Изучение генетической подразделенности приханкайской популяции дикой сои (Glycine soja Sieb. & Zucc., 1845)
")
Автор: Недолужко А.В., Тихонов А.В., Дорохов Д.Б.
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Ботаника и физиология растений
Статья в выпуске: 2, 2008 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14718991
IDR: 14718991
Текст статьи Изучение генетической подразделенности приханкайской популяции дикой сои (Glycine soja Sieb. & Zucc., 1845)
А. В. Недолужко,А. В. Тихонов,Д. Б, Дорохов
Изучение генетического разнообразия популяций дикорастущих родичей культурных растений — одна из важнейших проблем биологии. Данные о генетической структуре популяций необходимы для разработки мер по сохранению уникальных генетических ресурсов, которые могут быть использованы в селекционных работах по улучшению хозяйственно-ценных признаков культурных растений.
Юг российского Дальнего Востока (Амурская область, Приморский и Хабаровский край) считают одним из центров генетического разнообразия дикой сои (G. soja), ближайшего родича культурной сои (Glycine max (L.) Mere., 1917). Контрастность местных агроклиматических условий делает дальневосточные формы G. soja особенно ценными донорами для современных селекционных программ, направленных на повышение адаптивного потенциала новых сортов. В настоящее время накоплен большой опыт и показана перспективность использования генов из естественных популяций G. soja Дальнего Востока. Это дает реальную возможность для улучшения уже существующих и создания новых сортов культуры [1].
Наиболее часто на российском Дальнем Востоке дикая соя встречается вблизи озера
Ханка. Ее проективное покрытие здесв может достигать 90 %. Исследования позволили выяснить, что дикая соя, произрастающая на Приханкайской равнине, обладает повышенным генетическим разнообразием по сравнению с другими популяциями Дальнего Востока [4; 5]. Уровень ожидаемой гетерозиготности у G. soja варьирует от 0,1460 до 0,2780 в разных субпопуляциях. Высокое генетическое разнообразие популяции дикой сои, произрастающей на Приханкайской равнине, послужило поводом для выбора данного региона в качестве модельного для последующих исследо- i ваний генетической структуры популяций ' G. soja на юге Дальнего Востока [5].
Для изучения генетической подразделенности приханкайской популяции G. soja был использован семенной материал (67 образцов), собранный сотрудниками Центра »Био-инженерия» РАН в период экспедиционных работ в окрестностях оз. Ханка Приморского края (рис. 1). Список материала представлен в табл. 1.
Нами был проведен анализ бинарной матрицы, полученной по результатам электрофореза продуктов амплификации праймерами: j ОРА-08 (GTGACGTAGG), ОРА-09 i (GGGTAACGCC), ОРА-04 (AATCGGGCTG), ОРН-12 (ACGCGCATGT). ДНК для проведе-
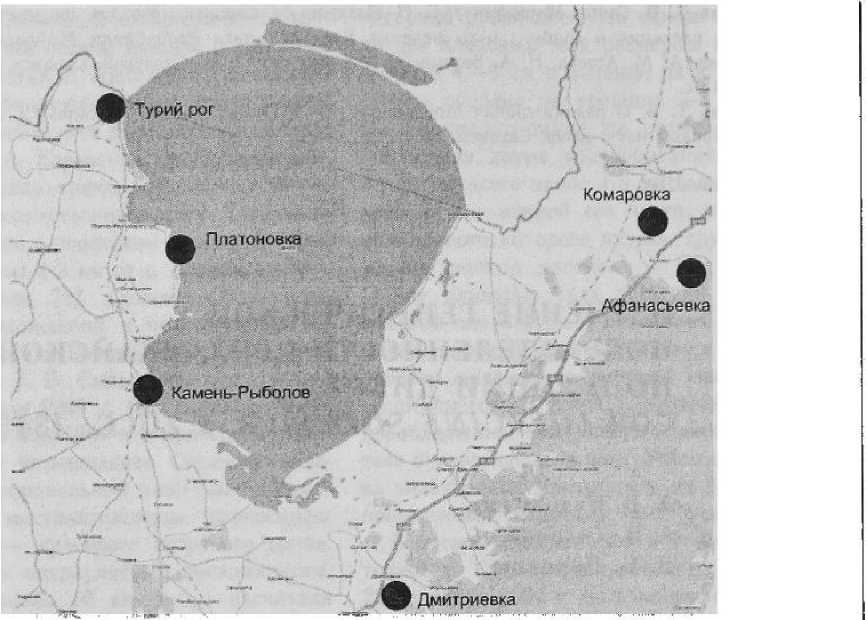
Рисунок /
Места сбора материала при проведении работ по изучению популяции G. soja в окрестностях оз. Ханка Приморского края ния амплификации выделяли из первых настоящих листьев молодых проростков по протоколу Эдвардса и др. [8], модифицированному Д. Б. Дороховым и Э. Клоке [2]. При пост-ППШ1ТЛГ fllTtrriT41inTI liTOTnirTfL! ГТ ПТ1ОиПЪТ1Л< » 1 хХ р» W4 umiujyii Vil 1'1 и 1 yi 1 i-\i-,i i ijvnx'iii^uv'-'i »-^ ч считалась каждая яркая полоса ДНК на геле; ампликоны, не ясно различимые на геле, не учитывались. Отсутствие признака обозначалось как «О». Матрица была обработана при помощи программы PopGene32. Полученные генетические показатели были использованы для сравнения популяций, произрастающих на восточном и западном побережье оз. Ханка Приморского края, при помощи метода корре ляционных плеяд [6]. Этот метод позволяет изучить связи набора показателей и установить наиболее важные из них (индикаторы) [7].
