Изучение химической структуры лигнинов родиолы розовой (Rhodiola rosea L.) и серпухи венценосной (Serratula coronata L.) методом 2D ЯМР-спектроскопии
 и серпухи венценосной (Serratula coronata L.) методом 2D ЯМР-спектроскопии")
Автор: Белый В.А., Алексеев И.Н., Садыков Р.А.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Химические науки
Статья в выпуске: 3 (11), 2012 года.
Бесплатный доступ
Впервые выделены лигнины растений родиолы розовой (Rhodiola rosea L.) и серпухи венценосной (Serratula coronata L.). Методом двумерной ЯМР- спектроскопии определены общие закономерности химического строения. По- казано, что изучаемые лигнины построены из гваяцильных, сирингильных и пара-кумаровых фрагментов, соединенных, преимущественно, -О-4-связью. Установлены структурные различия лигнинов. В частности, выявлено, что ацетатные группы и кумаратовые фрагменты в лигнине родиолы розовой присутствуют только в -положении фенилпропановых единиц. Предшествен- никами лигнина родиолы розовой могут быть кумараты и ацетаты конифери- лового и синапового спиртов.
Лигнин, фенилпропановая единица, phenyl-propanе unit, двумерная ямр-спектроскопия
Короткий адрес: https://sciup.org/14992544
IDR: 14992544 | УДК: 547.022
Studying of chemical structure of lignins of Rhodiola rosea L. and Serratula coronata L. by 2D NMR-spectroscopy
Lignins of plants Rhodiola rosea L. and Serratula coronata L. are extracted for the first time. By the method of two-dimensional NMR-spectroscopy the general regularities of chemical structure are defined, it is shown that studied lignins are built of quaiacic, syringic and p-coumaric fragments connected, mainly, by - О-4 binding. Structural distinctions of lignins are established, in particular, it is shown that acetated groups and coumaric fragments in lignin of Rhodiola rosea L. are present only at -position of phenyl-propane units. It is established that coumarates and acetates of coniferyl and sinapic spirits can be predecessors of lignin of Rhodiola rosea L.
Текст научной статьи Изучение химической структуры лигнинов родиолы розовой (Rhodiola rosea L.) и серпухи венценосной (Serratula coronata L.) методом 2D ЯМР-спектроскопии
Лигнин – это структурный компонент травянистых и древесных растений, образующийся путем полимеризации кониферилового, синапового и п-кумарового спиртов, придающий жесткость клеточным стенкам, а также стойкость к воздействию микроорганизмов [1]. Особенность лигнинов как особой группы биополимеров состоит в неупорядоченности их химической структуры, проявляющейся в большом разнообразии ди- и тримерных фрагментов, составляющих структуру. Однако лигнины растений различных видов характеризуются определенными особенностями строения, точнее, структура лигнина на каждом уровне структурной организации (как на молекулярном, так и на топологическом) определяется его ботаническим происхождением [2]. Особенностям химического строения и физикохимическим свойствам лигнинов травянистых рас- тений до настоящего времени уделялось мало внимания, а данные по лигнинам лекарственных растений отсутствуют вообще.
Родиола розовая ( Rhodiola rosea L.) – многолетнее травянистое растение семейства толстян-ковых, произрастающее на Алтае, Урале, заполярных районах Якутии, в горных районах Восточной Сибири и Дальнего Востока. В медицине используют ее экстракты, обладающие стимулирующим и адаптогенным действием, применение которых направлено на улучшение физической выносливости, внимания, памяти, а также резистентности организма [3].
Серпуха венценосная ( Serratula coronata L.) – многолетнее травянистое растение семейства Asteraceae, встречающееся в европейской части России, на Кавказе, в Западной Сибири, Дальнем
Востоке и Средней Азии. Серпуха венценосная является источником фитоэкдистероидов – растительных гормонов, обладающих целым рядом ценных фармакологических свойств [4]. Исследования низкомолекулярной, экстрактивной составляющей родиолы розовой и серпухи венценосной представлены в работах [5, 6].
Поиск путей рационального использования лекарственного растительного сырья, в частности, родиолы розовой и серпухи венценосной, требует детального изучения структуры их лигнинов.
