Изучение особенностей аккумуляции ионов тяжелых металлов водными растениями и роли липидов в адаптации к ним

Автор: Розенцвет O.A.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1 т.8, 2006 года.
Бесплатный доступ
Изучены особенности аккумуляции ионов тяжелых металлов (ТМ) - (Al+3, Cu+2, Cd+2, Pb+2, Zn+2) - водным растением Potamogeton perfoliatus. Степень накопления ионов ТМ зависела от экзогенных факторов (концентрации, времени экспозиции, природы ТМ) и эндогенных фазных органов растения, возраста). Выявлено, что действие ионов ТМ приводит к изменениям в липидном метаболизме: изменении соотношения липидных классов, индивидуальных фосфо- и гликолипидов.
Короткий адрес: https://sciup.org/148197814
IDR: 148197814 | УДК: 628.581:13.577.7.15
Studying of features of accumulation of ions of heavy metals by water plants and roles липидов in adaptation to heavy metals
Are investigated features of accumulation of ions of heavy metals (TM) (Al+3, Cu+2, Cd+2, Pb+2, Zn+2) by water plant Potamogeton perfoliatus. The degree of accumulation of ions TM depend экзогенных factors (concentration, time of an exposition, nature TM) and эндогенных (different bodies of a plant, age). It is revealed, that action of ions TM results in changes in липидном a metabolism: change of a parity (ratio) липидных classes, individual фосфо-and гликолипидов.
Текст научной статьи Изучение особенностей аккумуляции ионов тяжелых металлов водными растениями и роли липидов в адаптации к ним
Институт экологии Волжского бассейна РАН,г.
Изучены особенности аккумуляции ионов тяжелых металлов (ТМ) - (Al+3, Cu+2, Cd+2, Pb+2, Zn+2) -водным растением Potamogeton perfoliatus. Степень накопления ионов ТМ зависела от экзогенных факторов (концентрации, времени экспозиции, природы ТМ) и эндогенных (разных органов растения, возраста). Выявлено, что действие ионов ТМ приводит к изменениям в липидном метаболизме : -нении соотношения липидных классов, индивидуальных фосфо- И гликолипидов .
Проблема повышенного содержания тяже-лых металлов (ТМ) в окружающей среде с каждым годом приобретает все большую ак-туальность. Металлы представляют серьез-ную угрозу для биоты вследствие их острой токсичности для организмов и постепенно-го накопления их в окружающей среде до опасного уровня. Уровень, при котором они становятся действительно опасными, зави-сит не только от степени загрязнения ими окружающей среды, но и от химических осо-бенностей металла и его биохимического цикла [1].
В отличие от других факторов среды, вли-яние которых на живые объекты, равно как и ответные реакции живого на эти факторы, хорошо изучены, эколого-биохимические ме-ханизмы адаптации к воздействию ТМ ис-следованы в меньшей степени . Особенно это касается водных экосистем и водных обита-телей, поскольку вопросы адаптации водных организмов к загрязнению окружающей сре-ды имеют в настоящее время первостепен-ное значение [2].
Учитывая опасность ТМ для здоровья че-ловека, исследование механизмов токсично-сти ТМ на объекты различного уровня орга-низации проводятся во многих лаборатори-ях мира [3-7]. При этом усилия исследовате-лей направлены как на решение фундамен-тальных задач , связанных с изучением меха-низмов адаптации растительные организмов к изменениям условий обитания, так и при-кладных задач , таких как фиторемидиация загрязненных территорий [8].
Целью данной работы было сравнитель-ное изучение особенностей аккумуляции ионов ТМ водным растением Potamogeton perfoliatus (рдест пронзеннолистный), а так-же роли одного из важнейших компонентов жизнедеятельности клетки - липидов в про-цессах адаптации данного вида растений к этому виду загрязнений окружающей среды .