Нами были вычислены коэффициенты корреляции между полученными генетическими характеристиками для популяций дикой сои Приханкайской равнины и реки, затем построены корреляционные плеяды для разных уровней достоверности (р < 0,05, р < 0,01 и р < < 0,001), коэффициенты корреляции были вычислены с помощью программы Statist!са 6.0.
При изучении генетического полиморфизма популяций дикой сои на территории При-
■ Таблица /
Объем и источники материала дикой и культурной сои
Китайские коллеги, исследовавшие генетическую структуру естественных популяций Северного Китая при помощи lSSR-анализа, показали, что уровень полимофности (.Р ) в них варьирует от 68,2 до 72,0 %, а гетерозиготность трех изученных ими популяций G. so/а составила 0,2450, 0,2590 и 0,2620 соответственно (9]. Отметим, что ранее была оценена эффективность RFLP, RAPD, AFLP и SSR-маркеров при изучении генетического разнообразия образцов дикой и культурной сои [12]. Измерив, ожидаемую гетерозиготность, авторы показали, что микросаттелитные маркеры дают наибольшие значения Не показали SSR-маркеры — 0,60 [12]. В это же время отмечено, что при оценке внутри- и межпопуляционной изменчивости у покрытосеменных растений величины, полученные при использовании доминантно-наследуемых генетических маркеров (RAPD, ISSR, AFLP), сопоставимы между собой [11]. Полученные нами показатели в целом хорошо соотносятся с данными китайских коллег [9], несмотря на различные типы использованных генетических маркеров.
Таким образом, можно сделать вывод, что популяции дикой сои, произрастающие на территории Северного Китая и в центральном Приморье, обладают примерно одинаковым уровнем генетического разнообразия.
В характере построенных корреляционных плеяд для образцов восточного и западного побережья оз. Ханка обнаруживаются существенные различия (рис. 2). Данные корреляционного анализа указывают на наличие особенностей микроэволюционных процессов на уровне геномов в субпопуляциях, расположенных в разных частях видового ареала и находящихся под действием разных факторов естественного отбора, хотя величина генетических дистанций [10] между образцами с восточного и западного побережья озера невелика — 0,0249. Ожидаемая гетерозиготность для генотипов западного побережья составила — 0,2953, а для генотипов восточного — 0,2712.
Мы полагаем, что дифференциация субпопуляций О. soja складывается в процессе длительного воздействия метеорологических
Таблица 2
Параметры внутрипопуляционной изменчивости
|
Субпопуляция |
Не |
I |
Р, % |
пр! |
||
|
с, Афанасьевка |
1,7419 (0,4411) |
1,4153 (0,3460) |
0,2494 |
0,3771 (0,2623) |
74,19 |
46 |
|
с. Комаров ка |
1,7258 (0,4497) |
1,4115 (0,3692) |
0,2424 |
0,3654 (0,2708) |
72,58 |
45 |
|
с. Дмитриевка |
1,6774 (0,4713) |
1,3834 (0,3474) |
0,2311 |
0,3493 (0,2708) |
67,74 |
42 |
|
с. Платоиовка |
1,7742 (0,4215) |
1,4418 (0,3816) |
0,2564 |
0,3857 (0,2670) |
77,42 |
48 |
|
пос. Турий Рог |
1,7097 (0,4576) |
1,4320 (0,4064) |
' 0,2445 |
0,3640 (0,2848) |
70,97 |
44 |
|
пос. Камень-Рыболов |
1,7903 (0,4104) |
1,4709 (0,3736) |
0,2729 |
0,4077 (0,2644) |
79,03 |
49 |
Примечание: па — наблюдаемое число аллелей на локус; пг — эффективное число аллелей на локус; Не — средняя ожидаемая гетерозиготность по всем локусам; Р — доля полиморфных локусов; / — индекс гетерогенности выборки по Шеннону; в скобках среднее квадратичное отклонение факторов, характеризующихся неоднородностью для западного и восточного побережья оз. Ханка [3]. Климатические условия При-ханкайской равнины определяются в основном циркуляционными процессами, роль солнечной радиации сказывается здесь в меньшей степени. Некоторое влияние на особенности местных климатических условий оказывает рельеф: долина оз. Ханка ограничена горными хребтами Сихотэ-Алиня с востока и Восточно-Маньчжурским нагорьем с запада. При отсутствии ветра Приханкайская равнина хорошо прогревается, зимой же она является аккумулятором холодных воздушных масс. Тем не менее климатические показатели восточного и западного побережья отличаются — это относится к влажности, среднегодовому количеству осадков, времени наступления первых и последних заморозков, а также среднегодовой температуре. Например, среднегодовая температура на западном побережье составляет 3,4 °C (Астраханка), а на восточном колеблется от 2,6 °C (Новосельское) до 1,6 °C (Кировский) [3].