Спектроскопические методики, и особенно 1D и 2D ЯМР, дают исчерпывающую информацию о структуре макромолекул и являются мощными инструментами для исследования лигнинов различного ботанического происхождения. В этой статье представлены результаты исследования химической структуры лигнинов родиолы розовой и серпухи венценосной с использованием 2D ЯМР-спект-роскопии.
Материалы и методы
Образцы. Сырьем для извлечения лигнина в данной работе являлись корневища родиолы розовой, которые заготавливали с конца цветения до конца вегетации растения, и стебли серпухи венценосной. Растения выращены и собраны в Ботаническом саду Института биологии Коми НЦ УрО РАН (г. Сыктывкар, Республика Коми).
В исходных образцах растений по общепринятым методикам [7] предварительно было определено содержание лигнина, целлюлозы и других компонентов (табл. 1).
Таблица 1
Компонентный состав родиолы розовой и серпухи венценосной
|
Растение |
Целлюлоза, % |
Лигнин, % |
Зольность, % |
Легкогид-ролизуе-мые вещества, % |
Экстрактивные вещества, % |
|
Rhodiola rosea L. |
9,0±0,2 |
21,1±0,4 |
1,9±0,1 |
36,2±0,6 |
31,8±0,5 |
|
Serratula coronata L. |
44,9±0,7 |
16,3±0,4 |
7,1±0,1 |
25,1±0,4 |
6,6±0,1 |
Предварительная подготовка растительного сырья заключалась в экстракции водой и спирто-бензольной смесью. Образцы лигнинов серпухи венценосной (ДЛС) и родиолы розовой (ДЛР) получали экстрагируя растительное сырье смесью диоксан-вода 9:1 в присутствии HCl2 (0,7 %) при температуре кипения в течение 2 ч. Полученный раствор лигнина упаривали в вакууме и высаживали в 1 %-ный раствор сульфата натрия. Лигнин отмывали от соли, подсушивали на воздухе, затем переосаждали из диоксанового раствора в диэтиловый эфир и сушили в вакууме. Выход лигнина серпухи венценосной составил 22 %, родиолы ро- зовой – 30 % от содержания в исходном растительном сырье.
Двумерные спектры ЯМР регистрировали при 25°С в 5 мм ампулах на спектрометре Bruker AVANCE II 300, рабочая частота 300 МГц (1Н) и 75 МГц (13С). Около 80 мг лигнина растворяли в 0,6 мл дейтерированного диметилсульфоксида, затем 2D ЯМР спектры записывались в HSQC (Heteronuclear Single Quantum Correlation), COSY (Correlation Spectroscopy) и HMBC (Heteronuclear Multiple Quantum Coherence) экспериментах. Ширина спектров составила 5 тыс. и 20 тыс. Гц в 1Н и 13С измерениях соответственно. Число накоплений составляло 20 тыс. с задержкой 5 с. В качестве внутреннего стандарта использовался сигнал растворителя (δ С 40.1; δ Н 2.5). Кросс-сигналы спектров HSQC интерпретировались по литературным данным [8, 9]. При регистрации 13С-спектров время релаксации искусственно уменьшалось добавлением трис-ацетил-ацетоната хрома (0.02 моль/л).
Результаты и обсуждение
Лигнины, выделенные диоксаном по описанной выше методике, часто используются для структурных исследований, поскольку они являются наиболее представительной частью всего лигнина в растении [10]. Поэтому в данной работе для исследования структуры лигнинов лекарственных растений применяли именно диоксановый метод выделения образцов. Однако необходимо иметь в виду, что выводы, сделанные по выделенным препаратам, могут иметь лишь предположительный характер в отношении всего лигнина, находящегося в растении.
Строение лигнина характеризуется наличием ароматических ядер, замещенных метоксильными, гидроксильными функциональными группами и алкильными цепочками. Описание характеристик лигнина как вещества нерегулярного строения возможно через определение его фрагментного состава, демонстрирующего статистические параметры химической структуры лигнина.
