Изменения внутриорганного сосудистого русла тимуса в сравнительном орфологическом ряду неполовозрелых позвоночных

Автор: Юрчинский Владислав Янович, Ерофеева Людмила Михайловна
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 1 т.24, 2016 года.
Бесплатный доступ
В статье представлены результаты анализа морфологических изменений различных звеньев сосудистого русла тимуса в сравнительном ряду позвоночных животных и человека. С использованием методов световой микроскопии авторами исследованы гистологические препараты тимуса 15 видов неполовозрелых позвоночных, относящихся к четырем классам. В результате чего обнаружены отличия в характере кровоснабжения коркового и мозгового вещества тимуса позвоночных. Выявлена зависимость количества и площади сосудов микроциркуляторного русла (МЦР) тимуса от уровня организации, особенностей биологии вида, а также систематической принадлежности. Полученные результаты представляют теоретический и практический интерес для биологии морфологии, физиологии и иммунологии.
Тимус, кровеносное русло, позвоночные животные, сравнительная морфология
Короткий адрес: https://sciup.org/143177102
IDR: 143177102
Changes of intra-organ's vascular bed of the thymus in the comparative morphological rows of immature mammalian animals
With the use of methods of light microscopy we produce comparison morphological investigation of the intra organ vascular bed of a thymus in animals and human. The article represents of analysis of morphological imaging different parts of the vascular bed in the comparative morphological a number of four classes of vertebral animals (Chordata, Tetrapoda): Amphibia, Reptilia, Aves, Mammalia. Therefore differences in nature of blood supply of cortical and medullar substance of a thymus of vertebrata are found. It has revealed the similarities and the differences in the structure of vascular bed of thymus gland of vertebral animals which are remarkable for the organization level, biological features and for their adaptation to the habitat.
Текст научной статьи Изменения внутриорганного сосудистого русла тимуса в сравнительном орфологическом ряду неполовозрелых позвоночных
Введение. Всестороннее изучение сосудистого русла является одним из приоритетных направлений биологии и медицины [3, 9, 15, 17]. Большой теоретический и практический интерес с точки зрения морфологии, физиологии и иммунологии представляет собой изучение кровеносного русла тимуса. Исследования различных групп сосудов тимуса тесно связаны с проблемой эндокринной регуляции иммуногенеза [7, 12], продиктованы необходимостью выяснения механизмов циркуляции лимфоцитов [17, 18], а так же способствуют более глубокому пониманию морфологических основ гематотимусного барьера [2, 16]. Во многом благодаря микроциркуляторному руслу обеспечиваются стабильность тканевого метаболизма и механизмы поддержания гомеостаза [9, 15, 1, 4]. По этим причинам детальное изучение морфологии различных групп кровеносных сосудов тимуса позволит более глубоко понимать его роль в становлении иммунитета и формировании ряда важнейших адаптаций.
Различные стороны морфофункциональной организации иммунной системы в целом и тимуса в частности прошли длительный период формирования [14, 11]. Анализ современной научной литературы свидетельствует о недостаточной изученности вопроса, касающегося процессов преобразования сосудистого русла в ряду позвоночных животных, что создает основу для формирования противоречивых выводов. В складывающейся ситуации сравнительно-морфологическое изучение сосудистого русла тимуса различных представителей типа хордовые, может внести весомый вклад в решение целого ряда научных проблем.
Цель исследования - получить данные о плотности кровеносных сосудов мозговом и коркового веществе тимуса в сравнительно-морфологическом ряду неполовозрелых позвоночных.
