Изменения состояния нейроцитов и нейрон-глиальных соотношений в спинномозговых узлах собак после удлинения конечности

Автор: Сафонова Г.Д., Коваленко А.П., Ерофеев С.А., Попков Д.А., Еманов А.А.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 2, 2005 года.
Бесплатный доступ
В постдистракционном периоде при удлинении голени собак ≈ на 15 % в нейроцитах спинномозговых узлов обнаружены обратимые изменения структуры части клеток в виде гиперхромии нейроплазмы и ядра, периферического хроматолиза, дислокации ядрышка и ядра; увеличение количества пери- и межнейрональных глиоцитов. Изменения наиболее выраженны в узлах на стороне удлинения при темпе дистракции 3 мм в сутки, минимальны - контралатерально при удлинении с темпом 1 мм в сутки.
Удлинение голени, спинномозговой узел, нейрон, глиоцит
Короткий адрес: https://sciup.org/142120793
IDR: 142120793
Changes in leucocytes conditions and neuron-glia relations of canine spinal ganglia after limb lengthening
In the post-distraction period when a canine leg was subjected to about 15% lengthening reversible changes in the neurocytes of the spinal ganglia were noted in the structure of a part of cells as neuroplasm and nucleus hyperchromia, peripheral chromatolysis, nucleolus and nucleus dislocations; increase of peri- and interneuronal gliacytes in number. The changes were the most marked in the spinal ganglia of the lengthening side for the distraction rate of 3 mm per day and they were minimal - contralaterally for the lengthening rate of 1 mm per day.
Текст научной статьи Изменения состояния нейроцитов и нейрон-глиальных соотношений в спинномозговых узлах собак после удлинения конечности
В настоящее время дистракционный остеосинтез широко используется в клинической практике. Влиянию напряжения растяжения на окружающие кость ткани, в т.ч. периферические нервы, посвящено большое количество работ. Состояние же нейроцитов спинномозговых узлов, воспринимающих информацию от болевых и мышечносухожильных рецепторов удлиняемого сегмента конечности, представлено в единичных работах [8]. Поскольку большинство нейроцитов спинномозговых узлов является чувствительными клетками, т.е. частью афферентного звена рефлекторной дуги, они в первую очередь реагируют на происходящие на периферии изменения, а преобразования в теле нервной клетки, ее ядре и нейроплазме отражают состояние иннервируемых мышц и могут служить индикатором силы воздействующего фактора, его предельной величины. Известно, что при повышении функциональной активности нейроцитов увеличивается количество сателлитной глии, принимающей активное участие в его специфической деятельности [6, 7]. В работах многих авторов также показано увеличение количества перинейрональных глиоцитов в центральной и периферической нервной системе при различных воздействиях в сочетании с изменением либо сохранением структуры нейронов [2, 5, 6, 7, 10 и др.]. Однако количественная характеристика нейрон-глиальных взаимоотношений в условиях дистракции в доступной нам литературе не представлена.
Целью настоящего исследования было изучение влияния различных режимов дистракции на морфофункциональное состояние нейроци-тов спинномозговых узлов и количественная оценка изменения нейрон-глиальных соотношений в данных условиях.
МЕТОДИКА ИССЛЕДОВАНИЙ
Эксперимент выполнен на 6 взрослых беспородных собаках, которым производили удлинение правой голени на 28-30 мм (≈15%) раз- личными способами дистракционного остеосинтеза: в автоматическом режиме с темпом дистракции 3 мм в сутки за 180 приемов (n = 3, пе- риод дистракции – 10 суток, фиксации в аппарате 30 суток); в ручном режиме с использованием интрамедуллярного введения противо-изогнутых спиц с темпом 1 мм в сутки за 4 приема (n = 3, период дистракции – 28 суток, фиксации – 14-15 суток).
Животных выводили из опыта через 45-48 суток. Их содержание, кормление и эвтаназию осуществляли в соответствии с требованиями инструкции № 12/313 Министерства здравоохранения Российской Федерации «Санитарные правила по устройству, оборудованию и содержанию экспериментальных биологических клиник» от 06.01.73. После эвтаназии животного (посредством введения предельной дозы барбитуратов) вычленяли блок, содержащий поясничный и крестцовый отделы позвоночника, удаляли дужки позвонков и извлекали спинной мозг вместе с соответствующими каждому сегменту корешками и ганглиями (рационализаторское предложение авторов № 55/02). Для нейрогистологических исследований использованы спинномозговые ганглии (L 6 , L 7 , S 1 , n=36), нейроциты которых получают информацию непосредственно от мышц и сухожилий голени, подвергающихся растяжению. Материал фиксировали в смеси Бродского, обезвоживали в спиртах возрастающей концентрации и заливали в парафин. Продольные серийные срезы ганглиев толщиной 5 мкм окрашивали по методу Ниссля, галлоцианином-хромовыми квасцами по Эйнарсону, амидочерным 10Б-кислым фуксином.
