Изучение хирономид (Diptera, Chironomidae) в низовьях Волги и в Северном Каспии и их многолетнее изменение биомассы
 в низовьях Волги и в Северном Каспии и их многолетнее изменение биомассы")
Автор: Зинченко Татьяна Дмитриевна, Малиновская Любовь Васильевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 3-1 т.15, 2013 года.
Бесплатный доступ
Представлены результаты таксономического состава хирономид в водотоках Низовьев Волги и в Северной части Каспийского моря в различные периоды 1981-1995 гг. Показана многолетняя динамика биомассы хирономид в Северном Каспии за период с 1948 по 2000 г. Данные рекогносцировочных таксономических исследований свидетельствуют об отсутствии современного представления о составе хирономидофауны региона.
Хирономиды (chironomidae), таксономический состав, chironomus albidus konst. многолетняя динамика биомассы, водотоки дельты и авандельты волги, северный каспий
Короткий адрес: https://sciup.org/148201807
IDR: 148201807 | УДК: 595.771
A study of the Chironomidae (Diptera) of the Lower Volga river region and the Northern Caspian sea and their long-term biomass’ dynamics
There are given the results of taxonomical structure of chironomids in waterways of The Lower Volga river region and the Northern Caspian Sea during various periods of 1981-1995. Long-term dynamics of the chironomids' biomass in the Northern Caspian Sea from 1948-2000 is shown. Data of reconnaissance researches prove the lack of modern representation of the structure of chironomid fauna in natural habitats. Extensine use of Chironomidae in monitoring of The Lower Volga river region and the Northern Caspian Sea is consistent with the abundance and taxa richness of this group in natural habitats.
Текст научной статьи Изучение хирономид (Diptera, Chironomidae) в низовьях Волги и в Северном Каспии и их многолетнее изменение биомассы
И сторически сложившиеся повышения и понижения уровня Каспия, контролируемые динамикой глобальных гидроклиматических процессов приводят к перераспределению местообитаний гидробионтов разной толерантности по отношению к различным факторам воздействия в том числе и в результате трансгрессии Каспийского моря [38, 45] . Происходят динамичные преобразования гидролого-гидрохимических факторов, биотопической структуры, обусловливая изменения видового состава, плотности и биомассы планктонных и донных сообществ, что характерно для различных районов Каспийского моря и в первую очередь для Северной части Каспия. Этот район моря можно рассматривать как классическую модель для типизации природнопреобразовательных процессов. На этом основании Северный Каспий представляет собой уникальный полигон для исследований локальных и глобальных процессов динамики численности донных сообществ, изучение которых в многолетнем аспекте дает возможность показать динамику трофических условий и прогнозирования состояния промысловых рыб Каспия.
Период падения уровня Каспийского моря (1930-1977 гг.) сменился его подъёмом, который начался с 1978 г. Абсолютная отметка уровня Каспия достигла – 27,03 м. к 2002 году. Трансгрессия моря сказалась в первую очередь на экосистеме Северного Каспия, что явилось причиной значительного увеличения объема воды, возрастания, за счет затопления новых территорий, площади этой части моря, понижения солености, изменения гидрологического режима и степени
накопления и распределения осадков [21, 22, 23]. Указанные факторы в различной степени оказали влияние на процессы структурных преобразований донных сообществ, отдельных беспозвоночных, составляющих ядро кормовой базы рыб-бентофагов [27, 28, 29, 35, 36, 37, 40, 56 и др.]. Вышесказанное позволяет констатировать, что анализ многолетних изменений количественного развития ценозов макрозообентоса, имеющих значение биоиндикаторов в современный период трансгрессии экосистемы Каспия, является актуальным и практически значимым.
Гидрологические и геоморфологические изменения, вызванные колебанием уровня Каспийского моря, обусловили преобразования экосистемы водоемов авандельты и нижней зоны дельты р. Волги. Напомним, что в первой трети прошлого века при понижении уровня моря, уменьшении естественного стока Волги, а в дальнейшем - при продолжении снижения стока реки Волга в связи с ростом водопотребления, а также его регулированием плотинами водохранилищ, образовались обширные мелководья, осушные острова и произошло зарастание авандельты макрофитами. Амплитуда колебания уровня моря составила 3.13 м, достигая минимальной отметки (-29.02 м абс.) в 1977 г. [26]. С 1978 г. наблюдалось устойчивое повышение уровня Каспия (в 1986 г.- минус 27,8 м абс.), обусловленное рядом причин, среди которых общие изменения климата и интенсивность баланса «сток-испарение» вызывают колебания уровня моря [24, 41]. По данным Гидрометцентра РФ средний уровень моря в 1994 г достиг отметки -26,65 м абс., превысив средний уровень 1977 г. на 228 см. Трансгрессия моря сказалась в первую очередь на экосистеме Северного Каспия, что явилось причиной значительного увеличения объема воды, возрастания, за счет затопления новых территорий, площади этой части моря, понижения солености, изменения гидрологического режима и степени накопления осадков [21, 52].
По мере заиления островной зоны авандельты, начиная с 1988 г. и в последующие годы, была выявлена тенденция на увеличение плотности поселений малощетинковых червей и хирономид, когда численность олигохет достигла 3160 экз./м 2 , а хирономид - 3680 экз./м 2 . При продолжающемся повышении уровня Каспия регистрировалось увеличение численности пелофильной фауны [50].
