Изменения в миелоидном звене системы крови у стрессированных белых крыс с экспериментальным гипотиреозом

Автор: Гармаева Дэнсэма Владимировна, Васильева Людмила Сергеевна
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Физиология и экология человека и животных
Статья в выпуске: 4-1, 2014 года.
Бесплатный доступ
Выявлены нарушения реакции миелоидного звена системы крови на иммобилизационный стресс в условиях экспериментального гипотиреоза у белых крыс. Установлено, что при гипотиреозе стресс в стадии тревоги не вызывает лейкоцитоз, характерный для животных с эутиреозом, а в стадии резистентности провоцирует лейкопению, обусловленную эозинопенией и нейтропенией с истощением костномозгового резерва этих грану-лоцитов, который восстанавливается лишь через месяц.
Гипотиреоз, иммобилизационный стресс, эозинофилы, нейтрофилы, базофилы, лейкоциты, миелопоэз
Короткий адрес: https://sciup.org/148182428
IDR: 148182428 | УДК: 612.419:612.119:616.441-008.64
Changes in a myeloid part of blood system at stressed white rats with experimental hypothyroidism
It is revealed that at white rats at experimental hypothyroidism a myeloid part of blood system has nonadequate reaction on immobilization stress'. It is established that at hypothyroidism the stress does not cause the leucocytosis at an alarm stage as for animals with euthyroidism, but at a resistance stage it induces leucopenia caused by eosinoopenia and neutropenia with an exhaustion of a marrowy reserve of these granulocytes, which restores only in a month.
Текст научной статьи Изменения в миелоидном звене системы крови у стрессированных белых крыс с экспериментальным гипотиреозом
Актуальность исследования обусловлена важнейшей ролью гранулоцитов крови в защитных и адаптационных реакциях организма, которые могут существенно изменяться в условиях дефицита энергии, создаваемого гипотиреозом. На сегодняшний день сведений о состоянии миелоидного звена в условиях гипотиреоза в сочетании со стрессом в литературе крайне мало [5]. Становится очевидной необходимость исследований в этой области, что даст возможность расширить базу знаний об адаптивных возможностях организма к стрессорным факторам при гипотиреозе и разработать адекватные пути коррекции.
Цель исследования – выявление нарушений в миелоидном звене системы крови в условиях экспериментального гипотиреоза после стрессорного воздействия.
Материалы и методы. Экспериментальные исследования проведены на 48 беспородных белых крысах-самцах массой 180–200 г в осеннезимний период. Содержание, питание, уход соответствовали ГОСТ Р 5025892. Исследования проводились согласно правилам лабораторной практики при проведении доклинических исследований в РФ (ГОСТ З 51000.3-96 и 51000.4-96). Гипотиреоз моделировали введением перорально (с кормом) мерказолила в дозе 10 мг/кг ежедневно в течение 8 недель [3, 7]. Иммобилиза-ционный стресс моделировали однократной 6часовой иммобилизацией на спине [6]. Крысы были разделены на 3 группы. Первая подопытная группа (ГS) включала 24 крыс, которым моделировали гипотиреоз, а затем (сразу после отмены мерказолила) – иммобилизационный стресс. Вторая группа (S) включала 16 крыс, которые подвергались только иммобилизационно- му стрессу. Восемь крыс оставались интактными и составили контрольную группу. Крыс выводили из эксперимента с помощью эфирной эвтаназии. Материал для исследования брали на 2-е сут. (стадия тревоги стресса [4]), 7-е сут. (стадия резистентности стресса) и 28-е сут. (отдаленные результаты) после отмены мерказоли-ла. Кровь для исследования брали из хвостовой вены, затем после эвтаназии извлекали бедренную кость для изучения красного костного мозга (ККМ). В периферической крови определяли количество лейкоцитов в 1л. Мазки крови и ККМ окрашивали по Паппенгейму [2]. В мазках крови подсчитывали лейкоцитарную формулу (с последующим пересчетом %-ного количества лейкоцитов в абсолютное), в мазках костного мозга – миелограмму (на 1000 клеток). Высчитывали индексы пролиферации (ИП) и созревания (ИС) клеток миелопоэза по формулам [1]: ИП=[(МЦ*0+ММЦ*1)/(МЦ+ММЦ)]*∑;
ИС=[(ММЦ*0+ПЯ*1+СЯ*2)/(ММЦ+ПЯ+СЯ)]* ∑, где МЦ – количество миелоцитов, ММЦ – количество метамиелоцитов, ПЯ – количество палочкоядерных гранулоцитов, СЯ – количество сегментоядерных гранулоцитов, ∑ – сумма всех клеток нейтрофильного, эозинофильного или базофильного ростка.