Результаты и их обсуждение
При изучении влияния загрязнения вод-пых источников солями тяжелых металлов и их влияния на водные растения было исполь-зовано натурное моделирование с примене-нием мезокосмов, устанавливаемых непос-редственно в водоеме. Использование этого метода позволяло наиболее полно учесть всю совокупность абиотических и биотических факторов, определяющих поведение ТМ в це-лостной гидросистеме [9]. Согласно этой ме-тодике, в мезокосмы (в нашем случае, ЦИЛИН-дрические окрашенные емкости объемом 250 л, отграничивающие до дна несколько расте-ний вместе с корнями), изолирующие учас-ток материнской экосистемы с сохранением ее основных параметров, вводили заданные порции ТМ в виде нитратных солей. Конт-ролем служили растения, отобранные до на-чала эксперимента из каждой емкости, рас-
, ( ),
, -
,
( ).
В качестве исследуемых металлов были : Cu, Cd,
Pb, Zn Al.-
. -, -
-
Hg, Pb, Cd, As, Zn,-
,-
-
Из этого списка приоритетных загрязнителей Cd, Pb, Zn, наиболее распространенные и актуальные для акваторий Саратовского и Куйбышевского
.,
.-
, , ,-
Cd, Pb, Zn-
, ,-
--
,-
.,
,,
,-
,,
.
С химической точки зрения ТМ являются металлами, имеющими свободные d- орб ита-
. ,, -
, Zn Al,-
, Zn-
,
[11].,
-
.-
- трациях эти металлы способны оказывать
,
.-
-
.-
,-
,,
,
.
-копление ТМ в макрофитах отличается от
.-
-
[12,
13].- ренные элементы поглощаются из воды всей поверхностью и аккумуляция определяется
-
.-
-
-
-
, .
, ,- кумуляции ионов используемых металлов в различных органах исследуемого водного
.- тов по динамике поступления ТМ в листья . 1.
,-
, в присутствии ТМ в течение трех суток было
.
Через три дня содержание меди составило 74,4, 33,0 25,3 /
-
, 1000 / .
. 1, -
.
Исследуемый вид рдеста наиболее активно
.
-
[14-16].
-
, -
-
. . -
-
-
, , - ная способность листьев рдеста уменьшалась
.
Таблица 1.Динамика аккумуляции ионов ТМ в листьях Potamogeton perfoliatus по результа-там атомно-адсорбционного анализа
|
Концентрация металла в водной среде, мкмоль/л |
Концентрация металла в листьях, мг на г сухой массы |
||
|
Время экспозиции, сут. |
|||
|
1 |
2 |
3 |
|
|
Cd 1 |
0,3 |
0,5 |
0,7 |
|
Cd 10 |
1,0 |
1,1 |
3,7 |
|
Cd 100 |
12,4 |
22,3 |
24,7 |
|
Cd 1000 |
15,3 |
32,5 |
33,0 |
|
Cu 1 |
0,1 |
0,1 |
0,2 |
|
Cu 10 |
1,0 |
2,2 |
4 ,4 |
|
Cu 100 |
11,5 |
23,7 |
24,6 |
|
Cu 1000 |
44,3 |
68,0 |
74,4 |
|
Zn 10 |
0.3 |
0,4 |
0,6 |
|
Zn 100 |
4,2 |
13,3 |
15,7 |
|
Zn 1000 |
11,3 |
25,2 |
25,3 |
|
Pb 100 |
2,8 |
3,5 |
3,9 |
|
Al 100 |
3,2 |
3,6 |
4,4 |
Было проанализировано также накопле-ние металлов в разных органах Potamogeton perfoliatus (корнях, стеблях, листьях) и в лис-тьях разного возраста (табл. 2). При этом раз-деление листьев на разные возрастные кате-гории проводились следующим образом . Листья розетки, обычно 1-4 ярус , принима-лись за молодые листья, листья 5-9 ярусов -за листья среднего возраста и листья ниже 9 яруса - за старые листья.