Сведения о современной фауне хирономид дельты Волги и Каспия приобретают особую актуальность в связи с фрагментарностью исследований этого уникального региона [16]. Недавно опубликованные данные [1] о том, что «бентос Северного Каспия не отличается большим разнообразием видов», со ссылкой на В.П. Иванова и А.Ф. Сокольского (2000), по меньшей мере не отражают действительное фаунистическое состояние биоты Каспия вообще и его Северной части, в частности. Достаточно сказать, что ранее констатировалось более 400 видов и таксонов надвидового ранга в Каспийском море [9], а современные фаунистические сведения состояния макрозообентоса вообще и хирономид в частности, обитающих в дельтовых районах Волги и в Северном Каспии, отсутствуют. М.Г. Карпинским в 1986-1987 гг. по результатам съемок в августе, октябре и феврале было идентифицировано 120 видов донных беспозвоночных в Среднем и Южном Каспии [18]. Указывается на то, что бенто-фауна Каспийского моря в настоящее время насчитывает 379 видов животных, из которых 140 видов ракообразных, 55 видов червей и 116 видов моллюсков [12]. Однако, целые классы гидробионтов, например, такие как Oligochaeta, Nematoda, семейство двукрылах Chironomidae до настоящего времени в фаунистическом отношении остаются практически не изученными. Вместе с тем известно, что по предварительным данным, для России известно более 900 валидных видов хиро-номид [25]. По сведениям А.И.Шиловой и Н.И. Зеленцова [53], в водоемах Верхней Волги хиро-номиды представлены 234 видами. Нами для водоемов и водотоков бассейна Средней и Нижней Волги указывается 234 вида и личиночные формы хирономид [15]. Ранее достаточно интенсивно изучавшиеся хирономиды дельты Волги и аван-дельты Каспия [5,б,7,8,16] в настоящее время не находят своего исследователя.
Что касается пищевой ценности личинок хи-рономид, то в литературе приводятся данные, говорящие о том, что относительная доля мотыля в бентосе Северного Каспия составляет до 1.5%, тогда как доля его в кишечниках осетровых может достигать, например, в районе о. Тюлений в северном Каспии более 6% [33]. Известно, что в условиях предустьевого пространства Волги сазан потребляет в основном гаммарид (18,5%) и корофиид (15%). Но наряду с ракообразными он в большом количестве использует в пищу моллю- сков (18%) и хирономид (16%). При оценке суточных рационов бычковых, было установлено, что популяция некоторых видов выедает 3% общей продукции бентоса и 10% валового запаса ракообразных и хирономид [47].
Необходимость изучения хирономид, помимо вышесказанного, подтверждается их высокой биоиндикационной значимостью в оценке экологического состояния водоемов и водотоков Палеарктики [11, 13, 14, 15, 55].
В статье представлены материалы собственных исследований бентоса низовьев Волги и Северного Каспия (1981-1985, 1993, 1995 гг.), где акцент сделан на изучение таксономического состава хирономид и динамики распределения личинок в различных водотоках дельты и авандель-ты, а также в Северной части Каспийского моря на протяжении длительного периода исследований. Результаты анализа многолетней динамики сообществ зообентоса Северного Каспия в период подъем уровня моря представлены нами ранее [30].
МАТЕРИАЛЫ И МЕТОДЫ
Личинки хирономид из собранных образцов бентоса в акватории Северного Каспия в 19811984 гг. были любезно предоставлены В.Ф. Осадчих (КаспНИРХ), для определения таксономического состава. В култучной зоне (1981-1985 гг., 1993 г.), в ериках и протоках (1983, 1985, 1993, 1995 гг.), ильменях окрестностей г. Астрахани (1993 г.), в районе Дамчикского участка Астраханского государственного Биосферного заповедника и на станциях Гандуринского канала, а также в протоках Бузан, Царев, Быстрая (1993, 1995 гг.) и каналах в черте г. Астрахань (1995 г.) отбор образцов бентоса осуществлялся в период рекогносцировочных исследований при проведении хоздоговорных исследований, студенческих практик и плановых научных исследований на кафедре гидробиологии и ихтиологии Астраханского технического института рыбной промышленности и хозяйства (ныне АГТУ). Отбор проб бентоса производили дночерпателем Петерсена (1/40 м 2 , 2-х кратная повторность) или пробоотборниками, а также гидробиологическим скребком. Отобранные образцы грунта промывали через капроновый газ № 23 (размер ячеи 300-310 мкм) с последующей разборкой гидробионтов под бинокулярным микроскопом.
В Северном Каспии пробы бентоса отбирали дночерпателем «Океан-50», с площадью захвата 0,1 м². В зарослях макрофитов (култучная зона Северного Каспия) во время экспедиционного рейса сборы осуществлялись при помощи рамки Зимбалевской (0.5 м). Содержимое дночерпателя и пробоотборников промывали через капроновое сито из газа № 14. Образцы проб макрозообентоса фиксировали 4% - ным раствором формальдегида.
Обработку материала, собранного в Северном Каспии производили в лаборатории КаспНИРХа по методике [42]. За период исследований собрано и обработано более 200 образцов бентоса и хирономид. Для более полной таксономической сводки представлены данные сборов хирономид М.С. Алексевниной в дельте р. Волги [5].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Таксономический состав хирономид различных водоемов и водотоков дельты Волги и Северного Каспия включает 103 вида и таксона надвидового ранга. Наибольшим количеством таксонов (78 видов и личиночных форм) представлены водоемы дельты, ранее изученные М.С. Алексев-ниной, среди которых представители рода Chi-ronomus , при отсутствии возможности проведения цитологических исследований, не были идентифицированы до видового уровня. Начиная с 1981 года в различных водотоках и водоемах Низовьев Волги, при проведении рекогносцировочных исследований, было зарегистрировано 25-33 видов и таксонов (в ериках, водотоках култучной зоны и дельтовых каналах района заповедников); не более 7-11 таксонов было найдено в каналах в черте г. Астрахань, ильменях, реках Царев и Бу-зан. К представителям подсемейства Chironomi-nae относятся 66 таксонов, включая таксоны трибы Tanytarsini . К представителям Orthocladiinae относится 24 вида и личиночной формы, к Tanypodinae - 10, к Diamesinae – 2. В отдельных районах Северного Каспия в различные периоды отбора проб было зарегистрировано 17 видов и представителей надвидового ранга хирономид.