Ароматические фрагменты. Наиболее интенсивные корреляционные сигналы в ароматическом регионе HSQC-спектров (δ Н /δ С 5.5-8.5/96-150 м.д.), представленных на рис. 1, соответствуют ароматическим кольцам различных структурных единиц лигнина. В HSQC-спектрах шкала химических сдвигов атомов 1Н отложена по оси абсцисс, атомов 13С – по ординат. Сигнал 6.68/103.8 м.д. на рис. 1, согласно [9], соответствует атомам во втором и шестом положениях сирингильного кольца (рис. 2 S), для сирингильных структурных единиц, имеющих карбонильную группу в α-положении, характерен сигнал 7.36/104.0 м.д. [9], присутствующий на спектрах ДЛР (рис. 1 а) и ДЛС (рис. 1 б). Присутствие в образцах п -кумаровых звеньев (рис. 2 H) подтверждается корреляционным сигналом δ Н /δ С 7.28/127.5 м.д [9].
Хорошо разрешенные сигналы п -кумарато-вых фрагментов (рис. 2 Y) наблюдаются в спектрах лигнина родиолы розовой (рис. 1 а). Кросс-сигналы δ Н /δ С 7.5/145.0 м.д и 6.5/115.0 м.д. (рис. 1 а) соот-
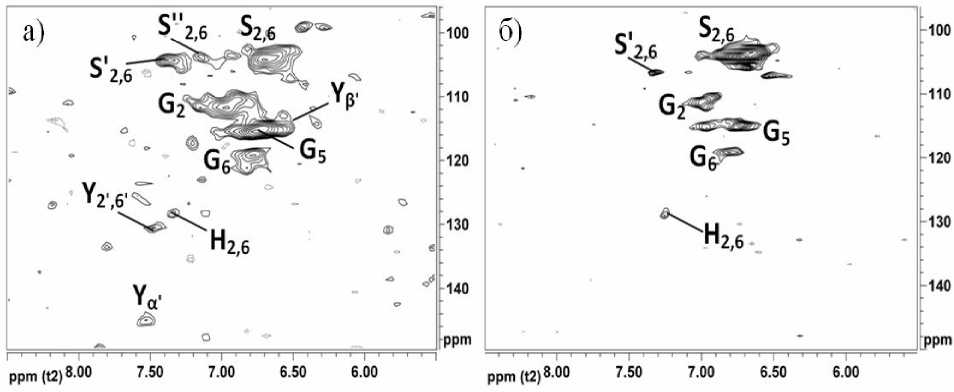
Рис. 1. Область сигналов ароматических атомов на HSQC-спектрах ДЛР (а) и ДЛС (б).
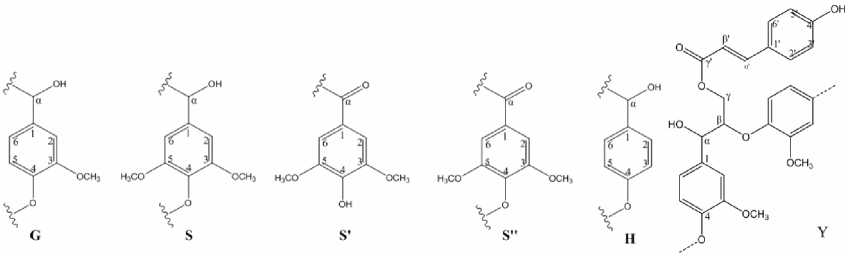
Рис. 2. Мономерные структурные единицы лигнинов: G – гваяцильные, S – сирингильные, H – п -ку-маровые, Y – ацилированные п -кумаровой кислотой.
ветствуют α (рис. 2 Y α' ) и β (рис. 2 Y β' ) атомам боковой цепочки п -кумаратов [9], а сигналы при δ Н /δ С 7.40/130.5 м.д. и 6.77/116.2 м.д. относятся к ароматическому кольцу п -кумаратов (рис. 2 Y 2', 6' ) и (рис. 2 Y 3', 5' ) соответственно [9].
Во многих исследованиях лигнинов травянистых растений указывалось на присутствие в структуре остатков п-кумаровой кислоты, соединенной с фенилпропановыми единицами сложноэфирной связью [11–14]. В этих работах предполагались два различных пути вовлечения п-кумаровой кислоты в процесс биосинтеза – либо этерификация п-кумаровой кислотой спиртовых групп уже собранной макромолекулы лигнина, либо ацилирование монолигнолов с образованием п-кумаратов синапо-вого и кониферилового спиртов. Для решения этого вопроса требовалось определение вариантов присоединения п-кумаратов к фенилпропановым единицам. ЯМР исследования лигнина, выделенного из кукурузы [15], пшеницы [16] и некоторых других трав [8, 17], показали, что п-кумараты присутствуют только в γ-положениях звеньев лигнина. Эти результаты однозначно указывали на то, что этерификация происходила до образования макромолекулы лигнина. Теперь широко признано, что предшественниками лигнинов могут быть сложные эфиры гидроксокоричных спиртов и п-кумаровой или бензойной кислот, а также ацетаты конифери- лового и синапового спиртов [18–21]. Ацилированные в γ-положении фрагменты были определены в лигнинах многих растений [18, 19].
