Изучение терморегуляционного поведения рептилий при подкожной имплантации терморегистраторов

Автор: Четанов Николай Анатольевич, Литвинов Николай Антонович, Ганщук Светлана Владимировна, Галиулин Данила Минуллович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 2, 2022 года.
Бесплатный доступ
Сделана попытка охарактеризовать терморегуляционное поведение рептилий без непосредственного наблюдения за ними при помощи сравнения данных двух терморегистраторов: вшитого подкожно объекту исследования и размещенному в биотопе. Выделены основные затруднения, возникающие при проведении подобных исследований, определены важнейшие условия для успешной реализации. На основании анализа собственных данных выделены и описаны пять основных форм терморегуляционного поведения у рептилий.
Терморегуляционное поведение, рептилии, подкожная имплантация, ibdl ds1922l
Короткий адрес: https://sciup.org/147238650
IDR: 147238650 | УДК: 591.543.1+59.087 | DOI: 10.17072/1994-9952-2022-2-119-124
The study of the thermoregulatory behavior of reptiles during subcutaneous implantation of thermorecorders
The article presents an attempt to characterize the thermoregulatory behavior of reptiles without direct observation of them by comparing the data of two thermal recorders: one sewn subcutaneously to the object of study and one placed in the biotope. The main difficulties that arise in the course of such studies are identified, and the most important conditions for successful implementation are identified. Based on the analysis of our own data, five main forms of thermoregulatory behavior in reptiles are identified and described.
Текст научной статьи Изучение терморегуляционного поведения рептилий при подкожной имплантации терморегистраторов
Первая работа, посвященная изучению термобиологии рептилий при помощи имплантации логгеров-терморегистраторов, появилась в нашей стране еще десять лет назад [Литвинов, Ганщук, 2012]. С тех пор логгеры периодически применялись исследователями при изучении вопросов термобиологии рептилий [Литвинов, Четанов, 2014; Берман и др., 2020].
В последние годы вышел целый ряд работ, посвященных изучению поведенческой терморегуляции рептилий при помощи имплантации логгеров [Литвинов и др., 2016; Ганюшина и др., 2019; Коросов и др., 2021, Ганюшина, Коросов, 2021].
Однако существует базовая проблема – нет возможности оценить корректность полученных с помощью терморегистраторов данных, если при этом не проводилось прямое наблюдение за формами активности рептилии. Что логично, так как зачастую при изучении в природных условиях это просто невозможно.
Мы поставили перед собой цель – разработать подход, позволяющий судить о терморегуляционном поведении рептилий, опираясь на данные всего лишь двух терморегистраторов – одного, вшитого подкожно, и второго, измеряющего температуру окружающей среды – без параллельной фиксации поведенческих актов. Данное сообщение по сути является предварительным, скорее всего, представляющим общую концепцию.
Материалы и методы исследования
В работе нами используются четыре относительно просто определяемые величины, для вычисления которых требуются показания всего лишь двух логгеров-терморегистраторов:
-
1) температура тела (T t );
-
2) температура окружающей среды (T s );
-
3) изменение температуры тела (ΔT t );
-
4) изменение температуры среды (ΔT s ).
Рассмотрим каждый из них. Начнем с температуры тела и температуры окружающей среды.
Размеры датчика накладывают ограничения на возможность проведения имплантации: объект должен быть достаточно крупным. Наши попытки повторить процедуру по миниатюризации логгеров [Robert, Thompson, 2003; Lovegrove, 2009] можно считать неудачными. Размер логгера при этом практически не уменьшается, но в значительной мере возрастает шанс на выход терморегистратора из строя и повреждение данных.
Также серьезной сложностью является повторный отлов животного для извлечения вшитого регистратора. В связи с этим для изучения можно рекомендовать только виды, имеющие выраженную территориальность, в противном случае шанс на получение данных с логгера крайне невелик.