Отметим, что, по результатам сборов бентоса, полученных нами в экспедиционных исследованиях, а также на основании литературных сведений, можно констатировать, что в результате подъема уровня моря в 80-х и начале 90-х годов в бентофауне водоемов островной зоны авандельты наметился процесс снижения доли пелофильной фауны при возрастании пелореофильной и рео-фильной. Глубины на дельтовых и авандельтовых участках возросли, что на фоне увеличения стока Волги привело к их проточности и уменьшению степени заиления [4]. В этот период среди хиро-номид из 17 отмеченных личиночных форм в островной зоне Северного Каспия, наряду с псаммо-пелофильными таксонами Chironomus plumosus, Fleuria lacustris, Polypedilum nubeculo-sum , стали встречаться личинки Cladotanytarsus mancus , Endochironomus albipennis , Glyptotendipes viridis , G.gripekoveni , Polypedilum sordens , Psectro-cladius sp., P. gr. dilatatus, Clinotanypus nervosus , Trissocladius sp. и другие эврибионтные виды, среди которых преобладали представители фито-фильной фауны.
Число видов хирономид увеличилось в аван-дельте от 55 в 1967-1970 гг. до 62 в 1983-1984 гг.
Численность личинок оставалась на уровне 19671970 гг., тогда как среднесезонная биомасса возросла и составила 4,76 г/м 2 . В отдельных ериках и протоках (ер. Дарма, проток Дамчик) биомасса личинок Chironomus plumosus в сентябре 1983 г. на отдельных заросших и заиленных участках достигала 300 г/м 2 . В дальнейшем, в 1991-1992 гг. при повышении водности Волги, подъеме уровня моря и увеличении его акватории наблюдается устойчивая тенденция к увеличению числа видов пресноводного комплекса [34].
В донной фауне островной зоны авандельты было зарегистрировано увеличение численности хирономид пелофильного комплекса, среднесезонная биомасса которых также была высокой -4,81 г/м 2 [51]. Видовой состав хирономид в период 1993-1995 гг. представлен устойчивым комплексом пело- и фитофильных (из фауны зарослей) эврибионтных видов (табл. 1). Помимо хи-рономид, в многолетней сезонной и пространственной динамике макрозообентоса дельтовых водотоков установлены изменения в структуре донных сообществ, связанные с увеличением численности олигохет в 2002-2003 гг. в 2-4,5 раза по сравнению с предшествующими годами [50,51], что наряду с данными гидрохимического анализа, свидетельствует об эвтрофировании водотоков низовьев Волги.
Процесс эвтрофирования водотоков дельты р. Волги, Северного Каспия и северной части Среднего Каспия в последние 15 лет существенно возрос [22, 43, 44].
Повышение уровня моря и его распреснение в отдельные годы внесли изменения в качественный состав и количественные показатели гидробионтов. При оценке динамики общего бентоса, изложенного в отдельных публикациях, хироно-мидам также уделяется мало внимания [4, 49]. Напомним, что хирономиды Каспийского моря представляют собой группу двукрылых, проникших, вероятно из пресноводных притоков и затем приспособившихся в медленно протекавшему процессу осолонения [3]. Анализ имеющихся в нашем распоряжении фактических данных таксономического состояния хирономид Северного Каспия в предоставленных В.Ф. Осадчих образцах бентоса различных районов Северного Каспия свидетельствует о том, что в 1981-1984 гг. на мелководных участках Северного Каспия было зарегистрировано 8 таксонов личинок хирономид, составляющих пресноводный ценоз в сообществах бентоса: Cryptochironomus gr. defectus , Chi-ronomus albidus Konst., Dicronendipes gr. tritomus , Glyptotendipes gripekoveni Kieff., Polypedilum con-victum (Walker), Polypedilum nubeculosum (Meig), Paratanytarsus confusus Pal., Cladopelma gr. lateralis ( Goetgh.). При дальнейших исследованиях бентоса островной зоны Северного Каспия в составе экспедиции КаспНИРХа (1993, 1995 гг.)
были отмечены дополнительно к названным Ana-topynia plumipes, Clinotanypus nervosus, Procladius choreus, Procladius ferrugineus, Psectrocladius gr. dilatatus, Dicrotendipes gr. nervosus, Endochirono-mus albipennis, Glyptotendipes viridis , Chironomus albidus. За исключением C. albidus , указанные хирономиды относятся к пресноводному и слабосолоноватоводному ценозу, распространены, в основном, в опресненных районах западного побережья Северного Каспия. Преобладют представители солоноватоводного комплекса, личинки C. albidus, которые в Северном Каспии составляют около 90% от общей плотности хирономид. Их биомасса в продуктивных районах Главного банка и Жесткого Осередка локально, на глубине 3,6
м может достигать 3,5-12,1 г/м 2 , при численности 700-1237 экз./м 2 . В Среднем Каспии личинки С. albidus были зарегистрированы нами на максимальной глубине в 135 м [16]. Интересно отметить, что в Среднем и Южном Каспии личинки хирономид и олигохеты, хорошо осваивая глубоководные районы, при отсутствии другой фауны, являются единственными представителями бентоса на глубинах, превышающих 100 м [18]. В настоящее время С. albidus известен в основном из Каспийского моря, хотя единичные личинки нами были найдены в 1991 г. в Волгоградском водохранилище, в реке Бузан в 1995 г. и отнесены к чужеродным видам, представителям инвазионного комплекса.
