раковинных амеб//Зоол. журн. 1999. Т. 78. Вып. 12. С. 1396-1406.
- Корганова Г.А. Адаптивные морфологические структуры и эволюция почвенных раковинных амеб (Protista, Testacea)//Зоол. жур. 2003. Т. 82. Вып. 2. С. 197-214.
- Корганова Г.А. К вопросу о системе простейших и таксономическом положении раковинных амёб (Rhizopoda, Testacea)//Успехи совр. биол. 2004. Т. 124. С. 443-456.
- Кордэ Н.В., Чибисова О.И. Микронаселение почвы основных типов леса Подмосковья -раковинные амёбы (Testacida). В кн.: Лесоводственные исследования в серебряноборском опытном лесничестве. М.: Наука, 1973. С. 167-174.
- Мазей Ю.А., Бубнова О.А. Структура сообщества раковинных амеб (Testacealobosea; Testaceafilosea; Amphitremidae) в напочвенных сфагнумах смешанных лесов Среднего Поволжья//Вестник зоологии. 2008. №1. С. 41-48.
- Мазей Ю.А., Бубнова О.А. Раковинные амебы в сфагновых биотопах заболоченных лесов//Зоол. журн. 2009. Т.88. №4. С. 387-397.
- Мазей Ю.А., Ембулаева Е.А. Структура сообщества почвенных раковинных амеб в Островцовской лесостепи (Среднее Поволжье): эффект лесостепного градиента//Успехи совр. биол. 2008. Т. 128. №5. С. 532-540.
- Мазей Ю.А., Ембулаева Е.А. Изменение сообществ почвообитающих раковинных амеб вдоль лесостепного градиента в Среднем Поволжье//Аридные экосистемы. 2009. Т.15. № 1(37). С. 13-23.
- Мазей Ю.А., Кабанов А.Н. Раковинные амебы в осоково-сфагновом заболоченном лесу на севере Карелии//Известия ПГПУ им. В.Г. Белинского. Сер. Естественные науки. 2008. № 10(14). С. 101-104.
- Мазей Ю.А., Цыганов А.Н. Пресноводные раковинные амёбы. М.: Товарищество научных изданий КМК, 2006. 300 с.
- Рахлеева А.А. Изменение структуры и разнообразия комплексов почвенных тестацей (Testacea, Protozoa) по элементам мезорельефа Южной Мещеры//Известия АН. Сер. Биол. 1998. № 6. С. 749-754.
- Рахлеева А.А. Изменение комплексов тестацей (Testacea, Protozoa) в ряду пойменных почв Окского заповедника (Южная Мещера)//Известия АН. Сер. Биол. 1999. № 4. С. 478-487.
- Рахлеева А.А. Раковинные амёбы (Testacea, Protozoa) таежных почв Западной Сибири (Сургутское Полесье)//Известия АН. Сер. Биол. 2002. № 6. С. 752-762.
- Andrew N.R., Rodgerson L., Dunlop M. Variation in invertebrate-bryophyte community structure at different spatial scales along altitudinal gradients//J. Biogeogr. 2003. Vol. 30. P. 731-746.
- Azovsky A. I. Size-dependent species-area relationships in benthos: is the world more diverse for microbes?//Ecography. 2002. Vol. 25. P. 273-282.
- Balik V. Testacean amoebae fauna (Protozoa, Rhizopoda, Testacea) from the Asian part of the USSR (Regions of the Baikal Lake and Khabarovsk)//Acta Soc. Zool. Bohemoslov. 1992. V. 56. P. 93-107.
- Bonnet L. Le peuplement thecamoebien des mousses corticoles//Protistologica. 1973. T. 9. P. 319-338.
- Chardez D. Ecologie generale des thecamoebiens//Bull. Inst. Agron. Stat. Rech. Gembloux. 1965. T. 33. P. 307-341.
- Fleishman E., Fay J.P., Murphy D.D. Upsides and downsides: contrasting topographic gradients in species richness and associated scenarios for climate change//J. Biogeogr. 2000. Vol. 27. P. 1209-1219.
- Mitchell E.A.D., Bragazza L., Gerdol R. Testate amoebae (Protista) communities in Hylocomium splendens (Hedw.) B.S.G. (Bryophyta): relationships with altitude, and moss elemental chemistry//Protist. 2004. V. 155. P. 423-436.
- Mitchell E.A.D., Meisterfeld R. Taxonomic confusion blurs the debate on cosmopolitan versus local endemism of freeliving protists//Protist. 2005. Vol. 156. P. 263-267.
- Schönborn W. The topophenetic analysis as a method to elucidate the phylogeny of testate amebas (Protozoa, Testacealobosia and Testaceafilosia)//Arch. Protistenk. 1989. Bd. 137. S. 223-245.
- Smith H., Bobrov A., Lara E. Diversity and biogeography of testate amoebae//Biodiv. Cons. 2007. Vol. 17. P. 329-343.
- Todorov M. Observation on the soil and moss testate amoebae (Protozoa: Rhizopoda) from Pirin mountain (Bulgaria)//Acta Zool. Bulg. 1998. V. 50. P. 19-29.
- Wilkinson D. Have we underestimated the importance of humans in the biogeography of free-living terrestrial microorganisms?//J. Biogeogr. 2010. Vol. 37. P. 393-397.