Таблица 2. Содержание ионов металлов в разных частях растения Potamogeton perfoliatus через одни сутки воздействия при начальной концентрации солей металлов в воде 100 мкмоль/л
|
Часть растения |
Cu |
Cd |
Zn |
Pb |
Al |
|
Молодые листья |
3,7 |
11,1 |
2,3 |
3,9 |
3,6 |
|
Зрелые листья |
8,3 |
6,9 |
5,6 |
3,8 |
3,6 |
|
Старые листья |
17,8 |
6,9 |
6,2 |
2,9 |
2,5 |
|
Стебли |
0,6 |
6,6 |
0,5 |
0,5 |
8,0 |
|
Корни |
0,3 |
1,6 |
0,2 |
0,3 |
1,8 |
Данные табл . 2 показывают, листья, но и стебли и корни, аккумулировали металлы . Причем степень накопления в раз- ных органах растения зависела от природы металла. Так, медь максимально накаплива-лась в ранее сформированных старых листь-ях , кадмий и свинец - в молодых макушеч-НЫХ листьях, а алюминий - в стеблях. Корни растения менее всего аккумулировали ионы ТМ .
Таким образом, несмотря на то,что ввод-ных растениях воздействию металлов под-вергаются все органы одновременно , распре-деление ионов металлов в этих органах ока-залось весьма неравнозначным и зависело от природы металла.
Сравнительное исследование роли липи-дов в ответных реакциях на воздействие ионов ТМ проводили по следующим показа-телям : , COOT-ношению липидных классов, соотношению индивидуальных фосфо- и гликолипидов.В табл . 3 приведены данные по содержанию общих липидов ( ОЛ ) через трое суток воздей-ствия всех испытанных металлов при началь-ных концентрациях 10 и 100 мкмоль/л. Из этих данных видно, что действие ионов ТМ приводило к снижению количества ОЛ в той или иной степени . Но наиболее токсичными для липидов рдеста оказались ионы меди и кадмия, которые, по-видимому, -ли синтез новых липидов , в результате чего разрушение липидов не восполнялось вновь синтезированными липидами и количество ОЛ снижалось в 1,5-2 раза.
Таблица 3. Зависимость изменения коли-чества общих липидов в листьях Potamoge-ton perfoliatus от природы ТМ, время экспо-зиции - трое суток
|
Концентрация металла в водной среде, мкмоль/п |
Содержание общих липидов, мг/г сырого веса |
|
Фон |
22,4± 1,8 |
|
Контроль |
20,1±1,6 |
|
Cu 10 |
9,4 ± 1,3 |
|
Cu 100 |
7,7 ± 0,8 |
|
Cd 10 |
13,5 ± 1,1 |
|
Cd 100 |
9,3 ± 0,8 |
|
Zn 10 |
16,8± 0,8 |
|
Zn 100 |
14,9 ± 1 |
|
Pb 10 |
15,8 ± 3,4 |
|
Pb 100 |
12,1 ± 2,1 |
|
Al 100 |
18,5±0,9 |
Таким образом, первый «ответ» липидов растений на присутствие ионов металлов в среде был связан со снижением количества синтезируемых липидов .
Дальнейшее определение содержания ос-новных классов липидов в данных условиях показывал на перестройку метаболизма ли-ПИДОВ , проявляющейся в изменении соотно-шения гликолипидов ( гл), фосфолипидов ( ФЛ ) и нейтральных липидов (НЛ). При этом разные металлы и разные концентрации этих металлов имели различный эффект на состав липидов . Сравнительное исследование вли-яния всех исследованных металлов в равных условиях на соотношение липидных классов приведено в табл. 4. Данные этой таблицы показывают, имеет разную направленность. Если ионы меди при 100 мкмоль/л и кадмия при 10 и 100 мкмоль/л действовали таким образом, что снижалось содержание полярных липидов, как ФЛ, так ГЛ, то действие ионов свинца при двух испытанных концентрациях (10 и 100 мкмоль/л) было направлено на снижение сии-теза хлоропластных ГЛ . Ионы цинка вызы-вали меньшие изменения в соотношении ли-пидных классов, хотя при концентрации 100 мкмоль/л наблюдалось снижение содержания ФЛ по сравнению с контрольными образца- МИ . Но это снижение содержания ФЛ было заметно меньше в сравнении со снижением количества ФЛ, наблюдаемое при действии ионов кадмия и меди.