Материал и методы исследования. Исследование тимуса проводили на примере 15-ти видов позвоночных, относящихся к четырем классам: класс Земноводные (Amphibia): Лягушка прудовая (Rana esculenta, n=18), Лягушка травяная (R. temporaria, n=14), Лягушка остромордая (R. terrestris, n=18); класс Пресмыкающиеся (Reptilia): Ящерица прыткая (Lacerta agilis, n=18), Веретеница ломкая (Anguis fragilis, n =6), Гадюка обыкновенная (Vipera berus, n=12), Уж обыкновенный (Natrix natrix, n=18); класс Птицы (Aves): Голубь сизый (Columba livia, n=18), Галка обыкновенная (Corvus monedula, n=6), Мухоловка серая (Muscicapa striata, n=8); класс Млекопитающие (Mammalia): Бурозубка обыкновенная (Sorex araneus, n=18), Бурозубка средняя (S. caecutiens, n=12), Рыжая полевка (Clethrionomys glareolus, n=23), Мышь лесная (Apodemus uralensis, n =6), Человек (Homo sapiens, n=28).
Возраст животных определялся по общепринятым методикам [8]. Исследование проводили на примере неполовозрелых особей: земноводные и пресмыкающиеся 1-2 года, птицы 1-3 года, насекомоядные млекопитающие 2-6 месяцев, грызуны 1-2 месяцев. Отлов животных осуществляли в экосистемах, не нарушенных антропогенным воздействием, на территории Национального природного парка «Смоленское Поозерье», Демидовский район, Смоленская область. Возраст человека определяли согласно классификации, принятой на 7-й Всесоюзной конференции по возрастной морфологии, физиологии и биохимии в 1965 году. Для исследования тимуса человека использовали материал, набранный на базе отделения клинической патологии при Смоленском областном институте патологии. Весь секционный материал тщательно отбирали по анамнезу с целью исключения причин смерти, которые могли бы существенно изменить структуру тимуса. Всего изучено 243 препарата неполовозрелых позвоночных. Эвтаназию животных осуществляли передозировкой эфирным наркозом (ЗАО Вектон) в соответствии с требованиями Министерства здравоохранения Российской Федерации к работе экспериментально-биологических клиник, а также «Европейской конвенции по защите позвоночных животных, используемых для экспериментов или в научных целях» (г. Страсбург, 1986). Тимус, изъятый сразу после эвтаназии, взвешивали и измеряли. Доли тимуса фиксировали 10% нейтральным формалином, обезвоживали и заливали в парафин по стандартной методике. Срезы тимуса (5 мкм) окрашивали гематоксилином и эозином, альдегид-фуксином и смесью Halmi по Габу-Дыбану.
Съемку препаратов проводили с использованием системы визуализации Sony Toup Сam 5.1. («Touptek», Китай), установленной на микроскопе Микромед 3 Professional («Микромед», Китай). Морфометрические исследования препаратов тимуса проводили с помощью компьютерной программы на базе цифровой камеры Toupcam 5.0. Общую площадь гистологического препарата тимуса измеряли при увеличении окуляра × 5, объектива × 4. В процессе тотального изучения всей площади препарата измеряли и определяли общую площадь сосудистого русла тимуса, которую затем выражали в % по отношению к площади среза (окуляр × 7, объектив × 20). С целью изучения отличий кровоснабжения мозгового и коркового вещества тимуса на условной единице площади (0,5 мм2) подсчитывали количество и площадь сосудов (мкм2) микроциркуляторного руса. Участки среза для подсчета сосудов выбирали в случайном порядке. На каждом препарате осуществляли подсчет на десяти таких участках. При идентификации звеньев микрососудистого русла за основу взяты общепринятые классификации [9, 15]. К микрососудам отнесены артериолы, прекапиллярные артериолы, капилляры, посткапиллярные венулы, венулы (окуляр × 15, объектив × 20; × 40). Значимость различий между сравниваемыми группами оценивали методами параметрической и непараметрической статистики (t-критерий Стьюдента, U-критерий Манна-Уитни и тест Краскела-Уоллиса).
Результаты исследования и их обсуждение. Как показывают полученные результаты, у холоднокровных позвоночных по сравнению с птицами и млекопитающими общая площадь кровеносного русла в тимусе снижена. Среди теплокровных представителей данный показатель наиболее развит у птиц и насекомоядных млекопитающих. Суммарная площадь всех сосудов тимуса у этих групп животных в 2-3 раза выше, чем у холоднокровных позвоночных, а также в 1,5 раза превышает площадь кровеносного русла грызунов и человека (табл.).
