Изучение популяционной структуры лекарственных растений как основы разработки мер по сохранению генофонда

Автор: Редькина Н.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 2 т.10, 2008 года.
Бесплатный доступ
С использованием изоферментов полиморфных локусов, разделенных электрофорезом в полиакрила- мидном геле, исследована межпопуляционная дифференциация девясила высокого на юге Башкорто- стана. Выявлено отсутствие закономерной пространственной структурированности изменчивости популяций и сравнительная близость структуры большинства из них. Полученные результаты обсуж- даются в отношении проблем сохранения генофонда вида.
Короткий адрес: https://sciup.org/148198101
IDR: 148198101 | УДК: 575.17:
Studies of population structure of medicinal plants in devising conservation of genetic resources
Polymorphic isozyme loci and polyacrylamide gel electrophoresis were used to characterize the patterns of genetic differentiation among populations of elecampane from the southern Bashkortostan. Random spatial structure was revealed and little genetic differentiation was evident among the most part of samples. Problems of gene conservation are discussed based on the information of diversity of the species.
Текст научной статьи Изучение популяционной структуры лекарственных растений как основы разработки мер по сохранению генофонда
В настоящее время проблема сохранения биоразнообразия является одной из глобальных проблем, стоящих перед человечеством, решение которой чрезвычайно важно для устойчивого развития мирового сообщества [4]. В этом плане особое внимание следует уделять редким и исчезающим видам, образующим его наиболее уязвимую часть.
Девясил высокий Inula helenium L. (Asteraceae) - ценное лекарственное растение, которое на Южном Урале находится под охраной государства. Постановлением Кабинета министров Республики Башкортостан № 231 от 11 сентября 2001 г. «О Красной книге Республики Башкортостан» он занесен в «Красную книгу» Республики Башкортостан по 3 категории редкости. К этой группе относят таксоны, представленные небольшими популяциями, которые в настоящее время не находятся под угрозой исчезновения и не являются уязвимыми, но рискуют стать таковыми. Следовательно, исследование генофонда девясила высокого и предложение мер по охране в природных условиях имеет не только высокую научную значимость (вид в этом отношении практически не изучен), но и выраженную прикладную направленность. Актуальность работы вызывается и тем, что растение чрезвычайно популярно в народной медицине и в его местообитаниях населением интенсивно собирается растительный материал для выращивания в садовых това- риществах и приусадебных хозяйствах. Тем самым наносится угроза целостности и устойчивости популяционной структуры девясила высокого. Кроме того, получаемая при исследовании информация может стать важной для научно обоснованной интродукции при создании плантаций для выращивание лекарственного сырья.
Цель исследования – изучение популяционной структуры и дифференциации популяций девясила высокого, обсуждение полученных результатов в сравнении с данными по другим ранее исследованных видам лекарственных растений Южного Урала в свете проблем сохранения их генофонда in situ и ex situ.
Материалы и методы исследований
Для изучения отобраны выборки из 9 природных популяций девясила высокого. Они расположены в южных районах Республики Башкортостан на территории Зианчуринско-го (пробные площади Zia-1 и Zia-2), Куюрга-зинского (Kum-1, Kum-2), Мелеузовского (Mel-1, Mel-2), Кугарчинского (Kug) и Ишибайско-го (Ish-1, Ish-2) районов в лесостепи Зилаир-ского плато и предгорий западного макросклона Южного Урала.
В качестве маркеров для характеристики популяционных структур нами использованы изоферменты. Для их анализа использован полиакриламидный гель-электрофорез [5, 10]. Гистохимическое окрашивание фермен- тов в гелях проведено по стандартным методикам [1]. Ферменты и локусы обозначались по одной из общепринятых номенклатур [11]. Более полное описание методов экстракции изоферментов и электрофореза было приведено ранее [2].