Таблица 4. Соотношение нейтральных (НЛ), глико- ( ГЛ ) и фосфолипидов ( ФЛ ) в листьях Potamogeton perfoliatus в зависимости от концентрации и природы ТМ. Время экспо-зиции - трое суток
|
Концентрация солей, мкМ |
Содержание липидов, %от суммы |
||
|
НЛ |
ГЛ |
ФЛ |
|
|
Фон |
28,5 ± 2,3 |
41,0 ± 3,9 |
30,5 ± 2,4 |
|
Контроль -1 |
30,0 ± 1,7 |
41,6 ± 4,9 |
28,4 ± 0,7 |
|
Cu - 10 |
29,9 ± 0,8 |
38,8 ± 0,5 |
31,3 ± 0,8 |
|
Cu - 100 |
53,0 ± 7,3 |
28,1 ± 3,3 |
18,8 ± 1,9 |
|
Cd - 10 |
48,6 ± 1,4 |
36,0 ± 2,8 |
15,2 ± 2,8 |
|
Cd -1 00 |
57,9 ± 4,1 |
26,4 ±0,2 |
15,8 ± 1,5 |
|
Zn - 10 |
27,2 ± 2,2 |
43,2 ± 3,8 |
29,5 ± 4,3 |
|
Zn - 100 |
35,4 ± 1,6 |
40,5 ± 3,1 |
23,8 ± 3,6 |
|
Pb - 10 |
32,7 ± 4,5 |
35,6 ± 4,2 |
31,6 ± 2,8 |
|
Pb - 100 |
35,0 ± 0,1 |
30,9 ± 2,1 |
34,0 ± 2,2 |
|
Al - 100 |
26,4 ± 2,5 |
34,1 ± 2,8 |
39,8 ± 4,2 |
Из полученных результатов можно пред- ПОЛОЖИТЬ , что , по-видимому, от специфичности металла затрагиваются разные линии синтеза полярных липидов.
Поскольку полярные липиды становились главной мишенью воздействия ионов ТМ, то более подробно был исследован состав ин-дивидуальных ФЛ под воздействием ионов ТМ . В наших предыдущих исследованиях было показано, что ФЛ данного вида расте-ния состоят из фосфатидилхолина ( ФХ), фос-фатидилэтаноламина (ФЭ), фосфатилглице-рола (ФГ), фосфатидилинозитола (ФИ) и фос-фатидной кислоты (ФК) [17]. Главными в количественном отношении являются ФХ, ФЭиФГ.
при одинаковой концентрации 100 мкмоль/л разных металлов приведены на рис. 1-3.
Все ионы исследуемых ТМ в данной кон-центрации мало влияли на количество ФХ, за исключением ионов кадмия, действие ко-торых приводило к снижению его уровня.в отличие от ФХ содержание ФЭ под действи-ем некоторых металлов менялось драмати-чески . Причем в изменении данного липида особенно четко проявилась специфика каж-дого из исследуемых ТМ . Если динамика из-менения этого липида под действием ионов цинка не отличалась от контроля, то под дей-ствием ионов алюминия количество ФЭ уве-личивалось . Но действие ионов меди, кадмия и свинца было направлено на снижение ко-личества ФЭ вплоть до полного исчезнове-ния из состава ФЛ. Специфичность воздей-ствия разных ТМ проявилась и в изменении содержания ФГ.
были индифферентны к ФГ, меди и кадмия приводило к увеличению вкла-да ФГ, .
Таким образом, на примере главных ФЛ выявлены устойчивые и чувствительные ли-нии синтеза индивидуальных ФЛ в зависи-мости от специфики металла и действующей концентрации этих металлов.
Одной из мишеней при атаке иона метал-ла становятся хлоропластные липиды, о чем свидетельствует увеличение доли ФГ - фос-фолипида, который стабилизирует светосо-бирающий комплекс ФС II. Это подтвержда-
ФХ
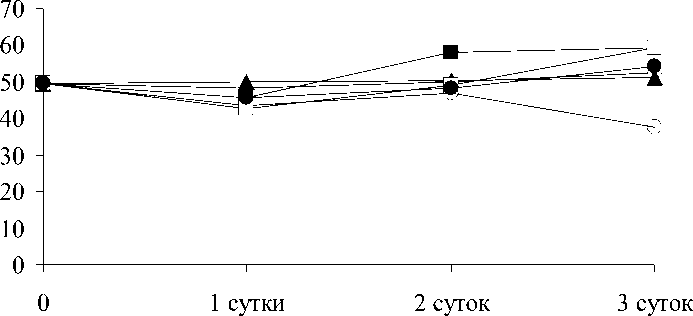
Время экспозиции
Zn Cu Cd Pb Al
. 1.