Таблица 1. Таксономический состав личинок хирономид в различных водотоках дельты Волги и в Северном Каспии
|
Хирономиды |
s ^ m ^l s s 5^ 4 |
s3 m § н G ^ ^ CO H CT' 00 |
§§ to mr |
s У c3 1 5 5 § | 4 |
g-a СЦ |
-Q Й У CQ 5 s |
G S Ф S o> )S g tH 3°s |
||
|
Ё |
S3 s- ® Ф |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
Подсем.Tanypodinae |
|||||||||
|
Ablabesmyia longistyla Fitt. |
+ |
||||||||
|
Ablabesmyia monilis (L.) |
+ |
||||||||
|
Ablabesmyia phatta Egg. |
+ |
||||||||
|
Anatopynia plumipes Fries |
+ |
+ |
+ |
||||||
|
Clinotanypus nervosus (Mg.) |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
Procladius choreus (Mg.) |
+ |
+ |
+ |
+ |
+ |
+ |
|||
|
Procladius ferrugineus (Kieff.) |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
Psectrotanypus varius (Fabr.) |
+ |
||||||||
|
Tanypus punctipennis (Mg.) |
+ |
+ |
+ |
+ |
+ |
||||
|
Tanypus vilipennis (Kieff.) |
+ |
+ |
+ |
+ |
|||||
|
Подсем. Diamesinae |
|||||||||
|
Syndiamesa sp. |
+ |
+ |
|||||||
|
Подсем. Orthocladiinae |
|||||||||
|
Acricotopus lucens (Zett.) |
+ |
+ |
|||||||
|
Corynoneura celeripes (Winn.) |
+ |
+ |
+ |
||||||
|
Corynoneura scutellata Winn. |
+ |
+ |
|||||||
|
Cricotopus algarum Kieff. |
+ |
+ |
+ |
||||||
|
Cricotopus gr . algarum |
+ |
||||||||
|
Cricotopus bicinctus (Mg.) |
+ |
+ |
+ |
||||||
|
Cricotopus sylvestris Fabr. |
+ |
+ |
+ |
+ |
+ |
+ |
|||
|
Cricotopus tibialis (Meig.) |
+ |
||||||||
|
Cricotopus latidentatus Tsh. |
+ |
||||||||
|
Eukiefferiella alpestris Goetgh. |
+ |
||||||||
|
Eukiefferiella sp. |
+ |
||||||||
|
Limnophyes pusillus Eaton = Limnophyes sp. |
+ |
+ |
+ |
||||||
|
Limnophyes transcaucasicus Tsch. |
+ |
||||||||
|
Nanocladius bicolor (Zett.) |
+ |
+ |
|||||||
|
Orthocladius oblidens (Walk.) |
+ |
+ |
|||||||
|
Propsilocerus orielicus Tsch. |
+ |
||||||||
Продолжение таблицы 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
Psectrocladius gr. dilatatus |
+ |
+ |
+ |
||||||
|
Psectrocladius barbimanus Edw. |
+ |
||||||||
|
Psectrocladius gr. рsilopterus |
+ |
+ |
+ |
+ |
|||||
|
Psectrocladius simulans Joh. |
+ |
||||||||
|
Trissocladius potamophilus Tsch. |
+ |
+ |
|||||||
|
Thienemanniella clavicornis Tsch. |
+ |
||||||||
|
Thienemanniella flaviforceps Tsch. |
+ |
||||||||
|
Thienemanniella sp. |
+ |
||||||||
|
Подсем. Chironominae |
|||||||||
|
Триба Chironomini |
|||||||||
|
Camtochironomus tentans (F.) |
+ |
+ |
+ |
||||||
|
Camtochironomus pallidivitatus Mall. |
+ |
||||||||
|
Chironomus albidus Kons. |
+ |
+ |
|||||||
|
Chironomus sp. 1 |
+ |
||||||||
|
Chironomus sp. 2 |
+ |
||||||||
|
Chironomus sp. 3 |
+ |
||||||||
|
Chironomus sp. 4 |
+ |
||||||||
|
Chironomus cingulatus |
+ |
+ |
+ |
||||||
|
Chironomus dorsalis Mg. |
+ |
||||||||
|
Chironomus heterodentatus Konst. |
+ |
||||||||
|
Chironomus gr. plumosus |
+ |
+ |
+ |
+ |
+ |
+ |
|||
|
Chironomus gr. tummi |
+ |
+ |
+ |
+ |
|||||
|
Cladopelma gr . lateralis |
+ |
||||||||
|
Cryptocladopelma fridmanae Tsch. |
+ |
||||||||
|
Cryptocladopelma gr. laccophila |
+ |
||||||||
|
Cryptochironomus gr. defectus |
+ |
+ |
+ |
+ |
+ |
+ |
|||
|
Cryptochironomus supplicans Mg. |
+ |
+ |
|||||||
|
Cryptochironomus obreptans (Walk.) |
+ |
||||||||
|
Cryptochironomus ussourennsis Goetgh. . |
+ |
+ |
+ |
||||||
|
Cryptochironomus gr. fuscimanus |
+ |
+ |
|||||||
|
Cryptochironomus gr. conjugens |
+ |
+ |
+ |
||||||
|
Cryptochironomus burganadzeae Tshern. |
+ |
||||||||
|
Cryptochironomus gr. camptolabis |
+ |
||||||||
|
Cryptochironomus nigronitens (Edw.) |
+ |
+ |
|||||||
|
Cryptochironomus sp. (№ 9 Lipina) |
+ |
||||||||
|
Demicryptochironomus vulneratus (Zett) |
+ |
||||||||
|
Dicrotendipes nervosus (Staeg.) |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Dicrotendipes gr. lobiger |
+ |
||||||||
|
Dicrotendipes pulsus objectans Walk. |
+ |
||||||||
|
Dicrotendipes gr. tritomus |
+ |
||||||||
|
Paratendipes transcaucasicus Tsch. |
+ |
||||||||
|
Endochironomus albipennis (Mg.) |
+ |
+ |
|||||||
|
Endochironomus gr. dispar |
+ |
||||||||
|
Endochironomus tendens (Fabr.) |
+ |
+ |
|||||||
|