Для вычисления популяционных параметров использованы показатели, определяемые компьютерной программой BIOSYS-1 [12] – частота аллелей, доля полиморфных локусов Р , среднее число аллелей на локус А наблюдаемая и ожидаемая гетерозиготность ( H O и HE ,соответственно), показатель уровня подразделенности популяций F ST , [9], а также коэффициент инбридинга (индекс фиксации Райта) и генетическое расстояние М. Нея D [8]. Формулы их расчета, а также определения степени гетерогенности выборок были приведены ранее [2].
Для изучения уровня дифференциации популяций особенно информативными являются полиморфные гены. В результате проведенного скрининга электрофоретической изменчивости ряда изоферментных систем установлено, что к поливариантным могут быть отнесены аллели 4 локусов. Они кодируют синтез неспецифических эстераз (EST, E.C. 3.1.1.1…), лейцинаминопептидазы (LAP, E.C. 3.4.11.1.), аспартатаминотрансферазы (AAT, E.C. 2.6.1.1.) и алкогольдегидрогеназы (ADH, E.C. 1.1.1.1.). Гипотезы о механизмах наследования ферментов были выдвинуты в ходе анализа электрофореграмм (пространственной разобщенности зон активности в гелях, образования гибридных молекул у гетерозигот и др.) и статистическим тестировании различий теоретически ожидаемых и наблюдаемых частот фенотипов изоферментов.
Результаты и их обсуждение
При сравнении частот наблюдаемых и теоретически ожидаемых распределений генотипов установлено, что их соотношение в большинстве популяций хорошо сбалансировано. Правило Харди-Вайнберга нарушается только в одном из 36 тестированных вариантов (2,8%). Этот прецедент отмечен в выборке Kum-1 из-за нехватки генотипов Aat-
11/2 ( χ 2 =10,2, статистически достоверно на уровне значимости Р < 0,01).
Частоты аллелей изученных четырех локусов приведены в таблице 1. В большинстве популяций в каждом локусе доминирует один из изоферментов. Исключениями являются Est-13 в выборках Kug-1 и Mel-2, Aat-11 в Mel-2, Adh-11 в Kum-2.
Среднее генетическое расстояние Нея, показывающее сходство и различия популяций по частотам аллелей, составило значение D= 0,081 ± 0,011 с изменениями от 0,005 до 0,275. Сходство генофондов по принципу «изоляции расстоянием» показано лишь для пары Zia-1/ Zia-2, разделенных расстоянием в 10 км (D = 0,005). У находящихся друг от друга на практически таком же удалении выборок Ish-1 и Ish-2 (13 км) генетическое расстояние больше на один порядок (D= 0,071). Различия частот аллелей довольно высоки и у пары Kum-1/ Kum-2 из одного района (D = 0,040, 14 км). У популяций Ish-2 и Zia-1, географически наиболее разобщенных (189 км), различия генофонда выражены сравнительно немного (D = 0,045). Корреляционный анализ также свидетельствует об отсутствии статистически достоверной связи между генетическим и географическим расстояниями. Более наглядно отсутствие пространственных закономерностей изменчивости изоферментов показаны на дендрограмме (рис.). На ней сформирована группа кластеров с небольшим [7] уровнем различий D = 0,060. В пределах этой группы какие-либо географические закономерности в пространственном расположении выборок практически не наблюдаются. От нее серьезно (D = 0,189) отделена выборка Mel-2. Этот феномен может быть вызван особенностями местообитания. Все остальные пробные площади представлены большими зарослями девясила высокого на влажных лугах, где в среднем на один квадратный метр приходится 0,71 ± 0,23 растение. Участок Mel-1 расположен в густом ольшат-нике вдоль реки и включает небольшие, удаленные друг от друга группы особей, со средней плотностью 0.04 особи на один квадратный метр. В таких условиях из-за их изоляции и затруднения переопыления растений может появиться «эффект основателя», приводящий к резкому изменению генотипического состава. Другим последствием скрещивания только в пределах изолированной группы особей может быть инбридинг. В подтверждение этого предположения на пробной площади Mel-2 коэффициент инбридинга F = +0,187 – ощущается экцесс гомозиготности (табл. 2). В другом местообитании с относительно небольшой численностью растений на единицу площади (0,18 шт.) также выявлен инбридинг (Kum-1, F = +0,232). На остальных местообитаниях, где на один квадратный метр приходится 0,45-2,25 растений, коэффициент близок к нулю или наблюдается экцесс гетерозиготности.