Potamogeton perfoliatus
100 / 3- ется и соотношением липидных классов
(. 4)
( ) ( . 4-6).- ния ионов всех изученных ТМ при равных
-
.
49,0-54,5% .- действии ионов цинка в течение трех суток
44,1
-59,7%, ионов кадмия - 41,7-35,5%, ионов
ФЭ
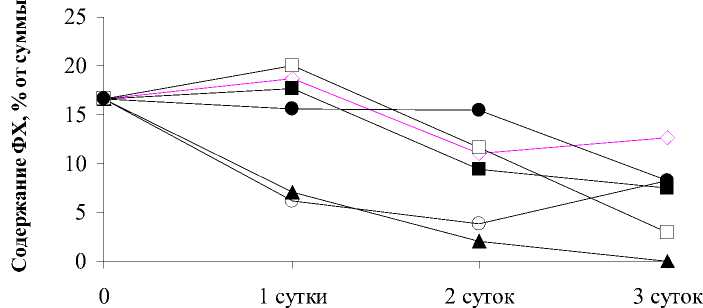
Время экспозиции
Zn Cu Cd Pb Al
. 2.
Potamogeton perfoliatus
100 / 3-
ФГ
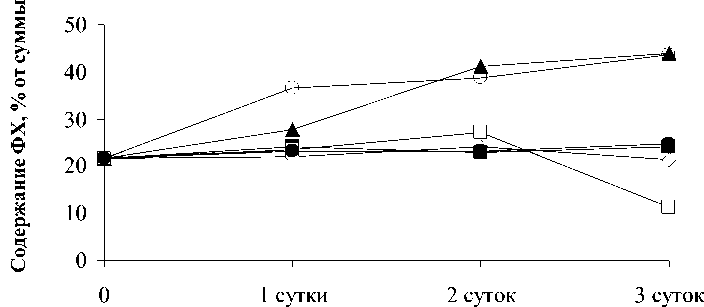
Время экспозиции
Zn Cu Cd Pb Al
. 3.
Potamogeton perfoliatus
100 / 3
48,2-60,8%,39,2
52,4%, - 48,6-49,1%-
. ,-
-
.-
,
( . 4).
В отношении второго галактолипида ДГДГ
,
33,6%
36,3% -
.
разных концентраций нитрата цинка за этот
-
32,9-36,0%,
.
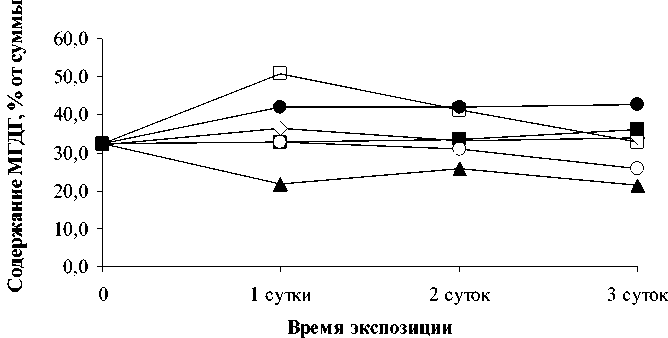
мгдг
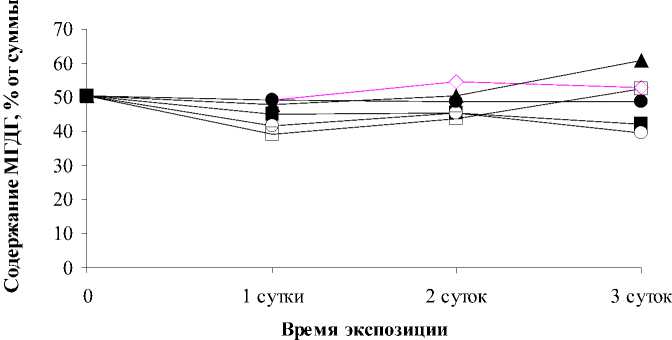
Zn 100 Cu 100 Cd 100
Pb 100 Al 100
. 4. Potamogeton perfoliatus при воздействии исследованных ионов ТМ в концентрации 100 мкмоль/л
ДГДГ
Zn 100 Cu 100 Cd 100
Pb 100 Al 100
. 5. Potamogeton perfoliatus
100 /
-
.-
-
24,6 -34,0%,
20,7-29,4%, 32,9-50,9%,
41,8-42,8%.