Идентификация п -кумаратов в ДЛР представлена на рис. 3. Спектр ЯМР 13С (рис. 3 а) демонстрирует сигналы четвертичных атомов углерода, на наличие п -кумаратов указывает сигнал γ'-атома 167.4 м.д [9]. HSQC- и COSY-спектры (рис. 3 б) также подтверждают наличие п -кумаратов в ДЛР, в представленной на них области отмечены сигналы α' и β' атомов. Для выяснения точки присоединения п -кумаратов к структурным единицам лигнина на спектре (рис. 3 в) были выделены области сигналов, характерные для структурных единиц с присоединенными в α-положении п -кумаратовыми фрагментами (α- p CA), а также в γ-положении во фрагментах Y (рис. 2). По результатам анализа спектров на рис. 3 определено, что фрагменты п -кумарата присоединены в ДЛР только в γ-положении фенилпропановых единиц (δ Н /δ С 4.1/64.5 м.д.) в структурах, подобных Y, поскольку не обнаружено сигналов α-атома, образующего связь с п -кумаратом (δ Н /δ С 6.1/75.0 м.д. [9]). Для лигнина серпухи венценосной (образец ДЛС) наличие п -кумаратов не характерно.
В спектрах ДЛР выявлены сигналы ацетатных групп (δ Н /δ С 1.7/20.7 и δ С 170.7 м.д. [9]). На рис. 3 в отмечены сигналы атомов боковой цепочки фенилпропановых единиц, связанных с ацетатными
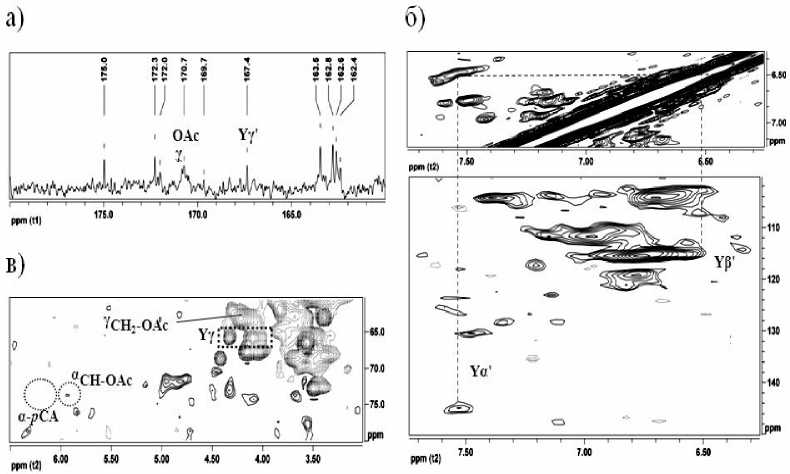
Рис. 3. Определение п-кумаратов в ДЛР по HSQC- и COSY-спектрам.
группами: в γ-положении (δ Н /δ С 4.2/63.0 м.д. [9]), а также в α-положении (δ Н /δ С 5.9/74.7 м.д. [9]). Связь с ацетатами в α-положении выявляется в лигнинах травянистых растений сравнительно редко, чаще встречается исключительно в γ-положении [17]. И, согласно предположению, выдвинутому Ральфом [10], может существовать миграция ацетатных групп от γ – к α-атому.