Заключение
Редкие и исчезающие виды растений составляют наиболее чувствительную часть биоразнообразия, подлежащего охране. Причиной этого является уязвимость генофонда этой категории организмов из-за ограничения потока и дрейфа генов [6]. Из их числа лекарственные растения находятся в более опасном положении, так как деградация их популяций происходит не только по природным, но и по антропогенным причинам. Примером может служить наше исследование по родиоле иремельской («золотому корню», эндемику Южного и Среднего Урала) [3]. Из-за хищнических нерегулируемых сборов растения населением более чем на один порядок снизилась численность растений в горном массиве Иремель, исчезли целые субпопуляции. Это привело с 1990 по 2001 г.г. к утере части аллелей, понижению ожидаемой и наблюдаемой гетерозиготности от 0.257 до 0,182 и от 0,288 до 0,221 для HOиHE ,соот-ветственно.
Таблица 1. Частоты аллелей полиморфных локусов девясила высокого
|
Аллели |
Частоты аллелей в выборках |
||||||||
|
Zia-1 |
Zia-2 |
Kug-1 |
Kum-1 |
Kum-2 |
Mel-1 |
Mel-2 |
Ish-1 |
Ish-2 |
|
|
Est-1 1 |
0.125 |
0.206 |
0.500 |
0.432 |
0.266 |
0.219 |
0.563 |
0.297 |
0.016 |
|
Est-1 2 |
0 |
0 |
0.028 |
0.023 |
0.031 |
0.016 |
0.016 |
0.016 |
0 |
|
Est-1 3 |
0.850 |
0.794 |
0.472 |
0.545 |
0.688 |
0.734 |
0.422 |
0.656 |
0.968 |
|
Est-1 4 |
0.025 |
0 |
0 |
0 |
0.016 |
0.031 |
0 |
0.031 |
0.016 |
|
Lap-1 1 |
1.000 |
1.000 |
1.000 |
0.727 |
0.813 |
0.563 |
0.922 |
0.703 |
0.806 |
|
Lap-1 2 |
0 |
0 |
0 |
0.273 |
0.188 |
0.438 |
0.078 |
0.297 |
0.194 |
|
Aat-1 1 |
1.000 |
0.882 |
0.889 |
0.750 |
0.953 |
0.953 |
0.344 |
0.969 |
0.710 |
|
Aat-1 2 |
0 |
0.118 |
0.111 |
0.250 |
0.047 |
0.047 |
0.656 |
0.031 |
0.290 |
|
Adh-1 1 |
0.275 |
0.294 |
0.306 |
0.341 |
0.516 |
0.313 |
0.250 |
0.344 |
0.177 |
|
Adh-1 2 |
0.725 |
0.707 |
0.694 |
0.659 |
0.484 |
0.688 |
0.750 |
0.656 |
0.823 |
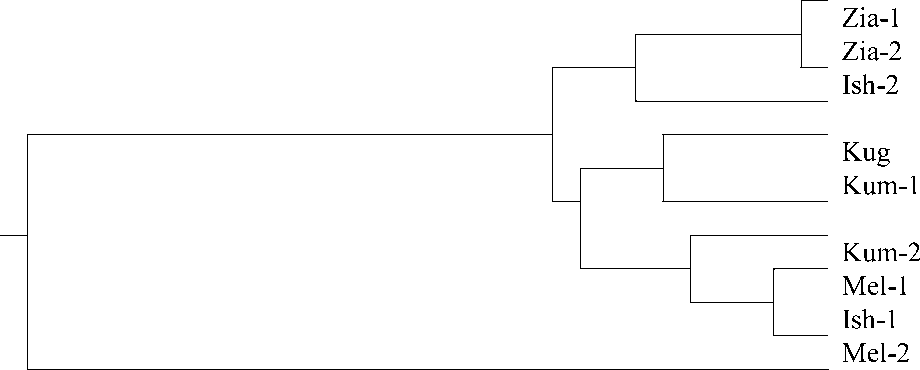
Рис. Дендрограмма, построенная на основе генетических расстояний между популяциями девясила высокого