Полученные данные обрабатывали статистически с определением типа распределения вариационных рядов, среднего арифметического, ошибки среднего, среднего квадратичного отклонения [Statistica v.6]. Достоверность различий средних величин определяли по t-критерию Стьюдента при р<0,05.
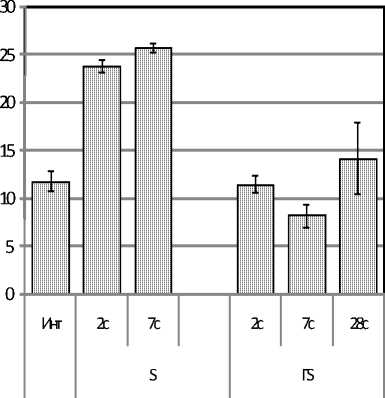
Результаты исследования. У крыс с эутиреоидным статусом к концу стадии тревоги стресса (на 2 сутки) развивался лейкоцитоз, который сохранялся до 7 суток. У крыс с гипотиреозом (который характеризовался сохранением количества лейкоцитов в диапазоне нормы) стресс на 2-е сутки не вызывал лейкоцитоз, а к 7-е суткам наблюдения (в стадию резистетно-сти) провоцировал даже лейкопению, устраняемую только к 28-е суткам (рис. 1А).
А Б
Рис. 1. Изменение количества лейкоцитов (*109/л) в периферической крови (рис. А)
и клеток базофилопоэза из 1000 клеток (рис. Б) у стрессированных крыс с эутиреозом (S) и с гипотиреозом (ГS)
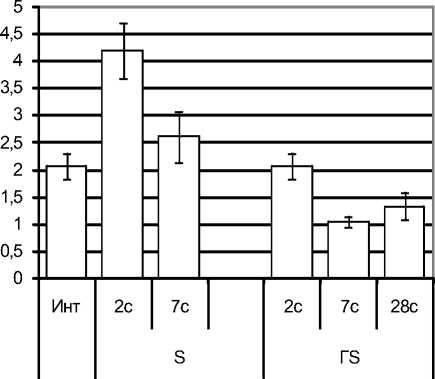
Численность базофилов в периферической крови не изменялась у обеих групп крыс на протяжении всего наблюдения (возможно, в связи с очень малой численностью этих клеток в крови). Вместе с тем в ККМ базофильный росток заметно изменялся (рис. 1-Б). У животных с эутирео-зом при стрессе на протяжении всего наблюдения количество клеток базофильного ростка в ККМ проявляло стойкую тенденцию к уменьшению. У крыс с гипотиреозом, наоборот, базофильный росток в стадии тревоги стресса сохранялся в диапазоне нормы, в стадии резистентно- сти увеличился в 4,7 раза (р<0,05), затем его численность постепенно снижалась, но даже к 28-м суткам превышала норму в 2,3 раза (р<0,05).