Endochironomus impar (Walk.) |
+ |
+ |
|||||||
|
Fleuria lacustris Kieff. |
+ |
+ |
+ |
||||||
|
Glyptotendipes glaucus (Mg.) |
+ |
||||||||
|
Glyptotendipes gripekoveni (Kieff.) |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
||
|
Glyptotendipes barbipes (Staeg.) |
+ |
||||||||
|
Glyptotendipes paripes Edw. |
+ |
||||||||
|
Glyptotendipes sp. |
+ |
||||||||
|
Glyptotendipes viridis (Mac.) |
+ |
+ |
+ |
+ |
+ |
||||
|
Harnischia virescens Mg. |
+ |
+ |
|||||||
|
Lauteborniella agrayloides (Kieff.) |
+ |
||||||||
|
Lipiniella araenicola Shilova |
+ |
||||||||
|
Microtendipes pedellus (De Geer) |
+ |
||||||||
|
Parachironomus arcuatus (Goetgh.) |
+ |
+ |
|
Окончание таблицы 1 |
|||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
7 |
8 |
10 |
|
Parachironomus varus Goetgh. |
+ |
||||||||
|
Parachironomus vitiosus Goetgh. |
+ |
||||||||
|
Paralauterborniella nigrohalter-alis (Mall.) |
+ |
+ |
|||||||
|
Pentapedilum exsectum Kieff. |
+ |
+ |
+ |
||||||
|
Polypedilum bicrenatum Kieff. |
+ |
+ |
+ |
||||||
|
Polypedilum breviantennatum Tsh. = P. сonvictum (Walk.) |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Polypedilum scalaenum (Schrank) |
+ |
+ |
+ |
||||||
|
Polypedilum nubeculosum (Mg.) |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Polypedilum pedestre Mg. |
+ |
||||||||
|
Polypedilum sordens (v.d.Wulp) |
+ |
+ |
|||||||
|
Tendipedini macrophtalma Tsh. |
+ |
+ |
|||||||
|
Xenochironomus xenolabis Kieff. |
+ |
||||||||
|
Триба Tanytarsini |
|||||||||
|
Cladotanytarsus gr. mancus |
+ |
+ |
+ |
+ |
+ |
+ |
|||
|
Micropsectra gr. praecox |
+ |
||||||||
|
Paratanytarsus confusus Palm. |
+ |
+ |
+ |
+ |
+ |
||||
|
Stempellina bausei (Kieff.) |
+ |
+ |
+ |
||||||
|
Tanytarsus gr. g regarius =T. sp. |
+ |
+ |
+ |
+ |
|||||
|
Tanytarsus gr. lauterborni = Paratanytarsus gr. lauterborni |
+ |
+ |
|||||||
|
Tanytarsus gr. lobatifrons |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
|
Всего видов – 102 вида и таксона рангом выше вида |
78 |
28 |
32 |
7 |
32 |
25 |
11 |
8 |
17 |
Примечение : *ерик Дарма, проток Дамчик; западные подстепные ильмени
Дальнейшие сборы хирономид позволили нам установить новый для науки вид, характерный для северной части Каспийского моря Сhironomus paralbidus sp.n (идентификация Н.В. Полуконовой, сборы личинок Д.С. Даировой), морфологически и кариотипически близкий к видам группы C. salinarius [39]. К сожалению, дальнейшие исследования распределения видов и их биологии не проводились.
На фоне многолетней динамики биомассы бентоса и личинок хирономид в Северном Каспии выделяется период 1981-1984 гг., 1990 гг., характеризующийся общим увеличением количественных характеристик макрозообентоса в Северном Каспии. Данные специалистов-бентологов, приведенные в табл. 2, свидетельствуют о значительном возрастании биомассы хи-рономид на фоне общего увеличения биомассы бентоса в 1980-1990-е годы (табл. 2).
Таблица 2. Многолетние изменения биомассы бентоса и хирономид (г/м 2 ) в Северном Каспии в различные периоды 1939-1969гг.; 1981-84 гг.; 1990 г. [54, 46, 31].
|
Годы |
1939- |
1941- |
1947- |
1950- |
1957- |
1959- |
1963- |
1981-1984 |
2001-2005 |
|
1940 |
1946 |
1949 |
1956 |
1958 |
1962 |
1969 |
1990 |
(макс.) |
|
|
Бентос |
9,9 |
23,3 |
26,4 |
23,7 |
68,8 |
73,3 |
49,5 |
64,2 |
51,3 |
|
Хирономиды |
0,3 |
0,3 |
0,4 |
0,2 |
0,5 |
0,3 |
0,2 |
1,3 |
0,8 |
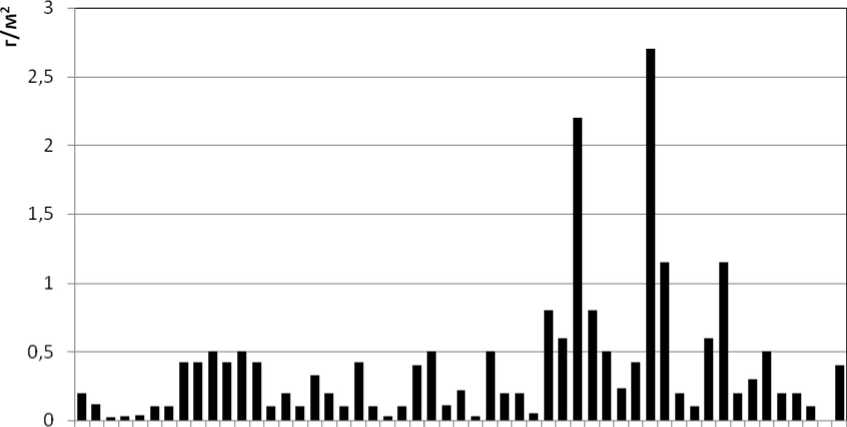
Подтверждением этому служат результаты мониторинговых изменений биомассы хирономид и бентоса в Северном Каспии, где пики биомассы хирономид (более 2 г/м 2 ) характерны для 1982 и 1987 годов (рис. 1, 2). Дальнейшие изменения структуры донных сообществ характеризуются снижением общей численности и биомассы хиро-номид в Северном Каспии [31] на фоне продолжающегося снижения кормовой базы бентосояд-ных рыб, характерного для отдельных районов Северного Каспия.
Анализ различных преобразований экосистемы Каспия, где донным гидробионтам отведена определяющая роль кормовой базы промысловых бентосоядных рыб, приводится в ряде публикаций [10, 29, 30, 32, 46, 48].