Субпопуляционная подразделенность даже в близко расположенных местообитаниях возросла от F ST = 0,033 до F ST = 0,073, генетическое расстояние Нея увеличилось с D = 0,010 до D = 0,033 до Средний для локусов коэффициент инбридинга изменился с F = 0,108 до F = + 0.176. Для родиолы иремельской показано [3], что сохранение ее генофонда возможно только методами ex situ. Фрагментация популяций, снижение численности растений в отдельных местообитаниях, генетическая дифференциация и снижение полиморфизма достигли такого уровня, что популяции не способны воспроизводить свое разнообразие в природных условиях. Рекомендовано создание синтетических популяций
Судя по полученным нами данным, генофонд девясила высокого на юге Башкортостана находится в более благополучном положении. Состав генотипов в большинстве местообитаний является хорошо сбалансированным, фрагментация частот аллелей не выражена – основная часть изменчивости приходится на долю внупопуляционной компоненты и свидетельствует о сравнительно интенсивном генетическом потоке. При исключении из анализа выборки Mel-2, значительно отличающейся от других групп изученных растений по частотам аллелей, генетическое расстояние между популяциями уменьшается почти в 1,7 (D = 0,049 ± 0,005). Параметр межвыборочной подразделенности составляет при этом FST = 0,094 – 90,06% изменчивости является внутрипопуляцион-ной. Эти оценки близки к данным по широко распространенным растениям (характеризующимся большими ареалами) со свойствами, которыми обладает и девясил высокий. В среднем у таких видов лесной и лесостепной зоны FST = 0,038, покрытосеменных растений FST = 0,073, насекомоопыляемых видов FST = 0,099 [7]. Влияние дрейфа генов не прослеживается. Этот микроэволюционный фактор стремится к фиксации аллелей (доведению их частот до 0 или 1.000, в первую очередь за счет редких вариантов) и снижает генетическое разнообразие. Однако в изученных нами полиморфных локусах альтернативные аллели имеют сопоставимые частоты, гетерозиготность является сравнительно высокой, обнаружены редкие аллели. Видимо, из этих обстоятельств и нужно исходить при вынесении рекомендаций по сохранению генофонда девясила высокого методами in situ и ex situ. Здесь требуют ответа следующие главные вопросы: 1) какие популяции требуют сохранения; 2) достаточно ли их сохранения в природных условиях; 3) не нужно ли сохранять генофонд вида также ex situ; 4) каким образом отбирать популяций для сохранения ex situ?