Простые эфирные связи между фенилпропановыми единицами. Область сигналов атомов боковых цепочек на HSQC-спектре (δН/δС 2,55,5/50-90 м.д.), представленная на рис. 4 и в табл. 2, дает информацию о связях между звеньями макромолекулы лигнина. Также в этой области спектра находятся сигналы метоксильных групп при δН/δС 3.7/55.7 м.д., которые характерны для всех известных лигнинов. Содержание метоксильных групп в разных лигнинах варьирует от 15 до 20 % в зависи- мости от соотношения гваяцильных, сирингильных и п-кумаровых структурных единиц [22]. Лигнин серпухи венценосной содержит еще и этоксильные группы. Это предположение основано на корреляционных сигналах СН3-фрагмента (δН/δС 1.10/15.0 м.д.) и СН2 (δН/δС 3.4/62.5 м.д. см. рис. 4 а). Подтверждением является кросс-сигнал на COSY спектре δН/δН 1.10/3.4 м.д.
Характерными для структуры β-арилового эфира (рис. 5 А) корреляционными сигналами, в соответствии с [9], являются δ Н /δ С 3.5/60.1 м.д. (рис. 4 Aγ) и 4.7/71.1 м.д. (рис. 4 Аα). Необходимо отметить, что сигнал β-атома гваяцильной или сирин-гильной единицы, соединенной с гваяцильной, имеет корреляцию при δ Н /δ С 4.4/84.0 м.д (рис. 4 Aβ S/G-O-G ) [9]. Этот корреляционный сигнал обычно четко разделен с корреляционными сигналами δ Н /δ С 4.0/87.7 м.д. β-атома сирингильных единиц,
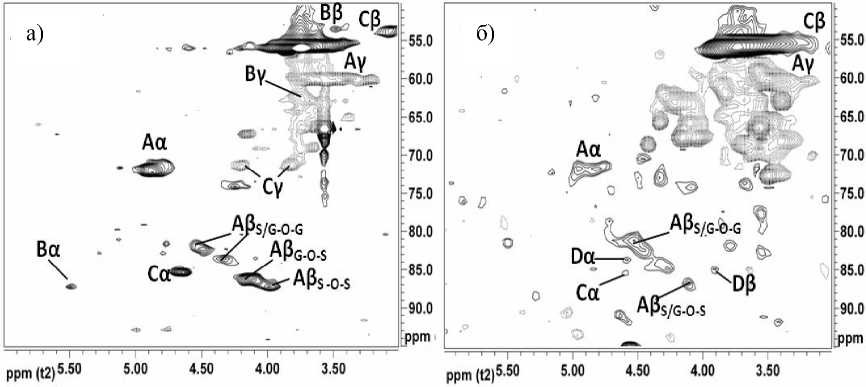
Рис. 4. Области HSQC-спектров лигнинов серпухи венценосной (а) и родиолы розовой (б).
Таблица 2
Отнесение кросс-сигналов 1Н-13С в HSQC-спектрах лигнинов ДЛС и ДЛР [8, 9]
Алифатические фрагменты. В диапазоне δ Н /δ С 3.0-1.1/50-10 м.д. на HSQC-спектре (рис. 6 нижний) присутствуют сигналы алифатических углеводородных фрагментов, не связанных с атомом кислорода, в основном групп СН 2 . Эти сигналы не
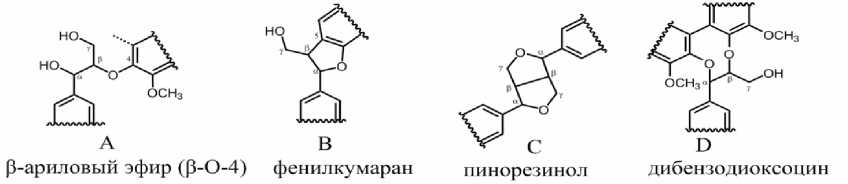
Рис. 5. Структурные фрагменты лигнинов.
имеющих в β-положении аналогичную связь с сирингильными (рис. 4 Aβ S-O-S ), а также со структурами с сигналом при δ Н /δ С 4.2/86.3 м.д. (рис. 4 Aβ G-O-S ) [9]. Структуры β-арилового эфира считаются преобладающими во всех лигнинах [2], и спектры лигнинов родиолы и серпухи (рис. 4 а и б) демонстрируют четкие и интенсивные характерные сигналы.