Наблюдаемые структурные изменения донных сообществ сопровождаются колебанием численности и биомассы популяций хирономид как в Северном Каспии, так и в водоемах авандельты р. Волги и обусловлены многими факторами, главными из которых являются водность года, уровенный режим, продолжительность паводка, что оказывает непосредственное влияние на биотопи-ческое разнообразие, питание, условия для развития ценоза хирономид. В свою очередь, темпера- турный и ветровой режим весной и в начале лета определяют условия роения имаго, откладку яиц самками и вылупление личинок хирономид, что в конечном итоге сказывается на величине популя ций хирономид.
ОООГМ<а-ЮОООГМ<9-ЮоаОГМ<»ЮОООГМ<ТЮОООГМ<»ЮООО ^тшьлшьл^^^ю^г^г^Г'Г^г^ oooooooooocjxa^crta^crto oScrtoo^o^crto^o^o^o^^o^crto^o^o^^o^o^o^o^o^o^o^o^o М^-<^ЧгНтНгН^ЧгН^-<1НгН^Ч^-<гЧтНтН^ЧгН^Ч^-<1Нг-1^Н^-<гНтНГ\Г
годы
Рисунок 1. Многолетняя динамика биомассы Chironomidae в Северном Каспии
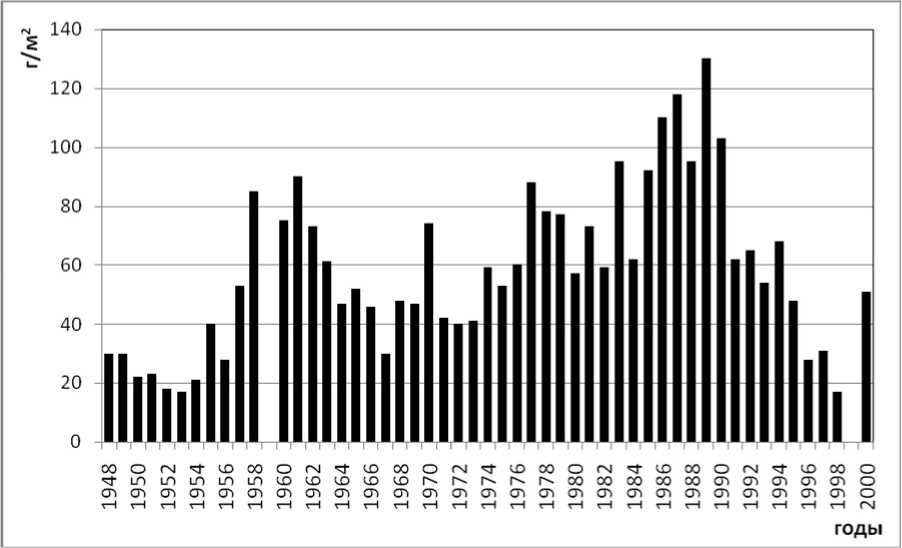
Рис. 2. Многолетняя динамика общей биомассы зообентоса в Северном Каспии
Формирующими факторами для хирономидо-фауны Северного Каспия, наряду со специфическими абиотическими условиями региона, являются биотические – наличие обширных незанятых пространств и свободных экологических ниш, которые появляются как при снижении уровня моря, так и во время его повышения.
ЗАКЛЮЧЕНИЕ
Произошедшие значительные изменения гидрологического режима Каспийского моря и его экосистемы с учетом антропогенного и техногенного воздействия разного генезиса, оказывают в настоящее время определяющее влияние на состав, структуру и распределение донных биоце- нозов. Разнообразие и динамика природных условий Каспийского моря обусловливают особенности формирования уникальной фауны. К сожалению, мелководная зона Северного Каспия, являющаяся биотопически благоприятной для развития таксономически разнообразной хирономи-дофауны Северного Каспия, не входит в зону мониторинговых исследований. Не смотря на приведенные нами данные о составе хирономид отдельных участков, многолетней динамики биомассы личинок хирономид в глубоководных районах Северного Каспия, следует констатировать, что современное состояние хирономидофауны как в низовьях Волги, так и в Северном Каспии продолжает оставаться белым пятном при изучении многолетней динамики сообществ макрозообентоса, что лишает имеющиеся сведения о кормовой базе Каспия, особенно его мелководных районов, объективной информативной направленности и практического использования для адекватной оценки продуктивности. Вместе с тем всестороннее изучение Chironomidae, включающее оценку численности, биомассы и таксономического богатства, является неотъемлемым и существенным в биомониторинге водотоков низовьев Волги и Северного Каспия.
Авторы благодарны зав. лабораторией Касп-НИРХа В.Ф. Осадчих за предоставленные пробы, содержащие личинок хирономид из Северного Каспия. Работа выполнена в рамках программы Президиума РАН «Биологическое разнообразие», раздела «Динамика биоразнообразия и механизмы обеспечения устойчивости биосистем» и при частичной поддержке грантов РФФИ (гранты № 07-04-96610, № 13-04-00740А).
Список литературы Изучение хирономид (Diptera, Chironomidae) в низовьях Волги и в Северном Каспии и их многолетнее изменение биомассы
- Абдурахманов Г.М., Сокольская Е.А., Сокольский А.Ф. Моделирование экологического прогноза состояния бентоса в Северном Каспии. 2009. http://www.dagecolog.ru/jurnal/032009/sokolski.htm.
- Абдурахманов Г.М., Сокольская Е.А., Калтаев А. Характеристика бентоса северо-восточного Каспия, технология добычи и потенциальное воздействие разработок месторождений нефти и газа на окружающую среду. 2009. http://www.dagecolog.ru/jurnal/032009/kaltaev.htm.
- Абросов В.Н. О видообразовании в озерах/Отв. ред. Г.Г. Винберг; АН СССР, Всесоюзное гидробиологическое общество. М.: Наука, 1987. 86 с.