Для количественных исследований использованы окрашенные галлоцианином-хромовыми квасцами по Эйнарсону срезы ганглиев L7, в которых производили подсчет ядер глиоцитов, формирующих капсулы крупных (n≥75) и мелких (n≥75) нейронов – перинейрональная глия. Также выполнен подсчет ядер глиоцитов, расположенных в межнейрональных пространствах и не включенных в капсулу нейрона. С этой целью определяли количество ядер на единицу площади с использованием сетки, разработанной авторами (рационализаторское предложение № 70/2001), площадь квадрата которой при увеличении 400 составляет 241,8 мкм2. В каждом ганглии количественному анализу подвергнуто не менее 50 полей зрения. Выбор исследуемых групп нейронов был обусловлен тем, что, по данным литературы, крупные нервные клетки ответственны за проприорецепцию, мелкие связаны с болевой чувствительностью [1, 3]. Как известно, капсулу нейроцитов в спинномозговых узлах формируют клетки олигодендроглии [1], характерным признаком которых на гистопрепаратах является округлая форма ядер, в отличие от ядер других сопутствующих клеток (фибробластов, эндотелиоцитов, шванновских клеток), имеющих на срезе другие профили (вытянутый, палочковидный и пр.) [9].
Статистическую обработку материала осуществляли в программе Microsoft Excel, достоверность различий оценивали с использованием t-критерия Стьюдента [4].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
Установлено, что удлинение тазовой конечности на 28-30 мм приводит к появлению в обозначенных спинномозговых ганглиях нейроци-тов с обратимыми изменениями ядра и перикариона и увеличению количества пери- и межнейрональных глиоцитов в сроки эксперимента 45-48 суток. Встречались следующие изменения структуры перикариона: периферической хроматолиз, как локальный, так и равномерно выраженный (рис. 1, а, б), умеренная гиперхромия нейроплазмы, иногда в сочетании с изменением формы тела и наличием большого количества сателлитов (рис. 1, а, г). Характерны были дислокация ядра и ядрышка вплоть до эктопии, их сочетанное перемещение (рис. 1, в, г). Иногда эктопия ядра сопровождалась центральным положением ядрышка, в единичных случаях наблюдалось отложение базофильного вещества на ядерной мембране. Подобные изменения структуры нейроцитов были выражены более значительно в ганглиях на стороне операции при дистракции с темпом 3 мм в сутки, несколько менее – при удлинении с темпом 1 мм в сутки. В контралатеральных узлах отклонения от нормы в структуре нейронов более умеренны, особенно при использовании интрамедуллярного введения спиц и более щадящего темпа дистракции. В основном они были представлены дислокацией ядрышка и краевым периферическим хроматолизом (рис. 1, а).
Ранее Г.Д. Сафонова и В.И. Калякина (1988), исследуя влияние различных режимов дистракции на состояние нервных клеток чувствительных узлов, отмечали, что при удлинении голени до 3 см в различных сериях эксперимента наблюдались умеренные изменения структуры нейроцитов. Авторами выявлено, что в левых (контралатеральных) спинномозговых узлах, наряду с преобладанием нормохромных нейро-цитов, встречались гипохромные клетки, как с центральным положением ядра, так и его дислокацией. В правых ганглиях, наряду с нормохромией большинства нейроцитов, определялись более выраженные обратимые изменения структуры нервных клеток с наличием в них периферического хроматолиза, эктопии ядра и ядрышка, прослеживалось усиление глиальной реакции. При удлинении голени свыше 3 см (до 4-5 см) в серии, где использовался более щадящий режим, изменения нейроцитов также преимуще- ственно носили обратимый характер. В серии с более жестким режимом в спинномозговых узлах значительная часть нейроцитов имела признаки перераздражения, вплоть до появления в небольшой части клеток необратимых структурных изменений, была выраженной глиальная реакция [8].
