Изучение потенциал зависимых ионных каналов биологических мембран

Автор: Зарипов А.А., Рахметуллаев П.М.
Журнал: Экономика и социум @ekonomika-socium
Рубрика: Основной раздел
Статья в выпуске: 6-1 (37), 2017 года.
Бесплатный доступ
В статье рассматриваются вопросы изучения потенциал зависимых ионных каналов биологических мембран под воздействием биологически активных соединений.
Клеточная мембрана, ионные каналы, лиганд-активируемые каналы
Короткий адрес: https://sciup.org/140124308
IDR: 140124308
Study of the potential of dependent ionic channels of biological membranes
The article examines the potential of dependent ion channels of biological membranes under the influence of biologically active compounds.
Текст научной статьи Изучение потенциал зависимых ионных каналов биологических мембран
Zaripov Abdisalim Abdikarimovich
Assistant of the Department of Biology
The Karakalpak State University of. Berdaha
Rakhmetullaev Pirimzhar Mukhtarovich Second-year student in the specialty "Biology"
The Karakalpak State University of. Berdaha
Nukus, Republic of Uzbekistan
STUDY OF THE POTENTIAL OF DEPENDENT IONIC CHANNELS OF BIOLOGICAL MEMBRANES
Ионные каналы – сложнейшие и высоко консервативные белковые структуры, которые могут управляться физическими воздействиями, такими как трансмембранная разность электрических потенциалов или механическое растяжение мембраны, либо химическими сигналами - лигандами. Высокая сложность позволяет точно распознавать ионы и пропускать их через мембрану управляемым образом. Ионные каналы высокоселективны; они пропускают ионы только определенного вида: Na+, K+, CaІ+ или Cl─. Их, соответственно, называют натриевыми, калиевыми, кальциевыми или хлорными каналами. Каналы для каждого вида ионов далее классифицируются по их структуре, пороговым характеристикам, кинетике, чувствительности к различным лигандам и т.д.
Основные части ионного канала: устья - сужающиеся воронки, обращенные в среду или цитоплазму, куда входят или откуда выходят ионы; селективный фильтр - наиболее узкая часть канала, позволяющая проходить только ионам определенного вида; и ворота - воротный механизм, открывающий или закрывающий канал (Рис.1). В потенциал зависимых ионных каналах должен существовать потенциал чувствительный сенсор, реагирующий на изменения трансмембранного потенциала и управляющий состоянием ворот.
В устье расположены отрицательно заряженные карбоксильные группы (СОО─). Они притягивают из раствора положительно заряженные катионы, например, Na+ или К+. Это создает повышенную концентрацию катионов у устья канала. Это место является первой потенциальной ямой в канале: катион, связанный отрицательными группами в устье имеет минимальную потенциальную энергию. В состав селективного фильтра входит еще одна или несколько групп СОО─, образующие вторую или третью потенциальные ямы. Потенциальный профиль канала изображен на рис.1. Проходя по каналу, положительный катион связывается с этой отрицательно заряженной группой и попадает в потенциальную яму. Для дальнейшего прохождения по каналу и выхода из него он должен преодолеть это электростатическое притяжение.
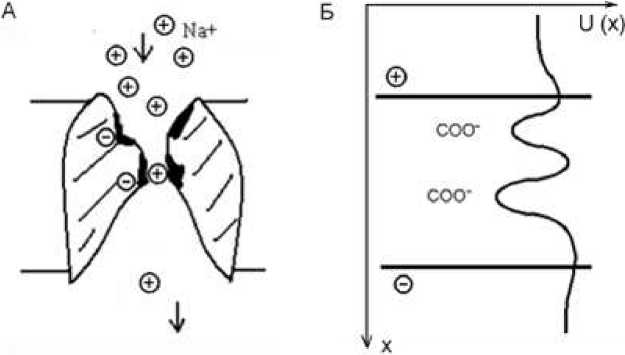
Рис.1. Схема строения ионного канала (А) и распределение в нем потенциальной энергии катиона (Б)
Первичным источником энергии в клетке является, разумеется, химическая энергия, запасенная в молекулах АТФ. За ее счет (Na+-K+)-АТФаза создает электрохимический градиент ионов натрия и калия, а Ca2+-АТФаза - ионов кальция, на плазматической мембране. Ионы двигаются по каналу за счет энергии, запасенной в этих градиентах. Отрицательные заряды во входном устье канала собирают большее катионное облако, чем у выходного устья. Эти ионы создают электростатическое поле, проталкивающее катионы в канал. Если в первом центре связывания есть ион, он будет выталкиваться, как биллиардный шар, во вторую яму следующим подлетающим ионом. Так как благодаря повышенной концентрации число таких ударов на входе в канал больше, чем на выходе, то ионы будут двигаться по каналу по градиенту концентрации. Если вторая яма занята, то ион, вытолкнутый из первой ямы, будет выталкивать второй ион, чтобы занять его место. То есть, одномерная диффузия ионов по каналу осуществляется путем прыжков ионов между потенциальными ямами (отрицательно заряженными группами СОО-).