,- вали в направлении снижения количества
.,
-
,-
( . 5).
-
,, свинца и снижалось под действием ионов
( . 6).
,
,- схдг

Zn 100 Cu 100 Cd 100
Pb 100 Al 100
. 6. Potamogeton perfoliatus при воздействии исследованных ионов ТМ в концентрации 100 мкмоль/л
чае с фосфолипидами, выявлен специфичес-кий отклик на воздействие разных ТМ.
Таким образом, изучены особенности ак-кумуляции ионов тяжелых металлов (Al+3, Cu+2, Cd+2, Pb+2, Zn+2) водным растением Potamogeton perfoliatus. Степень накопления ионов ТМ зависела от экзогенных факторов (концентрации, времени экспозиции , приро-дыТМ) и эндогенных (разных органах расте-НИЯ , возраста). На примере одного вида вод-ных растений показана роль липидов в от-ветных реакциях на воздействие ионов ТМ как одного из неблагоприятных абиотичес-ких факторов антропогенного характера. Не-смотря на разную направленность действия ионов ТМ, общая роль липидов заключается в изменении соотношения липидных классов,
. Аналогичные изменения были характерны для растений при встрече с другими стрес-совыми воздействиями (низкая температура, соленость, гипоксия ит. .) [18]. Это говорит о том , что механизмы адаптации (реагирова-НИЯ) живой системы на внешние воздействия на биохимическом уровне являются достаточ-но однотипными и рациональными . В то же время из полученных результатов можно еде-лать вывод, что растительная клетка имеет в своем арсенале набор приспособительных механизмов, которые она реализует, тем неблагоприятным факторам, которых она ранее не встречала.
Объекты и методы исследования
На мелководном участке акватории Сара-товского водохранилища с обильными зарос-лями рдеста пронзенолистного (Potamogeton perfoliatus L.) были установлены емкости, от-граничивающие до дна 10-12 растений вме-сте с корнями. В емкостях поддерживали объем природной воды 135-140 л. В течение трех суток через каждые 24 ч в емкости до-бавляли Cd (NO3)2 для поддержания концен-траций - 1, 10, 100 и 1000 мкмоль/л. Для ана- лизов использовали листья , стебли и корни . Отобранные для анализа образцы раститель-ного метериала тщательно промывали в про-точной воде, подсушивали фильтровальной бумагой , взвешивали и использовали для ана-лиза содержания липидов . Полученные дан-ные, приведенные в таблицах и на рисунках, представляют средние значения трех биоло-гических повторностей.
Экстракцию липидов проводили по мето-ду Блайя и Дайера [19]. Липиды разделяли методом тонкослойной хроматографии (тех) на пластинках (10х10 см и 6X6 см) с закреп-ленным слоем силиказоля (эстонская фирма «Хаапсалу»). Для анализа гликолипидов ис-пользовали систему растворителей :
бензол - вода (91:30:8). Фосфолипиды ана-лизировали двумерной ТСХ с применением систем растворителей :
- бензол - аммиак (130:60:20:12)- первое на-правление и хлороформ - метанол - бензол -ацетон - уксусная кислота (140:60:20:10:8) второе направление . Общее количество ли-пидов определяли гравиметрическим спосо-бом . Количественное определение фосфоли-пидов проводили методом Васьковского [20], гликолипидов - по содержанию галактозы с антроновым реагентом [21].
Определение содержания ТМ в тканях ра-стений проводили по методу Голубкиной [22]. Навеску воздушно-сухого растительно-го материала (30-50 мг) заливали смесью кон-центрированной азотной (1,5 мл) и хлорной (0,8 мл) кислот и оставляли на сутки. Затем с помощью термостата пробы прогревали по 1 часу последовательно при 1200С, 1500С и 1800С и после охлаждения до 1500С вносили в них по 5-6 капель концентрированной 2О 2. Через 10 мин к пробам приливали 1 мл 6 М раствора НСl и выдерживали их 10 мин при температуре 1100С. После обесцвечива-ния растворов концентрацию ТМ измеряли на атомно-абсорбционном спектрофотомет-ре Hitachi-207 (Япония).