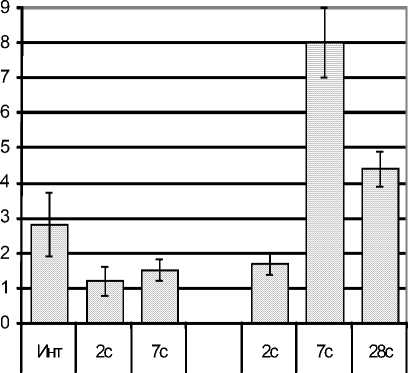
Количество эозинофилов в периферической крови у крыс с эутиреозом на 2-е сутки после стрессорного воздействия уменьшилось в 3 раза по отношению к норме (р<0,05), т.е. развивалась эозинопения (что является классическим признаком стадии тревоги стресса [4]), в стадии резистентности количество эозинофилов увеличивалось, приближаясь к норме (рис. 2).
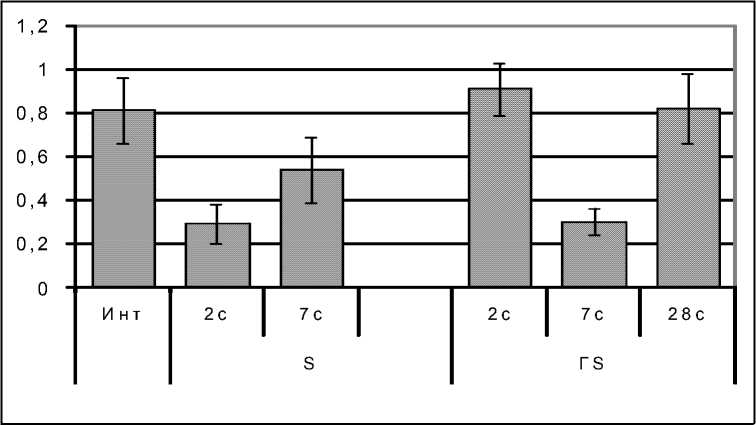
Рис. 2. Динамика абсолютного количества эозинофилов в периферической крови у стрессированных крыс с эутиреозом (S) и с гипотиреозом (ГS)
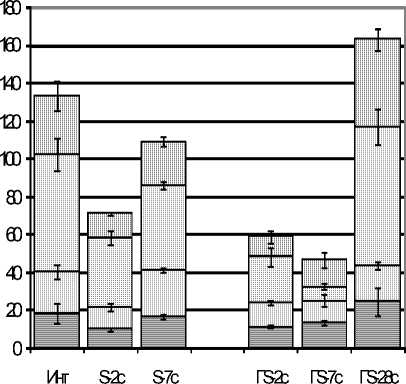
В ответ на эозинопению в стадии тревоги у крыс с эутиреозом компенсаторно стимулируется пролиферация клеток эозинофильного ряда. При этом индекс пролиферации увеличивается почти вдвое (рис. 3 Б), а количество МЦ в 1,5 раза, ММЦ в 2,4 раза, по сравнению с интактными крысами (рис. 3 А). Вместе с тем индекс созревания снижен по сравнению с нормой в 2,7 раза, что ведет к опустошению костномозгового депо зрелых эозинофилов, которых становится в 3,3 раза меньше нормы (р<0,05, рис. 3 А, Б). В стадии резистентности созревание эозинофилов ускоряется вдвое, появляется резерв зрелых клеток, развивается тенденция к нормализации эо-зинофилопоэза.
При гипотиреозе под влиянием стресса эо-зинопения формировалась позже, через 7 суток после стрессорного воздействия, когда их количество уменьшилось в 2,7 раза по сравнению с интактными крысами (р<0,05), к 28-м суткам их количество в крови нормализовалось (рис. 3 А).
Перестройка эозинофилопоэза в ККМ под влиянием стресса у крыс с гипотиреозом тоже запаздывала, как и эозинопения, и отличалась торможением (а не активацией) пролиферации клеток. К 7-м суткам количество МЦ уменьшилось в 2 раза, ММЦ – в 1,5 раза, индекс пролиферации – в 1,6 раза (р<0,05, рис. 3 А, Б) по сравнению с нормальным значением. Созревание клеток тоже замедлялось, и резерв зрелых эозинофилов проявлял тенденцию к уменьшению (рис. 3 А). Лишь через 28 суток показатели эозинофилопоэза приближались к норме.