Фенилкумарановые фрагменты (рис. 5 В) распространены в гваяцилсирингильных лигнинах (лигнины лиственных) реже, чем в гваяцильных (лигнины хвойных), где их количество составляет 810 на 100 фенилпропановых единиц [11]. Наличие большого количества сирингильных звеньев, с метоксилированным пятым поло-
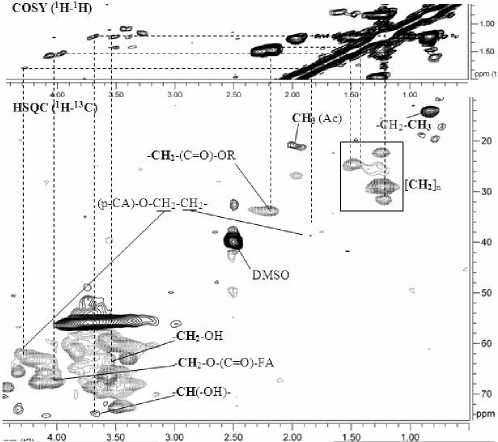
Рис. 6. COSY- и HSQC-спектры ДЛР.
принадлежат боковым цепочкам фенилпропановой единицы, они характерны для атомов углерода насыщенных ациклических структур с длиной цепи 7 и более атомов углерода [11]. Структурообразующими компонентами клеточных стенок растений, построенными из остатков жирных кислот и высших спиртов, являются кутин и суберин [23]. Эти компоненты обычно извлекаются из растения экстракцией нейтральными растворителями в ходе предварительной подготовки сырья для извлечения лигнина, поэтому присутствие жирных кислот и высших спиртов в препарате лигнина может быть либо в виде примеси, либо в виде фрагментов структуры, ковалентно связанных с фенилпропановыми единицами.
В работе [23] по исследованию состава жирных кислот корневищ ряда растений было выявлено, что для родиолы ( Rhodiola pamiroalaica ), как и для остальных, свойственно преобладание жирных кислот с цепочками С 16 и С 18 . На основе данной работы проведено соотнесение сигналов в спектрах лигнинов с фрагментами на примере фрагмента FA (от англ. fatty acids), представленного на рис. 7. Отнесение сигналов показано в табл. 3.
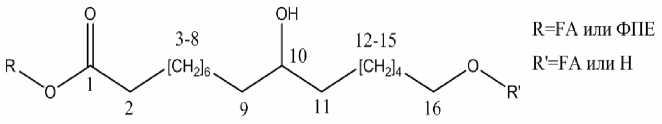
Рис. 7. Фрагмент 10, 16-дигидроксигексадекановой кислоты в составе лигнина (усл. обозн. – FA).
Таблица 3
Отнесение сигналов 1Н и 13С жирных кислот [23]
|
Положение |
Фрагмент |
1 13 С δ, м.д. |
1 Н δ, м.д. |
|
1 |
RO-( C =O)-CH 2 - |
172-175 |
- |
|
2 |
RO-(C=O)- CH 2 - |
34.08 |
2.20 |
|
3-8, 12-15 |
- CH 2 - |
25-32 |
1.2-1.5 |
|
9, 11 |
- CH 2 -CH(OH)- |
37.40 |
1.24 |
|
10 |
- CH (-OH)- |
73.85 |
3.69 |
|
16 |
- CH 2 -OH |
62.80 |
3.63 |
Возможно, что концевые спиртовые группы FA (положение 16 на рис. 7) этерифицированны п -кумаровой кислотой (δ Н /δ Н 4.30/1.80 м.д.; δ Н /δ С 4.30/62.0 м.д.) и аналогичными фрагментами FA (δ Н /δ Н 4.05/1.50 м.д.; δ Н /δ С 4.05/67.5 м.д.). Дополнительным подтверждением является сигнал δ Н /δ С 4.30/167.4 на HMBC-спектре образца ДЛР (рис. 8), регистрирующий дальние протон-углеродные взаимодействия четвертичного углерода п -кумара-тового фрагмента с протонами в 16-й позиции FA (рис. 7). Также на этом HMBC-спектре выявлены корреляционные сигналы, подтверждающие наличие сложноэфирных связей между цепочками FA (δ Н /δ С 4.05/172.5 м.д.). Тип связи фрагментов FA с фенилпропановыми звеньями лигнина родиолы розовой по полученным спектрам, установить не удалось. Причинами могут служить как наложение сигналов, так и сравнительно низкая распространенность связей между FA и лигнином. Однако высока вероятность того, что FA являются фрагментами индивидуального компонента клеточной стенки, оказавшегося связанным с лигнином в ходе выделения препарата, поскольку даже процедура переосаждения лигнина из диоксана в диэтиловый эфир не привела к отделению этого компонента.