У всех неполовозрелых позвоночных за исключением человека диаметр артериол тимуса оказывается сходным и колеблется в узких пределах. Только у человека размер данных сосудов значительно превышает показатели животных (табл.). Исследование диметра венул тимуса показало, что у всех позвоночных их диаметр превышает аналогичные показатели артериол, что наиболее выражено в тимусе птиц (в 2 раза) и в меньшей степени в тимусе человека. У других позвоночных, независимо от уровня организации, просвет венул 1,5 раза шире, чем соответствующий показатель артериол. Наиболее крупным диаметром отличаются венулы тимуса человека, а также птиц. Значимых отличий в диаметре венул холоднокровных позвоночных и мелких млекопитающих не обнаружено (табл. 1).
По количеству сосудов МЦР коры тимуса все, изученные нами, группы холоднокровных
Таблица.
Некоторые планиметрические характеристики кровеносного русла тимуса неполовозрелых позвоночных (%)
|
Относительная S сосудистого русла (%) |
Ø артериолл (мкм2) |
Ø венул(мкм2) |
|
|
Земноводные |
0,80±0,22 c,d,e,f,g,h |
25,54±3,25 h |
38,80±4,27 d, e, h |
|
Ящерицы |
0,71±0,14 c,d,e,f,g,h |
24,61±3,49 h |
35,62±3,89 d, e, h |
|
Змеи |
1,04±0,18 d,e,f,g,h |
24,25±4,21 h |
35,41±4,51 d, e, h |
|
Голубеобразные |
2,12±0,31 a,b,c,g,h |
24,89±2,98 h |
56,71±4,68 a,b,c,f,g,h |
|
Воробьеобразные |
1,88±0,25 a,b,c,g,h |
29,10±4,32 h |
61,85±7,83 a,b,c,f,g,h |
|
Насекомоядные млекопитающие |
2,05±0,33 a,b,c,g,h |
26,25±3,51 h |
33,62±2,69 d, e, h |
|
Грызуны |
1,33±0,37 a,b,c,d,e,f |
28,19±4,11 h |
42,71±5,81 d, e, h |
|
Человек |
1,56±0,31 a,b,c,d,e,f,g |
70,73±10,82 a,b,c,d,e,f,g |
92,38±10,11 b,c,d,e,f,g |
Примечание: достоверность отличий (p≤0,05) по сравнению: a - с земноводными, b - с ящерицами, c - со змеями, d - с голубеобразными, e - с воробьеобразными, f - с грызунами, g - с насекомоядными млекопитающими, h - с человеком.
позвоночных сходны друг с другом и существенно уступают птицам, а так же млекопитающим, среди которых наиболее значительным данный показатель оказывается у человека (рис. 1). В мозговом веществе тимуса количество сосудов МЦР остается высоким только у млекопитающих, тогда как все холоднокровные представители ряда, а так же птицы отличаются одинаково низкими показателями.
Полученные результаты свидетельствуют о том, что у холоднокровных позвоночных количество сосудов, участвующих в обменных процессах, в коре и мозговом веществе тимуса оказывается сходным (рис. 1).
В свою очередь, изучение тимуса птиц и млекопитающих указывает на отличия в кровоснабжении функционально разных зон тимуса. При этом у птиц микроциркуляторное русло оказывается более развитым в коре тимуса, тогда как у млекопитающих, напротив – в его мозговой зоне (рис. 1).
Сопоставление сосудов МЦР коркового вещества тимуса различных групп позвоночных показало, что наименее развитой данная группа сосудов оказывается у земноводных и ящериц. Напротив, змеи и насекомоядные млекопитающие отличаются повышенными значениями их площади, тогда как для грызунов и человека характерны промежуточные показатели (рис. 2).
Примечательно, что у змей площадь мелких сосудов коры тимуса превышает показатели птиц, грызунов и человека в 1,3 раза, а земноводных и ящериц почти вдвое.