Таким образом, при эутиреозе в стадии тревоги стресса эозинопения сопровождалась кратковременным истощением резерва эозинофилов в костном мозге и компенсаторной стимуляцией эозинфилопоэза. В условиях гипотиреоза стресс, несмотря на эозинопению, не только не стимулировал эозинофилопоэз, но и существенно подавлял его активность, что тем не менее не приводило к опустошению костномозгового резерва зрелых эозинофилов, а лишь намечало тенденцию к его уменьшению. Эти данные дают основание считать, что при гипотиреозе эозинопения возникает в результате торможения миграции эозинофилов из костного мозга в кровь из-за дефицита энергии, создаваемого гипотиреоидным состоянием.
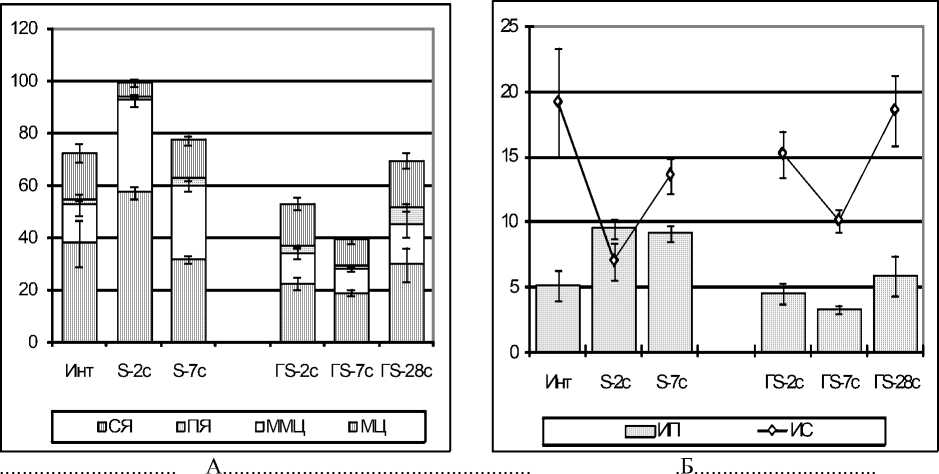
Рис. 3. Количество клеток эозинофилопоэза из 1000 клеток (рис. А), индекс пролиферации (ИП, усл. ед.) и созревания (ИС, усл. ед., рис. Б) у животных с эутиреозом (S) и стрессированных крыс с гипотиреозом (ГS)
Обозначения: МЦ – миелоциты, ММЦ – метамиелоциты, ПЯ – палочкоядерные эозинофилы, СЯ – сегментоядерные эозинофилы
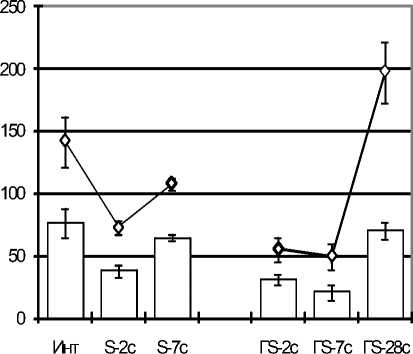
Количество нейтрофилов у крыс с эутирео-зом в стадии тревоги стресса увеличивалось преимущественно за счет увеличения в 2 раза количества СЯ–нейтрофилов (р<0,05; рис. 4 А), которое в стадии резистентности нормализовалось. У крыс с гипотиреозом, имеющих изна- чально сниженное количество СЯ-нейтрофилов, стрессорное воздействие увеличило на 2-е сутки наблюдения численность этих клеток в 2,5 раза (р<0,05), что привело к нормализации этого показателя.