Список литературы Изучение особенностей аккумуляции ионов тяжелых металлов водными растениями и роли липидов в адаптации к ним
- Бингам Ф.Т., Коста М., Эйхенбергер Э. и др. Некоторые вопросы токсичности ионов металлов/Под ред. X. Зигель, А. Зигель. М.: Мир, 1993.
- Ипатова В.И. Адаптация водных растений к стрессовым абиотическим факторам среды. М.: "Графикон-принт" 2005.
- Maksymiec W., Russa R., Urbanik-Sypniewska T., Baszynski T. Changes in acyl lipid and fatty acid composition on thylakoids of copper nontolerant spinach exposed to excess copper//J. Plant Physiol. 1992. V. 140.
- Kupper H., Kupper F., Spiller M. In situ detection of heavy metal substituted chlorophylls in water plants//Photosynthesis Res., 1998. V.
- Rama Deli S., Prasad M.N.V. Membrane lipid alteration in exposed plants//Heavy metal stress in plants. From molecules to ecosystems. Berlin: Springer, 1999.
- Stefanov K., Kimenov G., Popova I., et al. Lipid and sterol changes in plants and invertebrates caused by environmental pollution//Bulgarian Chemical Comm. 1992. V. 25.
- Guilizzoni P. The role of heavy-metals and toxic materials in the physiological ecology of submersed macrophytes//Aquatic Botany. 1991. V 41, № 1-3.
- Демидчик В.В., Соколик A.И., Юрин B.M. Токсичность избытка меди и толерантность к нему растений//Успехи совр. биол. 2001. Т. 121, №5.
- Ильин В.Б. Тяжелые металлы в системе почва-растение. Новосибирск: Наука, 1991.
- Холодова В.П., Волков К.С., Кузнецов Вл.В. Адаптация к высоким концентрациям солей меди и цинка растений хрустальной травки и возможность их использования в целях фиторемедиации//Физиология растений. 2005. Т. 52, № 6.
- Селезнев В.А., Рубцов М.Г., Купер В.Я., Розенберг Г.С. Оценка пространственной неоднородности качества вод Саратовского водохранилища//Изв. СНЦ РАН. 1999, № 2.
- Maksymiec W. Effects of copper on cellular processes in higher plants//Photosynthetica. 1997. V 34, №3.
- Лукина Л.Ф., Смирнова Н.Н. Физиология высших водных растений. Киев: Наук. думка, 1988.
- Христофорова Н.К. Химико-экологическая характеристика Кандалашского залива Белого моря по содержанию тяжелых металлов в моллюсках и водорослях//Биология моря. 1994. Т. 20, №2.
- Прасад М.Н. Практическое использование растений для восстановления экосистем загрязненных металлами//Физиология растений. 2003. Т. 50, № 35.
- Розенцвет O.A., Мурзаева С.В., Гущина И.А. Роль мембранных липидов в устойчивости Potamogeton perfoliatus L. к избытку Кадмия в воде//Изв. РАН. Сер. биол., 2004. № 6.
- Смоляков Б.С., Жигула M.B., Рыжих A.П., и др. Поведение различных форм меди (II) в пресноводной экосистеме//Водные ресурсы. 2004. Т. 31, № 1.
- Bligh E.G., Dyer W.J. A rapid method for total lipid extraction and purification//Can. J. Biochem. Physiol. 1959. V. 37.
- Vaskovsky V.E., Latyshev L.A. Modified Jungnickel's reagent for detecting phospholipids and other phosphorus compounds on thin-layer chromatography//J. Chromatogr. 1975. V. 115.
- Северин C.E., Соловьева Г.А. Практические работы в биохимии. М.: Изд-во Моск. у-та. 1989.
- Голубкина Н.А. Флуорометрический метод определения селена//Журн. аналит. химии. 1995. Т. 50.