Особенностью лигнина серпухи венценосной является наличие в его структуре этоксильных групп. Это предположение основано на корреляционных сигналах на HSQC-спектре ДЛС: δ Н /δ С 1.10/15.0 м.д. (СН 3 ; эта область спектра не представлена), δ Н /δ С 3.4/62.5 м.д. (СН 2 ; рис. 4 а), COSY(не представлен): δ Н /δ Н 1.1/3.4 м.д. Согласно [9], химический сдвиг протонов СН 2 -фрагмента этоксильной группы 3.4 м.д. характерен для этоксильной группы в алифатической цепочке фенилпропановых единиц.
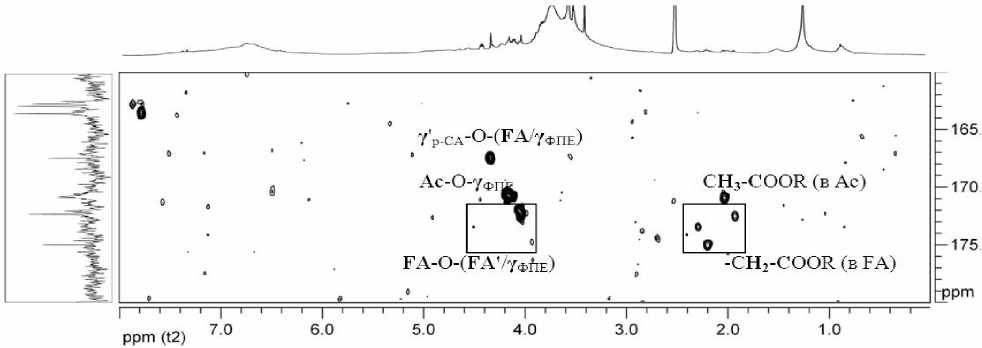
Рис. 8. HMBC-спектр ДЛР область корреляции сигналов атомов сложноэфирных связей.
Заключение
Лигнины, образовавшись в растительной ткани, выполняют разнообразные функции. В зависимости от роли растительной ткани, типа растения и его филогенетического происхождения, эти вещества могут различаться по химическому составу и структуре, но неизменно одно – все они построены из фенилпропановых структурных звеньев, соединенных простыми эфирными и углерод-углеродными связями [24]. Весьма необычно, по сравнению с известными на сегодняшний день лигнинами, выглядит лигнин, выделенный из родиолы розовой. В этом образце не выявлены фенилкумарановые фрагменты, считающиеся характерными для всех лигнинов. В составе этого образца значительную долю занимают длинные насыщенные алифатические цепочки. Предшественниками этого лигнина, помимо трех классических монолигнолов, могут быть кумара-ты кониферилового и синапового спиртов и, возможно, ацетаты этих спиртов. Лигнин серпухи венценосной не содержит сложноэфирных связей и структур дибензодиоксоцина, но его особенность в том, что в его структуре имеются этоксильные группы в боковой цепочке фенилпропановых единиц.
Работа выполнена в рамках Программы Президиума РАН «Создание и совершенствование методов химического анализа и исследования структуры веществ и материалов» (проект № 12-П-3-1024).
Список литературы Изучение химической структуры лигнинов родиолы розовой (Rhodiola rosea L.) и серпухи венценосной (Serratula coronata L.) методом 2D ЯМР-спектроскопии
- Лигнины (структура, свойства и реакции)/Под ред. К. Сарканена, К. Людвига. М.: Лесная пром-сть, 1975. 632 с.
- Карманов А.П. Самоорганизация и структурная организация лигнина. Екатеринбург: УрО РАН, 2004. 270 с.
- Wiegant F., Surinova S., Ytsma E. et al. Plant adaptogens increase lifespan and stress resistance in C. Elegans//Biogerontology. 2009. Vol. 10. № 1. P. 27-42.
- Уфимцев К.Г., Ширшова Т.И., Якимчук А.П., Володин В.В. Гормональное, токсическое и адаптогенное влияние экдистероидов Serratula coronata L. на личинок Ephestia Kiihniella. Zell.//Растительные ресурсы. 2002. Т. 38. №2. С. 29-39.