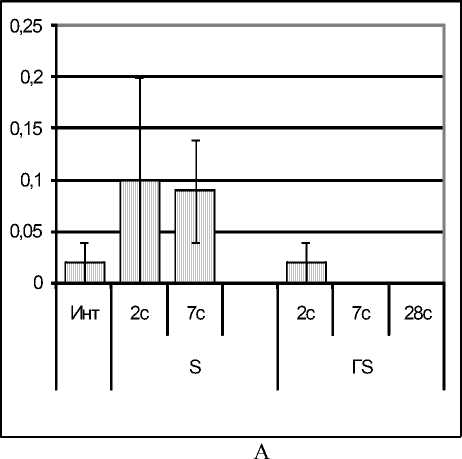
Рис. 4. Динамика абсолютного количества ПЯ-нейтрофилов (рис. А) и СЯ-нейтрофилов (рис. Б) в периферической крови у крыс с эутериоидным статусом (S) и стрессированных крыс с гипотиреозом (ГS) из 100 клеток
Однако с 7-х суток наблюдения количество нейтрофилов в крови этих крыс резко уменьшалось, при этом ПЯ-нейтрофилы совсем не обнаруживались, а численность СЯ-нейтрофилов уменьшилась в 2 раза (р<0,05, рис. 4 Б), по сравнению с интактными крысами. На основании этих данных можно говорить о формировании под действием стресса в условиях гипотиреоза устойчивой нейтропении с отсутствием ПЯ-нейтрофилов.
В нейтрофильном ростке ККМ после стрессорного воздействия у крыс и с эутирео-зом, и с гипотиреозом на 2-е сутки наблюде- ния в ККМ происходило уменьшение количества всех клеток нейтрофильного ряда. При эутиреозе количество МЦ и ММЦ было в 1,9 раза меньше нормы, при гипотиреозе – в 1,6 раза (рис. 5 А).
в мд D мщ □ гя D ся
А
□ип -о-ис
Б
Рис. 5. Количество клеток нейтрофилопоэза из 1000 клеток (рис. А), индексы пролиферации и созревания (усл. ед., рис. Б) у крыс с эутиреозом (S) и стрессированных крыс с гипотиреозом (ГS). Обозначения: МЦ – миелоциты, ММЦ – метамиелоциты, ПЯ – палочкоядерные нейтрофилы, СЯ – сегментоядерные нейтрофилы, ИП – индекс пролиферации, ИС – индекс созревания
Резерв ПЯ- и СЯ-нейтрофилов при эути-реозе уменьшился в 1,7 и 2,4 раза соответственно, а при гипотиреозе – в 2,5 и 3 раза по сравнению с интактными крысами (р<0,05, рис. 5 А). При этом скорость деления бластных форм (ИП) и созревание СЯ-нейтрофилов (ИС) у крыс с эутиреозом снизились в 2 раза, а у крыс с гипотиреозом – в 2,5 раза по сравнению с нормальным значением (р<0,05, рис. 5 Б). Таким образом, через 2 суток после стрес-сорного воздействия, независимо от тиреоидного статуса, в ККМ происходит истощение резерва зрелых нейтрофилов в результате торможения нейтрофилопоэза в сочетании с усиленным выбросом нейтрофилов в кровь.
Через 7 суток после стрессорного воздействия при эутиреозе в ККМ количество клеток нейтрофильного ряда и их пролиферация нормализовались (рис. 5 А), но скорость созревания (ИС) осталась по-прежнему низкой по сравнению с нормой (рис. 5 Б). У стрессированных крыс с гипотиреозом на 7-е сутки наблюдения пролиферация и созревание нейтрофилов еще больше замедлялись и количество клеток нейтрофильного ряда было по-прежнему низким, но через 28 суток экспери- мента пролиферация бластных форм нормализовалась, а созревание ускорилось и в 1,4 раза и превышало норму (р<0,05, рис. 5 Б), что привело к нормализации количества клеток нейтрофильного ряда (рис. 5 А). Таким образом, при эутиреозе численность нейтрофильного ростка нормализовалась уже на 7-е сутки (в стадию резистентности стресса), а при гипотиреозе – лишь на 28-е сутки наблюдения.