Учитывая изложенные ограничения, из шести видов рептилий, достоверно обитающих на территории Камского Предуралья, по всей видимости, лучше всего подходит для проведения подобного исследования обыкновенная гадюка Vipera berus (Linnaeus, 1758). Прыткая ящерица Lacerta agilis Linnaeus, 1758 и живородящая ящерица Zootoca vivipara (Lichtenstein, 1823) не подходят по размерам тела. Теоретически можно проводить экспериментальную работу с обыкновенным ужом Natrix natrix (Linnaeus, 1758), обыкновенной медянкой Coronella austriaca Laurenti, 1768 и даже колхидской веретеницей Anguis colchica (Nordmann, 1840), однако только при использовании какого-то вольера, в котором в течение всего периода наблюдений и будут содержаться объекты со вшитыми терморегистраторами. В принципе, первые опыты по имплантации и проводились с обыкновенным и водяным ужами, содержащимися в специально оборудованном вольере [Литвинов, Ганщук, 2002].
В качестве температуры окружающей среды (T s ) нами принималась температура приземного воздуха, примерно на высоте 2–4 см от поверхности субстрата. Температура фиксировалась при помощи логгера-терморегистратора, размещенного в характерном для вида биотопе, желательно в том же месте, где проводился отлов животного. Обязательным условием является размещение логгера в затененной области (например, среди растений), чтобы избежать прямого нагрева солнечными лучами, что может привести к неверной оценке температурных условий.
Для корректности получаемых данных необходима синхронизация логгеров, измеряющих температуру тела и окружающей среды, с персональным компьютером, а также одновременный их запуск и одинаковые интервалы отсчетов измерений.
Результаты исследований
Изменение температуры тела (ΔT t ) может быть положительным или отрицательным, то есть животное может или нагреваться (ΔT t > 0), или остывать (ΔT t < 0). О причинах изменения температуры говорить трудно, но, так как эндотермия для современных рептилий недоступна, можно сделать обоснованное предположение, что все эти изменения связаны с какими-то внешним факторами: или температурой окружающей среды, или солнечной радиацией. Причем благодаря использованию энергии Солнца рептилии способны длительное время поддерживать стабильную температуру [Saint-Girons, 1975].
Изменение температуры окружающей среды (ΔT s ) в значительной мере предсказуемо: в утренние часы наблюдается повышение температуры (ΔT s > 0), в вечерние – понижение (ΔT s < 0). Однако нельзя забывать о случайных изменениях температуры в течение суток (облачность, осадки). Соответственно, подобные отклонения должны или учитываться при анализе данных, или отбраковываться из общего массива.
Какие, на наш взгляд, возможны варианты сочетаний температурных параметров и как они связаны с формами терморегуляционного поведения? Общие идеи отражены в представленной ниже таблице. Сразу же уточним, выделение в данном случае лишь пяти форм терморегуляционного поведения не охватывает весь спектр поведенческой терморегуляции у рептилий, который, несомненно, значительно шире [Черлин, 2010, 2012], однако выделение прочих форм без непосредственного наблюдения за объектом, по всей видимости, невозможно.
Характеристика основных форм терморегуляционного поведения рептилий [Characteristics of the main forms of thermoregulatory behavior of reptiles]
|
Форма поведения |
Характерные признаки |
||
|
ΔT t |
ΔT s |
Соотношение температур тела и среды |
|
|
Активное нагревание (АН) |
> 0 |
> 0 |
ΔT t > ΔT s ; T t > T s |
|
Пассивное нагревание (ПН) |
> 0 |
> 0 |
ΔT t < ΔT s ; T t < T s или ΔT t > ΔT s ; T t < T s |
|
Вечернее нагревание (ВН) |
> 0 |
< 0 |
ΔT t > ΔT s |
|
Активное остывание (АО) |
< 0 |
> 0 |
ΔT t < ΔT s |
|
Пассивное остывание (ПО) |
< 0 |
< 0 |
ΔT t < ΔT s |
Постараемся охарактеризовать каждую из этих форм терморегуляционного поведения подробнее и проиллюстрируем на примере данных за одни «типичные» сутки, полученных при изучении термобиологии обыкновенного ужа при вольерном содержании (рисунок).