- Avula B., Wang Y. H., Ali Z., Smillie T. J. et al. RP-HPLC determination of phenylalkanoids and monoterpenoids in Rhodiola rosea and identification by LC-ESI-TOF//Biomed Chromatogr. 2009. Vol.23. № 8. P. 865-872.
- Ангаскиева А.С., Андреева В.Ю., Калинкина Г.И. и др. Исследование химического состава серпухи венценосной, культивируемой в Сибири//Химия растительного сырья. 2003. № 4. С. 47-50.
- Оболенская А.В., Ельницкая З.П., Леонович А.А. Лабораторные работы по химии древесины и целлюлозы. М.: Экология, 1991. 256 с.
- Ralph J., Marita J. M., Ralph S. A. et al. Solutionstate NMR of lignin//AdVances in Lignocellulosics Characterization. 1999. P. 55-108.
- Ralph S.A., Ralph J., Landucci L. NMR database of lignin and cell wall model compounds. Madison, WI, U.S.: Forest Products Laboratory, 2004. (http://ars.usda.gov/Services/docs.htm?docid). 498 p.
- Ralph J. An unusual lignin from kenaf//J. Nat. Prod. 1996. № 59. P. 341-342.
- Калабин Г.А., Каницкая Л.В., Кушнарев Д.Ф. Количественная спектроскопия ЯМР природного органического сырья и продуктов их переработки. М.: Химия, 2000. 408 с.
- Smith D. C. C. Ester groups in lignin//Nature. 1955. Vol. 176. P. 267-268.
- Nakamura Y., Higuchi T. Ester linkage of pcoumaric acid in bamboo lignin. III. Dehydrogenative polymerization of coniferyl p-hydroxybenzoate and coniferyl p-coumarate//Cellul. Chem. Technol. 1978. Vol. 12. P. 209-221.
- Ralph J., Helm, R. F. Lignin/hydroxycinnamic acid/polysaccharide complexes: Synthetic models for regiochemical characterization. In Forage Cell Wall Structure and Digestibility, International Symposium. Madison, WI: ASACSSA-SSSA, 1993. P. 201-246.
- Ralph J., Hatfield R. D., Quideau S. et al. Pathway of p-coumaric acid incorporation into maize lignin as revealed by NMR//J. Am. Chem. Soc. 1994. Vol. 116. Р. 9448-9456.
- Crestini C., Argyropoulos D. S. Structural analysis of wheat straw lignin by quantitative 31P and 2D NMR spectroscopy. The occurrence of ester bonds and R-O-4 substructures//J. Agric. Food Chem. 1997. Vol. 45. Р. 1212-1219.
- Rio J., Rencoret J., Marques G. et al. Highly acylated (acetylated and/or p-coumaroylated) native lignins from diverse herbaceous plants//J. Agric. Food Chem. 2008. Vol. 56. No. 20. P. 9525-9534.
- Ralph J., Lundquist K., Brunow G. et al. Lignins: natural polymers from oxidative coupling of 4-hydroxyphenylpropanoids//Phytochem. 2004. No 3. P. 29-60.
- Landucci L. L., Deka G. C., Roy D. N. A. 13C NMR study of milled wood lignins from hybrid Salix clones//Holzforschung. 1992. Vol. 46. Р. 505-511.
- Sun R. C., Fang J. M., Goodwin A. et al. Fractionation and characterization of ball-milled and enzyme lignins from abaca fibre//J. Sci. Food Agric. 1999. Vol. 79. Р. 1091-1098.
- Lu F., Ralph J. Detection and determination of p-coumaraloylated units in lignin//J. Agric. Food Chem. 1999. Vol. 47. Р. 1985-1992.
- Далимова Г.Н., Абдуазимов Х.А. Лигнины травянистых растений//Химия природных соединений. 1994. № 2. С. 160-177.
- Tsydendambaev V.D., Christie W.W., Brechany E.Y., Vereshchagin A.G. Identification of unusual fatty acids of four alpine plant species from the Pamirs//Phytochemistry. 2004. Vol. 65. P. 2695-2703.
- Резников В.М., Михасева М.Ф. О филогении лигнина//Химия древесины. 1982. № 6. С. 77-87.