Таблица 2. Изменчивость полиморфных локусов девясила высокого
|
Параметры |
Выборки |
||||||||
|
Zia-1 |
Zia-2 |
Kug-1 |
Kum-1 |
Kum-2 |
Mel-1 |
Mel-2 |
Ish-1 |
Ish-2 |
|
|
Р , % |
50 |
75 |
75 |
100 |
100 |
100 |
100 |
100 |
100 |
|
А |
1.8 (0.5) |
1.8 (0.3) |
2.0 (0.4) |
2.3 (0.3) |
2.5 (0.5) |
2.5 (0.5) |
2.3 (0.3) |
2.5 (0.5) |
2.3 (0.3) |
|
НЕ |
0.169 (0.102) |
0.245 (0.093) |
0.295 (0.121) |
0.444 (0.032) |
0.343 (0.094) |
0.361 (0.092) |
0.375 (0.081) |
0.358 (0.100) |
0.274 (0.075) |
|
НО |
0.213 (0.133) |
0.279 (0.106) |
0.278 (0.138) |
0.341 (0.132) |
0.344 (0.095) |
0.367 (0.099) |
0.305 (0.087) |
0.367 (0.107) |
0.266 (0.073) |
|
F |
-0.260 |
-0.139 |
+0.058 |
+0.232 |
-0.003 |
-0.017 |
+0.187 |
-0.025 |
+0.029 |
Примечание: в скобках приведены ошибки средних арифметических значений.
При межпопуляционной подразделенности до уровня FST = 0,200 для сохранения 95 % изменчивости достаточно использование всего лишь двух популяций [6]. У девясила высокого, с его более слабой дифференциацией популяций, в больших местообитаниях возможно сохранение довольно репрезентативной части генофонда вида (по крайней мере, для исследованной части ареала). Выбор расположения объектов охраны in situ и их формы при этом мало ограничивается генетическими причинами. Это обстоятель- ство дает возможность большего учета других (экономических, организационных и т.д.) факторов. Это заключение, видимо, справедливо и при отборе растений для сохранения генофонда и их использования в условиях ex situ. Следует исключить создание особо охраняемых природных территорий и формирование коллекции для интродукции на базе местообитаний с нетипичной популяционной структурой растений. Как показали результаты настоящего исследования, в таких условиях формируется генофонд, в целом не
Список литературы Изучение популяционной структуры лекарственных растений как основы разработки мер по сохранению генофонда
- Корочкин Л.И., Серов О.Л., Пудовкин А.И. и др. Генетика изоферментов. М.: Наука, 1977.
- Межпопуляционная дифференциация родиолы иремельской (Rhodiola iremelica Boriss., Grassulaceae) на Южном Урале/Янбаев Ю.А., Байрамгулов Н.Р., Редькина Н.Н., Муллагулов Р.Ю.//Генетика. 2007. Т. 43. № 11.
- Популяционная структура и принципы сохранения генофонда родиолы иремельской на Южном Урале. Уфа: БашГУ, 2000.
- Розенберг Г.С., Краснощеков Г.П., Крылов Ю.М. и др. Устойчивое развитие: мифы и реальность. Тольятти: ИЭВБ, 1998.
- Davis B.J. Disc electrophoresis. 11. Methods and application to human serum proteins//Ann. New York Acad. Sci. 1964. Vol. 121. № 2.
- Godt M.J.W., Hamrick J.L. Genetic diversity and population structure in Tradescantia hirsuticaulis (Commmelinaceae)//Am. J. Bot. 1993. V. 80.
- Hamrick J.L., Godt M.J.W., Sherman-Broylers S.L. Factors influencing levels of genetic diversity in woody plant species//New Forest. 1992. №. 6.
- Nei M. Genetic distance between populations//Amer. Natur. 1972. V. 106. №. 949.
- Nei M. F-statistics and analysis of gene diversity in subdivided populations//Ann. Hum. Genet. 1977. V. 41.
- Ornstein L. Disc-electrophoresis. I. Background and theory//Ann. New York. Acad. Sci. 1964. Vol. 121.
- Prakash S., Lewontin R.C., Hubby J.L. A molecular approach to the study of genetic heterozygosity in natural populations. IV. Patterns of genic variation in central, marginal and isolated populations of Drosophila pseudoobscura//Genetics. 1969. V. 61.
- Swofford D.L., Selander R.B. BIOSYS-1: a FORTRAN program for the comprehensive analysis of electrophoresis data in population genetics and systematics//J. Heredity. 1981. V. 72.