- Аладин Н.В., Филипов А.А., Петухов В.А., Плотников И.С., Смуров А.О. Гидробиологические исследования Зоологического института РАН в дельте Волги и Северном Каспии в 1994-1997 гг. Ч. 2. Изучение зоопланктона и зообентоса//Касп. Плавучий Ун-т: Науч. бюл. Астрахань, 2000. № 1. С. 93-102.
- Алексевнина М.С. Видовой состав личинок хирономид в дельте Волги//Гидробиологический журнал. 1973. Т. 9. № 3. С. 78-81.
- Алексевнина М.С. Рост и продукция массовых видов хирономид (Diptera, Tendipedidae) авандельты Волги//Зоологический журнал, 1974, Т. 53, № 5, С. 720-727.
- Алексевнина М.С. К биологии и распространению хирономид Fleuria lacustris Kieff. (Chironomidae, Diptera)//Фауна и экология насекомых: межвузовский сб. научных трудов. Пермь. 1981. С. 140-144.
- Алексевнина М.С., Белянина С.И. О биологии Fleuria lacustris Kieff. (Chironomidae, Diptera) из авандельты Волги./Физиологическая и популяционная экология животных. 1974. Вып. 2(4). Саратов. С. 153-156.
- Атлас беспозвоночных Каспийского моря/Под.ред. Бирштейна Я.А., Виноградовой Л.Г.: М: Пищевая промышленность,1968. 430 с.
- Беляева В.Н., Катунин Д.И., Осадчих В.Ф. Влияние колебания уровня моря на формирование биологической и рыбной продуктивности Северного Каспия//Биологические ресурсы Каспийского моря (Тез. докл. 1-ой Междунар. конф.). Астрахань, 1992. С. 38-42.
- Даирова Д.С., Зинченко Т.Д. История исследований макрозообентоса Нижней Волги и ее дельты//Изв. СамНЦ РАН. 2003. Спец. вып. № 1. С. 91-101.
- Зайцев В.Ф., Калиманова Д.Ж. Сравнительная характеристика состояния зообентоса восточной части Северного Каспия//Вестник АГТУ, 2007. № 2 (37). С. 194-196.
- Зинченко Т.Д. Хирономиды поверхностных вод бассейна Средней и Нижней Волги (Самарская область). Эколого-фаунистический обзор. Самара: ИЭВБ РАН, 2002. 174 с.
- Зинченко Т.Д. Биоиндикация как поиск информативных компонентов водных экосистем (на примере хирономид Diptera, Chironomidae)//Чтения памяти В.Я. Леванидова. 2005. Вып.3. Владивосток: Дальнаука. С. 338-359.
- Зинченко Т.Д. Эколого-фаунистическая характеристика хирономид (Diptera, Chhironomidae) малых рек бассейна средней и нижней Волги (Атлас). Тольятти: Кассандра. 2011. 258 с.
- Зинченко Т.Д., Алексевнина М.С. Изменения хирономидофауны (Diptera, Chironomidae) в низовьях дельты Волги и в Каспийском море в связи с подъемом его уровня//Материалы Х Рос. симпоз. по хирономидам «Экология, эволюция и систематика хирономид». Тольятти; Борок, 1996. С. 154-165.
- Иванов В.П., Сокольский А.Ф. Научные основы стратегии защиты биологических ресурсов Каспийского моря от нефтяного загрязнения. -Астрахань.: КаспНИРХ, 2000. 178 с.
- Карпинский М.Г. Экология бентоса Среднего и Южного Каспия. М.: Изд-во ВНИРО, 2002. 283 с.
- Карпинский М.Г. Еще раз к вопросу о глубоководной донной фауне Каспия//Jour. Os Siberian Federal University Biology. № 3. 2010. C. 322-334.
- Карпевич А.Ф. Потребление корма Pontogammarus maeoticus Каспийского моря//Зоологический журнал. 1946. Т. 25. Вып. 6. С. 517-522.
- Катунин Д.Н., Беспарточный Н.П., Хрипунов И.А. Особенности гидролого-гидрохимического режима Каспийского моря//Научные основы устойчивого рыболовства и регионального распределения промысловых объектов Каспийского моря. М.: Наука. 1998. С. 128-142.
- Катунин Д.Н., Егоров С.Н., Кашин Д.В., Хрипунов И.А., Галушкина Н.В., Кравченко Е.А., Гуляев В.Ю., Дектярева О.А. Основные черты гидролого-гидрохимического режима нижнего течения р. Волги и Каспийского моря//Вопросы промыслов. океан. Вып. 1. М.: Изд-во ВНИРО, 2004. С. 69-96.
- Катунин Д.Н., Сорокин Р.В. Каналы-рыбоходы дельты реки Волги. Астрахань/Касп. НИИ рыб. хоз-ва, ООО "Касп. рыбопром. компания "Юленаст", Каспрыбпроект. 2010. 126 с.
- Макарова Р.Е. Водный баланс и уровень моря//Каспийское море. Гидрология и гидрохимия. М.: Наука. 1986. С. 29-38.
- Макарченко Е.А., Макарченко М.А. Chironomidae. Комары-звонцы//Определитель пресноводных беспозвоночных России и сопредельных территорий. Т. 4. Высшие насекомые. Двукрылые. СПб: ЗИН РАН, 1999. С. 210-296.
- Малинин В.Н. Проблема прогноза уровня Каспийского моря. СПб: Изд-во. РГМИ, 1994. 160 с.
- Малиновская Л.В. Особенности развития зообентоса Северного Каспия в условиях продолжающегося подъема уровня вод//Х Междунар. конф. по промысловой океанологии. Тез. докл. (СПб. 29-23 мая 1997), М.: Изд.-во ВНИРО. 1997. С. 82.
- Малиновская Л.В. Состояние зообентоса Северного Каспия в летний период//Осетровое хозяйство водоемов СССР. Ч.1. Астрахань. 2007. С. 216-217.