Физиологическое значение увеличения количества сателлитов состоит в метаболическом обеспечении усиленно функционирующих нервных клеток [7]. Показано, что при кратковременном воздействии не происходит деления глиальных элементов, однако при продолжительном действии раздражающего фактора изменение нейрон-глиальных соотношений осуществляется не только за счет миграции глиоцитов из межнейрональных пространств к нейронам, но и за счет их деления [7]. Поскольку в нашем исследовании выявлено увеличение не только пери-, но и межнейрональной глии, можно предположить, что в данном случае наблюдается деление глиальных клеток с их миграцией к нейроцитам. Увеличение количества межнейрональных глиоцитов прослеживалось в виде образования небольших групп глиоцитов (рис. 1, а), а также обширных скоплений глиальных элементов (рис. 1, г). Формирование капсулы в изученных сериях существенно не различалось и можно было выделить 4 основных варианта увеличения количества сателлитов в ганглиях на стороне удлинения конечности по сравнению с контралатеральной: 1) равномерное увеличение количества глиальных клеток, включенных в однослойную капсулу нейроцита (рис. 1, а); 2) скопление глиальных элементов в области отхождения псевдоуниполярного отростка (рис. 1, б); 3) скопление ядер в области локального периферического хроматолиза (рис. 1, б); 4) образование второго и более слоев капсулы (рис. 1, в, г).
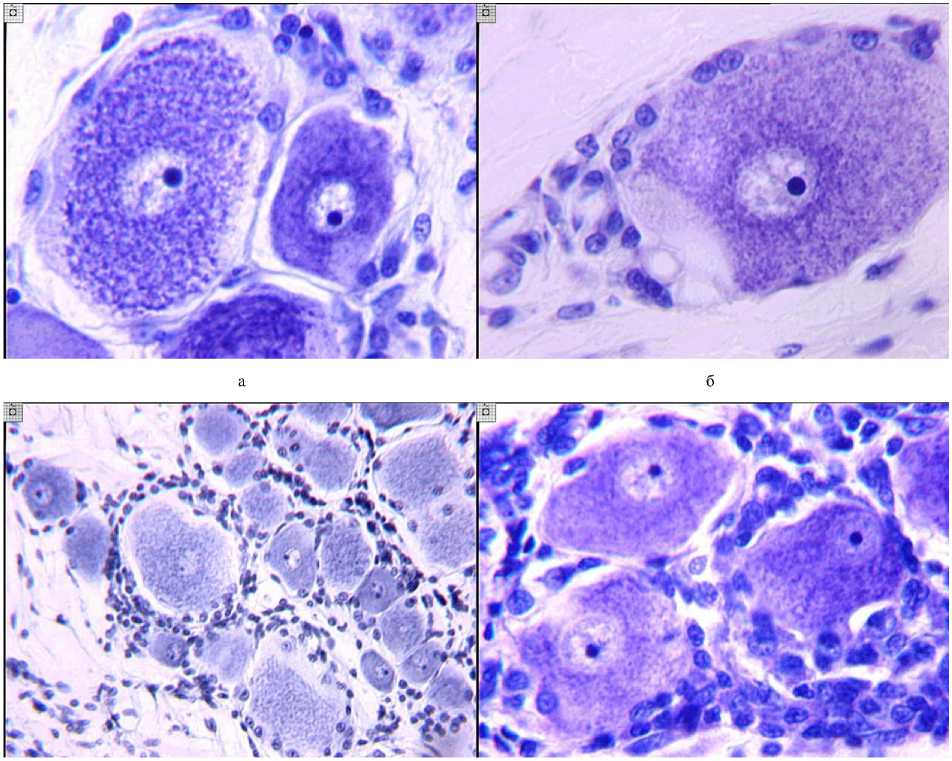
в г
Рис. 1. Сочетание структурных изменений нейронов и увеличения количества глиоцитов: а - равномерно выраженный периферический хроматолиз и дислокация ядрышек при умеренном увеличении количества глиоцитов в капсуле и межнейрональных пространствах; б - сочетание локального периферического хроматолиза и скопления глиоцитов в области отхождения отростка; в - формирование многослойной капсулы крупного нейрона, эктопия ядрышек средних и мелких нейро-цитов; г - сочетание выраженной глиальной реакции с изменением формы отдельных нейронов и эктопией ядра. а, б, г -окраска по Нисслю, ув. 400; в - окраска по Эйнарсону, ув. 160
Наиболее распространенным являлся первый вариант усиления глиальной реакции, особенно при удлинении голени с темпом 1 мм в сутки, где структурные изменения нейроцитов встречались реже. Формирование многослойной капсулы более характерно для автодистракции. Второй и третий варианты распределения глиальных элементов часто встречались в сочетании. Увеличение количества сателлитов наблюдалось как вокруг нейронов с измененной, так и сохранной структурой. Учитывая, что в большинстве нервных клеток исследованных ганглиев не прослеживалось необратимых структурных преобразований, изменения нейрон-глиальных взаимоотношений, вероятно, были обусловлены усиленным функционированием нервных клеток, которые, участвуя в иннервации конечности, подобным образом реагировали на перестройки, происходящие в удлиняемом сегменте и после окончания периодов дистракции и фиксации.