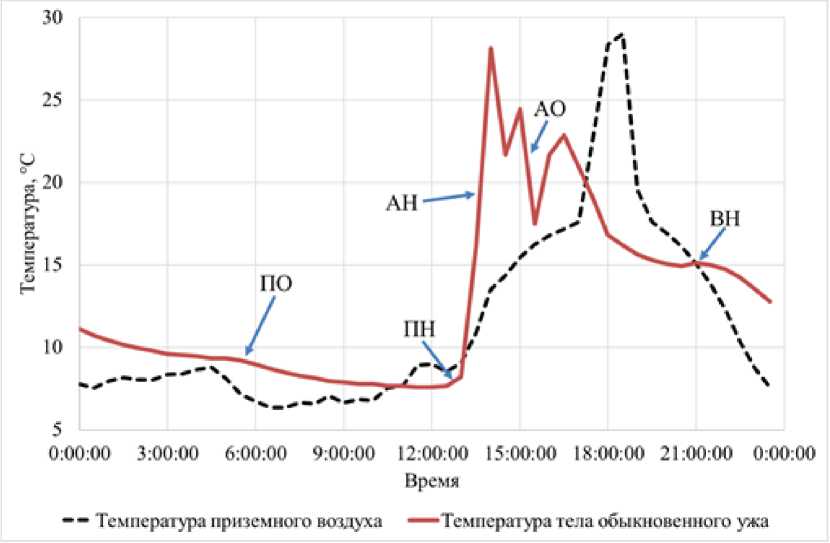
Суточная динамика температуры приземного воздуха и температуры тела обыкновенного ужа [Daily dynamics of surface air temperature and body temperature of common grass snake]
Как видно из таблицы, по нашему мнению, можно выделить минимум три варианта нагревания: активное нагревание, вечернее нагревание, а также пассивное нагревание, причем при активном и пассивном обогреве изменения температуры тела и среды однонаправленное.
При активном нагревании можно наблюдать повышение как температуры тела ужа, так и температуры окружающей среды, однако скорость роста температуры окружающей среды значительно ниже. В подавляющем большинстве случаев подобная ситуация будет наблюдаться в утренние часы, когда рептилия повышает температуру своего тела за счет инсоляции. Это, по всей видимости, позволяет животному за короткий промежуток времени достичь оптимальной температуры тела, более высокой, чем температура среды обитания. Соответственно, окончанием данной формы поведения может служить выравнивание температур или скоростей нагрева.
На рисунке этот вариант отмечен как АН.
При пассивном нагревании возможны два основных варианта.
В первом случае температура животного пассивно повышается совместно с повышающейся температурой среды, причем скорость роста температуры среды выше, чем у животного. Тогда, по всей видимости, речь идет о постепенном нагревании в тени.
Второй вариант, когда температура тела рептилии повышается быстрее, чем температура окружающей среды, значительно сложнее идентифицировать и отделить от активного нагревания. На наш взгляд, подобное возможно, если рептилия покидает убежище с низкими температурами. Единственным относительно надежным маркером в таком случае может служить значительно более низкая температура тела рептилии по сравнению с окружающей средой.
На рисунке этот вариант отмечен как ПН.
Гораздо легче выделить вечернее нагревание – при снижении температуры окружающей среды наблюдается, пусть и непродолжительное, но повышение температуры тела. Нами в природных условиях неоднократно наблюдались рептилии, которые перед уходом в ночное убежище грелись под последними лучами заходящего солнца, при этом их температура тела при отлове была зачастую значительно выше, чем температура окружающей среды.
На рисунке этот вариант отмечен как ВН.
Для остывания, на наш взгляд, можно выделить только две основные формы поведения – активное и пассивное.
При активном остывании наблюдается снижение температуры тела при повышении температуры окружающей среды. Возможны два варианта объяснения подобного явления:
-
1) если температура среды выше, чем температура тела, то, скорее всего, произошел уход животного в некую холодную зону в связи с превышением температурой среды комфортных для рептилии условий. Это может быть расселина между камней, нора, какое-то сильно затененное место;
-
2) если температура окружающей среды ниже, чем температура тела, но последняя при этом все равно снижается, то, по всей видимости, происходит уход из освещаемой, хорошо прогретой зоны, которую рептилия использовала для активного нагревания.
На рисунке этот вариант отмечен как АО.
Пассивное остывание в этом плане значительно менее интересно – температура тела снижается вслед за снижением температуры окружающей среды, однако с некоторым запаздыванием. Скорее всего, это связано с пребыванием в ночном убежище. По сути, полноценной формой поведения считать пассивное остывание сложно, так как активность животного при этом близка к нулю.