К группе управляемых каналов относятся также механочувствительные ИК, которые повышают свою проницаемость в ответ на механические воздействия на клеточную мембрану. Сенсорные рецепторы растяжения, содержащие в своей мембране такие ИК, найдены, например, в механорецепторах кожи. Такие ИК мы предлагаем называть «стимул-управляемыми». Другие ИК открываются тогда, когда определённые химические вещества (их обобщённое название - «лиганды») активируют связывающие центры на молекуле канала. Такие лиганд -активируемые ИК подразделяются на две подгруппы, в зависимости от того, являются ли активные центры внутриклеточными или внеклеточными. Каналом, отвечающим на внеклеточную активацию, является, например, катионный канал постсинаптической мембраны в скелетной мышце. Этот канал активируется нейротрансмиттером ацетилхолином, освобождающимся из двигательного нервного окончания. Открытие ацетилхолин-активируемого ионного канала позволяет ионам натрия войти в клетку, вызывая деполяризацию мышечного волокна. Такие ИК мы предлагаем называть «лиганд-управляемыми». Лиганд-активируемые каналы, отвечающие на внутриклеточные стимулы, включают в себя каналы, чувствительные к местным изменениям концентрации специфических ионов. Например, кальций-активируемые калиевые каналы активируются локальным повышением концентрации внутриклеточного кальция. Такие каналы играют важную роль в реполяризации клеточной мембраны во время завершения потенциала действия.
Помимо ионов кальция, типичными представителями лигандов, активирующих ИК с цитоплазматической стороны мембраны, являются циклические нуклеотиды. Циклический ГМФ, например, отвечает за активацию натриевых каналов в палочках сетчатки. Такой тип канала играет принципиальную роль в работе зрительного анализатора. Такие ИК мы предлагаем называть «опосредованно-управляемыми» ( «мессенджер-управляемыми»). Предлагаемая классификация ИК по способу управления их состоянием не является достаточно строгой, поскольку выделяемые нами группы не являются взаимоисключающими, и каналы могут одновременно входить в несколько групп. Например, кальций- активируемые
калиевые
каналы (опосредованно-управляемые)
чувствительны также и к изменению потенциала (т.е. являются одновременно и потенциал-управляемыми или же совместно-управляемыми), а некоторые потенциал-управляемые ИК чувствительны к внутриклеточным лигандам, т.е являются дополнительно опосредованно-управляемыми.
ИК различаются по своей избирательности (селективности). Так, некоторые катионные каналы пропускают в основном только один вид ионов: натрий, калий или кальций, другие же являются менее избирательными. Анионные каналы сравнительно не избирательны для малых анионов, но они пропускают в основном ионы хлора, так как хлор является самым распространенным анионом в составе внеклеточной и внутриклеточной жидкости в организме.
Список литературы Изучение потенциал зависимых ионных каналов биологических мембран
- Зефиров А.Л., Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология)//Казань: Арт-кафе, 2010. 271 с.
- Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология//Учебное пособие для студентов медицинских вузов. М.: ООО "Медицинское информационное агентство", 2003. 544 с.
- Сазонов В.Ф. Функциональная классификация мембранных ионных каналов//Научные труды III Съезда физиологов СНГ/Под ред. А.И. Григорьева, О.А. Крышталя, Ю.В. Наточина, Р.И. Сепиашвили. М.: Медицина-Здоровье, 2011. С. 72. (Электронная версия: physiology-cis.org/Page181.html)
- Фундаментальная и клиническая физиология. Под ред. А.Г. Камкина и А.А. Каменского. М.: Академия, 2004. 1072 с.