Выводы
-
1. Иммобилизационный стресс вызывает при эутиреозе лейкоцитоз в стадии тревоги (на 2-е сутки), а у крыс с гипотиреозом – лейкопению в стадию резистентности (на 7-е сутки).
-
2. Количество базофилов в периферической крови не зависит от тиреоидного статуса и не изменяется под влиянием стресса, но численность клеток базофильного ростка ККМ под влиянием стресса у крыс с эутиреозом проявляет стойкую тенденцию к уменьшению, а у крыс с гипотиреозом увеличивается в 4,7 раза (р<0,05) в стадию резистентности.
-
3. Под влиянием стресса независимо от тиреоидного статуса развивается эозинопения, но при эутиреозе – на 2-е сутки наблюдения, а при
-
4. Независимо от тиреоидного статуса стрессорное воздействие приводит к увеличению в стадии тревоги (2-е сут.) количества нейтрофилов в крови, формируя при эутиреозе лейкоцитоз и нейтрофилию со сдвигом лейкоцитарной формулы влево, а при гипотиреозе – компенсируя исходную нейтропению за счет нормализации численности нейтрофилов. Этот эффект стресса обусловлен усиленным выбросом нейтрофилов из костного мозга в кровь и сопровождается торможением нейтрофилопоэза с истощением резерва зрелых нейтрофилов.
-
5. В стадию резистентности стресса (7-е сут.) при эутиреозе количество нейтрофилов в крови нормализуется за счет активации ней-трофилопоэза, а при гипотиреозе развивается устойчивая нейтропения со сдвигом лейкоцитарной формулы вправо на фоне углубляющегося торможения нейтрофилопоэза, который активизируется лишь к 28-м суткам наблюдения.
гипотиреозе – на 7-е сутки. Эозинопения при эутиреозе временно истощает резерв зрелых эозинофилов в костном мозге и компенсаторно стимулирует эозинфилопоэз, а при гипотиреозе она обусловлена торможением миграции эозинофилов из костного мозга в кровь и сопровождается лишь тенденцией к уменьшению костномозгового резерва этих клеток на фоне подавления активности эозинофилопоэза.
Заключение. Хорошо известно, что низкий уровень тиреоидных гормонов, свойственный гипотиреоидному состоянию, приводит к нарушению основного обмена и дефициту энергии [5, 7], которая необходима для реализации всех защитных реакций организма, в том числе адаптационной. При стресс-реакции происходит мобилизация энергетических ресурсов за счет активации гипоталамо-гипо-физо-адреналовой и адренергической систем, которая в стадии тревоги стресса приводит к повышению в крови уровня катехоламинов и глюкокортикоидов [4], влияющих на метаболизм. По-видимому, эти гормоны в условиях гипотиреоза могут частично компенсировать недостаток регуляторных влияний на метаболизм со стороны тиреоидных гормонов и временно (на период стадии тревоги стресса)
уменьшить дефицит энергии в клетках. По данным проведенного исследования, это проявляется временной нормализацией гранулоцитарного состава крови. Вероятно, этот эффект стресса в условиях гипотиреоидного состояния следует считать положительным. Вместе с тем, как убедительно показали результаты исследования, в условиях гипотиреоза «ценой» этого кратковременного улучшения является резкое ослабление гранулоцитарных ростков с истощением резерва нейтрофилов в стадии резистентности стресса, развитием лейкопении со сдвигом лейкоцитарной формулы вправо, что, безусловно, является негативным для организма. В совокупности полученные данные дают основание считать, что в условиях дефицита энергии, создаваемого гипотиреозом, гранулоциты не способны адекватно реагировать на стрессорное воздействие и осуществлять свои защитные функции.