Изучение площади сосудов МЦР мозгового вещества тимуса свидетельствует о том, что у холоднокровных позвоночных, а также у птиц данные показатели снижены. Напротив млекопитающие отличаются повышенными значениями площади обменных сосудов мозгового вещества, что особенно сильно выражено у насекомоядных млекопитающих. По состоянию МЦР русла мозгового вещества тимуса змеи оказываются сопоставимы с млекопитающими, но не с холоднокровными позвоночными, к которым они принадлежат.
При сопоставлении коркового и мозгового вещества тимуса выявлено, что у змей и птиц площадь сосудов микроциркуляторного русла более развита в коре тимуса, тогда как у млекопитающих в его мозговой зоне. В свою очередь, у холоднокровных тетрапод (земноводные и ящерицы) отличий в площади микроциркуляторного русла коркового и мозгового вещества тимуса не обнаружено.
Изучение капилляров тимуса неполовозрелых позвоночных животных показало, что наименьший диаметр данных сосудов характерен для мелких насекомоядных и грызунов: соответ-
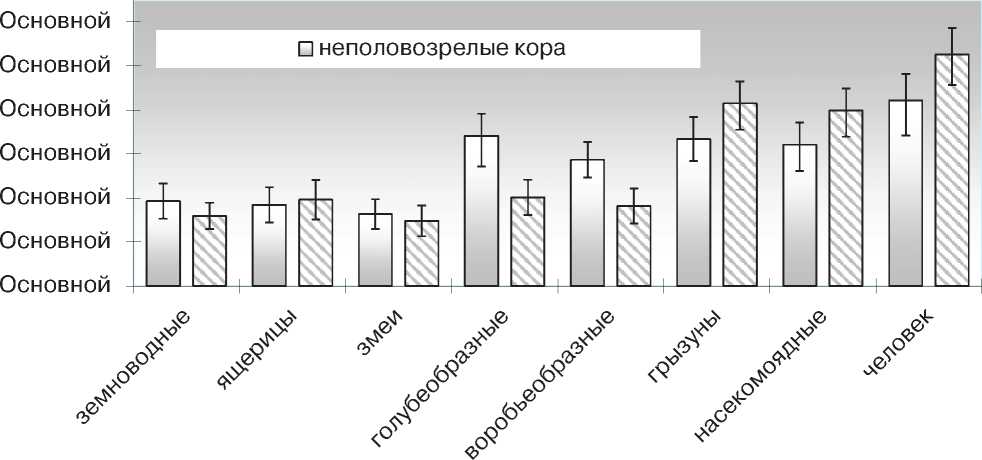
Рис. 1 Количество сосудов микроциркуляторного русла на условной единице площади (0,5 мм2) коркового и мозгового вещества тимуса неполовозрелых позвоночных.
Основной Основной Основной Основной Основной Основной Основной Основной Основной
x0
x^
O'
z

Рис. 2. Площадь сосудов микроциркуляторного русла (в мм2) на условной единице площади (0,5 мм2) мозгового и коркового вещества тимуса неполовозрелых позвоночных.
ственно 4,2±0,12 и 5±0,10 мкм. Максимальным просветом капилляров отличается тимус человека (6,3±0,54 мкм) и земноводных (7,4±0,7 мкм), тогда как у пресмыкающихся и птиц размеры капилляров отличаются мало и изменяются в пределах 5±0,34 – 6±0,31 мкм. У животных и человека просвет капилляров тимуса оказывается на 1-1,5 мкм меньше, чем диаметр эритроцитов. Для эл- липсовидных эритроцитов птиц, земноводных и пресмыкающихся сопоставлялись данные меньшего диаметра эритроцитов.