Следует отметить, что существенный вклад в генетическую структуру популяции дикой сои могли внести разная степень хозяйственного освоения этих территорий и рельеф. Вероятно, полному разделению популяции препятствует антропогенная деятельность: дикая соя использует обочины дорог в качестве искусственных коридоров для своего распространения, что приводит к постоянному смешению генофондов субпопуляций в этом регионе.
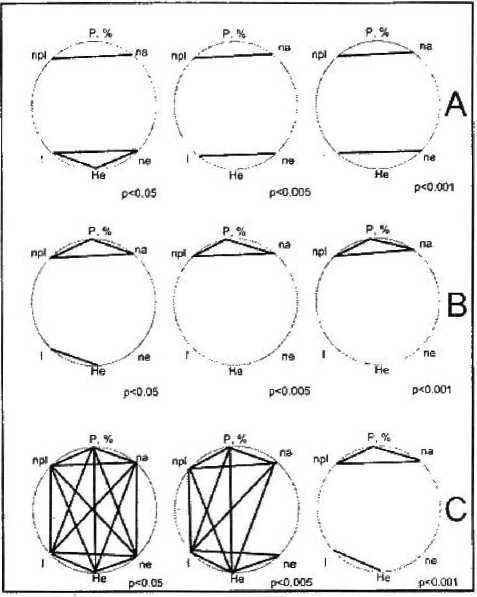
Рисунок 2
Корреляционная структура молекулярногенетических характеристик Glycine soja.
А — бостонное побережье оз. Ханка;
В — западное побережье оз. Ханка;
С — обитая плеяда для популяции оз. Ханка, пе, па, h, I, Не. Р — основные генетические параметры
Следующим этапом исследований мы видим изучение динамики «молодых» популяций дикой сои, сформировавшихся на заброшенных рисовых чеках в конце 1980-х гг.
Список литературы Изучение генетической подразделенности приханкайской популяции дикой сои (Glycine soja Sieb. & Zucc., 1845)
- Ала А. Я. Соя: генетические методы селекции G. max (L.) Merr. Ч G. soja/А. Я. Ала, B. А. Тильба. Благовещенск: ПКИ «Зея», 2005. 128 с.
- Дорохов Д. Б. Быстрая и экономичная технология RAPD анализа растительных геномов/Д. Б. Дорохов, Э. Клоке//Генетика, 1996. Т. 33. с. 358-365.
- Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Приморский край. Л.: Гидрометеоиздат. 1988. Ч. 1-6, вып. 26. 416 с.
- Недолужко А. В. Исследование генетической структуры популяций дикорастущей сои (G. soja Sieb. & Zucc.) Приханкайской равнины/А. В. Недолужко//Биотехнология -охране окружающей среды. Москва, 2005. С. 342-344.
- Недолужко А. В. Изучение биобезопасности генетически модифицированной сои в центре происхождения и разнообразия на Дальнем Востоке Российской Федерации/А. В. Недолужко, Д. Б. Дорохов//Цитология и генетика, 2007. № 3. С. 72-85.
- Терентьев П. В. Метод корреляционных плеяд/П. В. Терентьев//Вестн. ЛГУ. 1959. № 9. C. 137-141.
- Шереметьева И. Н. Оценка генетического разнообразия островных и материковых популяций дальневосточной полевки Microius jortis (Rodentia, Cricetidae): данные RAPD-PCR анализа/И. Н. Шереметьева, Г. Н. Челомина//The North Pacific island biological researches, 2003. V. 9. p. 1 -18.
- Edwards К. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis/K. Edwards, C. Johnstone, C. Thompson//Nucleic Acids Res, 1991. V. 19 (6). P. 1349.
- Jin Y. Genetic spatial clustering: significant implications for conservation of wild soybean (Glycine soja: Fabaceae)/Y. Jin, T. He, B.-R. Lu//Genetica. 2006. V. 128. P. 41-49.
- Nei M. Estimation of average heterozygosity and genetic distance from small number of individuals/M. Nei//Genetics. 1978. V. 89. P. 583-590.
- Nybom H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants/H. Nybom//Molecular Ecology, 2004. V. 13. p. 1143 -1155.
- The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis/W. Powell, M. Morgante, C. Andre, M. Hanafey, J. Vogel, S. Tingey, A. Rafalski//Molecular Breeding, 1996. V. 2 (3). P. 225-238.