Отмечена зависимость количественного проявления глиальной реакции от способа и темпа удлинения. Через 1,5 месяца от начала эксперимента максимальные средние значения количества ядер капсульных глиоцитов наблюдались при автоматическом удлинении голени с темпом 3 мм в сутки. Капсула крупных нейронов в контралатеральном узле состояла из 12,69 0,24 сателлитов, мелких – 4,343 0,1, на стороне удлинения конечности – 15,18 0,27 и 5,286 0,13 соответственно (p<0,001). При интрамедуллярном введении спиц с темпом дистракции 1 мм в сутки исследуемые показатели на стороне операции в данный период существенно не отличались от таковых при удлинении с темпом 3 мм в сутки и составили 14,43 0,34 сателлитов крупных и 5,0 0,15 сателлитов – мелких нейронов. Однако эти же показатели в контралатеральном узле были достоверно (p<0,001) меньше: 10,59 0,23 и 3,88 0,09 соответственно (табл.). Увеличенное количество ядер капсульных глиоцитов в контралатеральных ганглиях при темпе дистракции 3 мм в сутки по сравнению с темпом 1 мм может быть обусловлено двумя факторами: во-первых, более интенсивным опосредованным влиянием дистракции с высоким темпом удлинения, во-вторых, увеличением нагрузки на левую тазовую конечность в связи с меньшим использованием правой конечности.
Плотность расположения ядер глиальных элементов в межнейрональных пространствах в изученный срок в зависимости от эксперимента не различалась. Однако сохранялась тенденция к увеличению количества глиоцитов на квадрат сетки площадью 241,8 мкм2 в ганглиях на стороне операции, по сравнению с контралатеральной (p<0,001). Средние значения этого показателя составили 4,261 0,081 на стороне удлинения и 3,377 0,084 контралатерально при дистракции с темпом 3 мм в сутки, 4,166 0,057 и 3,26 0,049 соответственно при дистракции с темпом 1 мм в сутки (табл.). Данный факт свидетельствует о том, что в обозначенный период эксперимента вследствие уменьшения напряжения в системе «нейрон-глия» снижена или отсутствует необходимость пролиферации глиальных элементов и их перемещения к усиленно функционирующим нейронам как при интенсивном воздействии – удлинении с темпом 3 мм в сутки по истечении достаточно длительного периода (1 месяц после окончания дистракции), так и после менее интенсивного воздействия удлинения голени с темпом 1 мм в сутки при меньшем периоде покоя (2 недели). Это можно объяснить стиханием перестроечных процессов в стадии адаптации реконструктивного нейромиогенеза мягкотканных структур удлиняемого сегмента тазовой конечности [11].
Таким образом, при увеличении длины голени на 28-30 мм, т.е. на 15 % от исходной длины, после окончания периодов дистракции и фиксации в состоянии нейроцитов чувствительных узлов прослеживались изменения преимущественно обратимого характера в сочетании с глиальной реакцией, выраженной в различной степени. Несмотря на то, что постдистракционный период был более продолжительным (более чем в 2 раза) при удлинении с темпом 3 мм в сутки, в исследованных спинномозговых узлах этих животных прослеживались изменения более выраженные, чем при удлинении с темпом 1 мм в сутки.
Таблица
Количественная характеристика нейрон-глиальных соотношений в спинномозговых узлах беспородных собак при удлинении голени различными способами (М m)
|
Серия эксперимента |
Количество глиоцитов, образующих капсулу крупных нейронов |
Количество глиоцитов, образующих капсулу мелких нейронов |
Количество глиоцитов, в межнейрональном пространстве (S=241,8мкм2) |
|||
|
контралатерально |
ипсилатерально |
контралатерально |
ипсилатерально |
контралатерально |
ипсилатерально |
|
|
Автодистракция с темпом 3 мм в сутки за 180 приемов |
12,69 0,24 |
15,179 0,267* |
4,349 0,095 |
5,286 0,13* |
3,377 0,084 |
4,261 0,081* |
|
Дистракция с темпом 1 мм в сутки за 4 приема и интрамедуллярным введением спиц |
10,59 0,23 |
14,43 0,34* |
3,88 0,089 |
5,0 0,147* |
3,26 0,049 |
4,166 0,057* |