Размеры пре- и посткапиллярных сосудов тимуса у неполовозрелых позвоночных изменяются в узких пределах: от 11,5±3,2 мкм (пресмыкающиеся), до 14,4±3,8 мкм (грызуны). Лишь только у человека размер сосудов этой группы оказался повышенным (18,2±4,4 мкм). Статистическая обработка результатов показала значимость отличий при сравнении холоднокровных позвоночных с теплокровными, за исключением насекомоядных млекопитающих и голубеобразных птиц.
Сравнительно-морфологический ряд позвоночных животных является надежной моделью, в рамках которой происходило поступательное совершенствование механизмов иммунной защиты [14, 11, 13]. Изменения морфологических характеристик кровеносного русла тимуса, выявленные в рамках нашего исследования, свидетельствуют о том, что сосудистое русло непосредственным образом задействовано в данных процессах. Развивающаяся теплокровность предъявляет повышенные требования к тимусу, что приводит к усилению гормональной регуляции иммуногенеза [7, 12], ускоряет темпы транспорта и созревания предшественников тимоцитов [10, 18], а также стимулирует возрастание совокупной площади гемато-тимического барьера [19].Такие изменения не возможны без соответствующих морфологических трансформаций сосудистого русла тимуса, которые мы наблюдаем в процессе сравнительно-морфологических исследований. В силу пластичности кровеносного русла наблюдается зависимость его морфологии от уровня активности животного, что, по нашему мнению, приводит к отличиям количественных характеристик сосудов у птиц и насекомоядных млекопитающих, образ жизни которых обуславливает повышение энерготрат. Несмотря на то, что в целом уровень организации птиц и млекопитающих оказывается сопоставимым, все же эти группы животных представляют собой разные эволюционные ветви, отличающие по срокам возникновения и степени накопления рептильных черт в рамках морфофункциональной конституции. Эти условия приводят к отличиям в кровоснабжении коркового и мозгового вещества тимуса птиц и млекопитающих, а значит способствуют формированию характерных особенностей иммунитета, связанных с транспортом и созреванием тимоцитов.
Высокий уровень специализации животного, формирующийся в процессе возникновения особой жизненной формы, так же способствует развитию кровеносного русла тимуса, а значит затрагивает и иммунные процессы. Наглядным примером тому являются змеи (безногая жизненная форма) у которых по некоторым показателям кровеносное русло тимуса более сопоставимо с птицами и млекопитающими, но не с холоднокровными позвоночными, к которым они принадлежат.
Исследования кровеносной системы тимуса лабораторных млекопитающих свидетельствуют о более интенсивном кровоснабжении его коркового слоя по сравнению с мозговым [5]. Однако данные, полученные в нашем исследовании, на примере обитателей естественных природных биотопов, свидетельствуют об обратном. У всех изученных групп мелких млекопитающих ми-кроциркуляторное русло преобладает именно в мозговом веществе. Нечто подобное наблюдали и другие исследователи, изучавшие тимус животных, изъятых из природных биотопов [6]. Такая ситуация свидетельствует о сильной зависимости тимуса в целом и его кровеносной системы от условий среды обитания [10, 17].
Морфология различных сосудов микроцир-куляторного русла тимуса позвоночных животных определяется закономерностями обменных процессов в тканях. Общее сходство таких процессов и приводит к идентичности размеров артериол и венул, а также пре и пост капиллярных сосудов. Напротив, отличия диаметра капилляров тимуса позвоночных обусловлено размерами эритроцитов, что также связано с явлениями обмена веществ между кровью и тканями. Такое несоответствие размеров способствует трению эритроцита о стенки капилляра, что существенно усиливает диффузный обмен.
Таким образом, отличия в морфологии сосудистого русла тимуса позвоночных животных возникают под влиянием важнейших адаптаций, связанных как с совершенствованием уровня организации в целом, так и с приспособлением к конкретным условиям существования.
Список литературы Изменения внутриорганного сосудистого русла тимуса в сравнительном орфологическом ряду неполовозрелых позвоночных
- Аминова, Г. Г. Морфологические основы регуляции кровотока в микроциркуляторном русле/Г. Г. Аминова//Регионарное кровообращение и микроциркуляция. -2003. -№ 4. -С. 80-84.