На рисунке этот вариант отмечен как ПО.
Таким образом, при проведении анализа данных, полученных всего лишь с двух логгеров, можно изучить достаточно подробную характеристику терморегуляционного поведения рептилий. Но подобный подход без прямых наблюдений за объектом не позволяет судить о причинах изменения температуры и, естественно, не отражает всего спектра терморегуляционного поведения у рептилий
Заключение
Постараемся подвести предварительные итоги:
-
1) для проведения исследований терморегуляционного поведения с помощью имплантированных лог-геров-терморегистраторов необходим достаточно крупный объект с выраженной территориальностью. Из видов, обитающих на территории Камского Предуралья, лучше всего подходит обыкновенная гадюка;
-
2) миниатюризация логгеров не является обязательным условием для изучения термобиологии рептилий;
-
3) анализируя информацию, полученную с двух логгеров (имплантированного подкожно и размещенного в биотопе), можно выявить основные аспекты терморегуляционного поведения без проведения прямых наблюдений за животным;
-
4) выявленные при помощи данного подхода формы терморегуляционного поведения рептилий можно условно разделить на пять видов: активное нагревание, пассивное нагревание, вечернее нагревание, активное остывание и пассивное остывание. Отнесение пассивного остывания к терморегуляционному поведению достаточно спорно. Более точное описание форм поведения без непосредственных наблюдений за объектом, по всей видимости, невозможно.
Список литературы Изучение терморегуляционного поведения рептилий при подкожной имплантации терморегистраторов
- Берман Д.И. и др. Холодостойкость и зимовка обыкновенной гадюки (Vipera berus, Reptilia, Viperidae) на о-ве Кижи, Карелия // Зоологический журнал. 2020. Т. 99, вып. 8. С. 10-30.
- Ганюшина Н.Д., Коросов А.В. Изменчивость параметров терморегуляции обыкновенной гадюки // Труды Зоологического института РАН. 2021. Т. 325, № 1. С. 99-112.
- Ганюшина Н.Д. и др. Наружная и внутренняя температуры тела обыкновенной гадюки // Известия высших учебных заведений. Поволжский регион. Естественные науки. 2019. № 2 (26). С. 17-27. DOI: 10.21685/2307-9150-2019-2-2.
- Коросов А.В. и др. Параметры терморегуляции обыкновенной гадюки (Vipera berus L.) в разных частях ареала // Принципы экологии. 2021. № 3. С. 54-63.
- Литвинов Н.А., Ганщук С.В. Температура тела обыкновенного ужа Natrix Natrix (Linnaeus, 1758) в Волжском бассейне в естественной среде и эксперименте // Известия Самарского научного центра РАН. 2012. № 1-1. С. 154-157.
- Литвинов Н.А., Панова М.К., Окулов Г.А. Изучение терморегулирующего поведения рептилий методом имплантации регистраторов температуры // Вестник Санкт-Петербургского университета. Сер. 3. Биология. 2016. Вып. 3. С. 69-76. DOI: 10.21638/11701/spbu03.2016.313
- Литвинов Н.А., Четанов Н.А. Непрерывная регистрация температуры тела обыкновенной гадюки летом // Известия Самарского научного центра РАН. 2014. Т. 16, № 5. С. 430-435.
- Черлин В.А. Термобиология рептилий. Общая концепция. СПб.: Русско-Балтийский информационный центр БЛИЦ, 2012. 362 с.
- Черлин В.А. Термобиология рептилий. Общие сведения и методы исследований (руководство). СПб.: Русско-Балтийский информационный центр БЛИЦ, 2010. 124 с.
- Lovegrove, B.G. Modification and miniaturization of Thermochron iButtons for surgical implantation into small animals // J. Comp. Physiol. B. 2009. Vol. 179. P. 451-458. DOI: 10.1007/s00360-008-0329-x.
- Robert K.A., Thompson M.B. Reconstructing Thermochron iButtons to reduce size and weight as a new technique in the study of small animal thermal biology // Herpetol. Rev. 2003. Vol. 34. P. 130-132.
- Saint-Girons H. Observations préliminaires sur la thermorégulation des Vipères d'Europe // Vie et Milieu. Ser. C. 1975. Vol. 25. P. 137-168.