- Малиновская Л.В., Зинченко Т.Д, Кочнева Л.А. Современная оценка макрозообентоса Каспийского моря как объекта питания бентосоядных рыб (на примере восточного района Северного и Среднего Каспия).//Аграрная Россия, 2008. № 3, С. 29-34.
- Малиновская Л.В., Зинченко Т.Д. Многолетняя динамика сообществ макрозообентоса Северного Каспия.//Изв. Сам НЦ РАН, 2010. Т. 12. № 1. С. 179-184.
- Малиновская Л.В., Кочнева Л.А. Состояние донной фауны Каспийского моря//Рыбохозяйственные исследования на Каспии. Результаты НИР за 2004 г. Астрахань: КаспНИРХ. 2005. С. 140-148.
- Молодцова А.И. Питание осетра в Каспийском море//Рыбохозяйственные исследования на Каспии. Изд-во КаспНИРХ. 2001. Астрахань. С. 125-129.
- Моря СССР. Моря и океаны России. Атлас морских млекопитающих. (Приматы моря. 2011-2013). http://www.bruo.ru/pages/142.html.
- Научные основы устойчивого рыболовства и регионального распределения промысловых объектов Каспийского моря/Под ред. В.Н. Беляевой, В.П. Иванова, В.К. Зиланова. М.: Изд-во ВНИРО, 1998. 167 с.
- Осадчих В.Ф. Бентос Северного Каспия в условиях зарегулирования стока р.Волги//Зоологический журнал. Т. 42. Вып. 2. 1963. С. 184-196.
- Осадчих В.Ф. Изменение биомассы бентоса в Северном Каспии за последнее пятилетие//Труды КаспНИРХ. Т. 24. 1968. С. 100-112.
- Осадчих В.Ф., Ардабьева А.Г., Белова Л.Н., Елизаренко М.М., Курашова Е.К., Малиновская Л.В., Попова М.К., Степанова Т.К., Тиненкова Д.Х., Татаринцева Т.А. Особенности развития и использования кормовой базы рыбами в условиях повышения уровня Каспийского моря//Комплексные рыбохозяйственные исследования на Каспии. М. 1989. С. 119-138.
- Полуконова Н.В., Белянина С.И., Зинченко Т.Д. Chironomus paraalbidus sp. n. (Chironomidae, Diptera) из Каспийского моря//Зоологический журнал, 2005. Т. 84, № 8. С. 1017-1024
- Полянинова А.А. Виды вселенцы в Каспии и их роль в экосистеме моря. Астрахань: КаспНИРХ, 2007. 104 с.
- Панин Г.Н., Мамедов Р.М., Митрофанов И.В. Современное состояние Каспийского моря. М.: Наука, 2005.
- Родионов С.С. Современное изменение климата Каспийского моря. М.: Гидрометеоиздат. 1989. 124 с.
- Романова Н.Н. Методические указания к изучению бентоса южных морей СССР. М., 1983. 13 с.
- Салманов М.А. Экология и биологическая продуктивность Каспийского моря. 1999. Баку. 398 с.
- Салманов М.А. Экологическое состояние основ биологической продуктивности Каспийского моря//Каспийский Плавучий Ун-т. Науч. бюл. 2000. № 1. С. 43-50.
- Свиточ А.А, Селиванов А.О., Янина Т.А. Палеогеография событий плейстоцена Понто-Каспия и Средиземноморья (материалы по реконструкции и корреляции) М.: РАСХН. 1998. 315 с.
- Смирнова Л.В. Динамика донной фауны в Северном Каспии//Биологические ресурсы Каспийского моря. Тез. докл. I-ой Междунар. конф. Астрахань, 1992. С. 364-366.
- Степанова Т.Г., Сокольский А.Ф. Естественные враги гребневика Мnеmiopsis leidyi в Каспийском море//Междунар. конф. «Рыбохозяйственная наука на Каспии: задачи и перспективы» (Махачкала 15-18 июня 2003 г.). Астрахань, 2003. С. 145-147.
- Стыгар В.М., Мутышева Г.К. Зообентос восточной части Северного Каспия в зоне предполагаемой разработки месторождений углеводородного сырья в период стабилизации уровня моря/Морские гидробиологические исследования. Сборник научных трудов. М.: Изд-воВНИРО, 2000. С. 111-127.
- Тарасов А.Г. Многолетние количественные исследования донных биоценозов Северного Каспия//Океанология. 1998. Т. 38, № 1. С. 105-113.
- Фильчаков В.А. Состав и многолетние изменения зообентоса в предустьевом взморье Каспия//Биологические ресурсы Каспийского моря: Тез. докл. I-ой Междунар. конф. Астрахань, 1992. С. 438-441.
- Фильчаков В.А. Фауна, экология и распределение перакарид (бокоплавы, мизиды, кумовые и равноногие ракообразные) в водоемах дельты Волги: Автореф. дис.. канд. биол. наук. СПб., 1994. 24 с.
- Хрипунов И.А., Д.Н. Катунин, А. В. Азаренко. Многолетние изменения гранулометрического состава и пространственного распределения донных отложений Северного Каспия//Водные ресурсы. 2010. Т. 37, № 6. С. 709-716
- Шилова А.И., Зеленцов Н.И. Фауна хирономид (Diptera, Chironomidae) бассейна Верхней Волги//Биология внутренних вод. 2003. № 2. С. 27-34.
- Яблонская Е.А., Осадчих В.Ф. Изменение кормовой базы бентосоядных рыб Северного Каспия//Тр. ВНИРО. Т. 80. 1973. С. 48-73.
- Freshwater biomonitoring and benthic macroinvertebrates. N.Y.: Chapman and Hall, 1993. 488 p.
- Malinovskaya L.V., Zinchenko T.D. Mytilaster lineatus (Gmelin): Long Term Dynamics, Distribution of Invasive Mollusk in the Northern Caspian Sea. ISSN 2075-1117, Rus. Jur. of Boilogical Invasions 2010, Vol. 1. № 4, pp. 288-295.