- Бабкина, И. В. Организация сосудистого русла тимуса в постнатальном периоде онтогенеза/И. В. Бабкина//Актуальные проблемы человека и животных. Матер, науч. конф. УГПУ. Ульяновск. -1996. -С. 18-19.
- Банин, В. В. Механизмы обмена внутренней среды/В. В. Банин -Монография. М.: Изд -во РГМУ, 2000. -278 с.
- Васильев, А.П. Микроциркуляторная картина у здоровых и больных артериальной гипертензией с различными вариантами периферической гемодинамики/А. П. Васильев, Н. Н. Стрельцова, М. А. Секисова//Российский кардиологический журнал. -2011. -№ 1 (87). -С. 34-39.
- Волошин, Н. А. Особенности микроциркуляторного русла коркового и мозгового вещества вилочковой железы крыс/Н. А. Волошин, М. Е. Иванов//Актуальные вопросы морфогенеза сердца: Материалы научного симпозиума, посвященного 80 -летию кафедры анатомии человека ДГМА. Днепропетровск. -1996. -С.43-45.
- Грушко, М. П. Особенности гистологической организации некоторых органов кроветворения озерной лягушки (Rana ridibunda)/М. П. Грушко//Вестник Астраханского государственного технического университета. -2010. -№ 1. -С. 78-80.
- Кветной, И. М. Нейроиммуноэндокринология тимуса//И. М. Кветной, А. А. Ярилин, В. О. Полякова. СПБ: Издательство ДЕАН. -2005. -160 с.
- Клевезаль, Г. А. Принципы и методы определения возраста млекопитающих//Г. А. Клевезаль. М.: КМК. -2007. -288 с.
- Куприянов, В.В. Пути микроциркуляции//В. В. Куприянов. Изд-во «Картя Молдовенскэ», Кишинев. -1969. -254с.
- Пасюк, А. А. Характеристика кровеносного русла тимуса белой крысы в эмбриогенезе/А. А. Пасюк, П. Г. Пивченко//Вестник ВГМУ. -2006. -Том. 5. -№ 3. -С. 2-6.
- Пестова, И. М. Морфофункциональная организация системы гемииммунопоэза в эволюции/И. М. Пестова, В. А. Четверных//Архив анатомии, гистологии и эмбриологии. Ленинград, -1990. -Т. 99. -№ 11. -С. 90-99.
- Полетаева, А. В. Влияние гормонов на иммунологическую реактивность/А. В. Полетаева, А. И. Леванюк, Е. В. Сергеева//Вопросы экологической эндокринологии. -№7. -2009. -С. 42-46.
- Селезнев, С. Б. Эволюция лимфоидных структур позвоночных/С. Б. Селезнев//Морфология. -2006. -Т. 129. -№ 4. -С. 111.
- Флоренсов, В. А. Очерки эволюционной морфологии/В. А. Флоренсов, И. М. Петрова//Иркутск: Изд-во Иркутского ун-та, -1990. -245 с.
- Чернух, А. М. Микроциркуляция/А. М. Чернух, П. Н. Александров//монография М.: Медицина, -1984. -432 с.
- Mori, K. The perivascular spaceasa path of hematopoetic progenitor cells and mature T cells between the blood circulation and thymic parenchyma/K. Mori, M. Itoi, N. Tsukamoto//Int. Immunol. -2007. -19(6). -P. 745-753.
- Nanka, O. Experimental hypoxia and embryonic angiogenesis/O. Nanka, P. Valasek, M. Dvorakova, M. Grim//Development dyndmics. -2006. -Vol. 235. -P. 723-733.
- Pearse, G. Normal structure, function and histology of the thymus/G. Pearse//Toxicol. Pathol. 2006. -34. -(5). -P. 504-514.
- Roberts, R.L. Transport of transferrin across the blood-thymus barrier in young rats/R. L. Roberts, A. Sandra//Tissue Cell. -1994. -Vol. 26. -P. 757-766.
