Изучение возможного участия лектинов Azospirillum в механизмах адаптивного ответа растений, подвергшихся воздействию солей тяжелых металлов

Автор: Аленкина С.А., Рощупкина К.Д., Никитина В.Е.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 1 т.15, 2019 года.
Бесплатный доступ
Азоспирилл, способный стимулировать рост растений, относится к бактериям, способствующим росту растений. Лектин, обнаруженный на его поверхности, может связывать определенные углеводы и обеспечивает адгезию бактерий к поверхности корня. Мы исследовали влияние лектинов двух штаммов - A. brasilense Sp7 (эпифитный штамм) и A. brasilense Sp245 (эндофитный штамм) - на активность антиоксидантных ферментов в корнях 4-дневных проростков пшеницы, подвергшихся воздействию тяжелых металлов. (CoSO4, ZnSO4, Pb (CH3COO) 2 и CuSO4). При всех стрессах оба лектина увеличивали активность пероксидазы и супероксиддисмутазы и снижали активность каталазы, но периоды воздействия и соответствующие концентрации были различными. Эти различия могли быть вызваны различной структурой и углеводной специфичностью лектинов, что привело к различиям во взаимодействии с поверхностью растительных клеток, и такие различия имеют решающее значение для «включения» последующих стадий. Лектины Azospirillum участвуют в адаптационных изменениях корней проростков пшеницы. Это вовлечение способствует нормальному обмену веществ и обеспечивает регуляцию взаимодействия растения с азоспирилом в широком диапазоне почвенных и климатических факторов.
Короткий адрес: https://sciup.org/143166904
IDR: 143166904
Study of the possible involvement of Azospirillum lectins in the mechanisms of adaptive response of plants exposed to heavy metal salts
Azospirillum, which has the potential to stimulate plant growth, belongs to plant-growth-promoting bacteria. The lectin found on its surface can bind specific carbohydrates and ensures adhesion of the bacteria to the root surface. We examined the effect of the lectins from two strains - A. brasilense Sp7 (epiphytic strain) and A. brasilense Sp245 (endophytic strain) - on the activities of antioxidant enzymes in roots of 4-day-old seedlings of wheat exposed to heavy metals (CoSO4, ZnSO4, Pb(CH3COO)2 and CuSO4). Under all stresses, both lectins increased peroxidase and superoxide dismutase activities and decreased catalase activity, but the periods of effect and the concentrations involved were different. These differences may have been caused by the different structures and carbohydrate specificities of the lectins, which resulted in differences in the interaction with the plant cell surface such differences are of deciding importance for the “switch on” of the subsequent stages. Azospirillum lectins are involved in adaptational changes in wheat seedling roots. This involvement promotes the normal course of metabolism and ensures regulation of the plant- Azospirillum interaction in a wide range of soil and climatic factors.
Текст научной статьи Изучение возможного участия лектинов Azospirillum в механизмах адаптивного ответа растений, подвергшихся воздействию солей тяжелых металлов
Ассоциативные азотфиксирующие бактерии рода Azospirillum относятся к микроорганизмам, стимулирующим развитие и рост растений. Это достигается за счет ряда эффектов – способности к азотфиксации, продукции фитогормонов, солюбилизации фосфатов, улучшению водного и минерального статуса, продукции ряда молекул, приводящих к увеличению мембранной активности и пролиферации тканей корней, способности нивелировать влияние стрессоров на растение и осуществлять контроль многочисленных фитопатогенов (Bashan et al ., 2014). Данные бактерии способны индуцировать у растений защитные реакции, которые направлены на повышение устойчивости (Bhattachar a, Jha, 2012). До сих пор, несмотря на активные исследования в этой области, вопрос о приоритетности какого-либо из перечисленных факторов, оказывающих благоприятное влияние при инокуляции азотфиксирующими бактериями на развитие растения, остается открытым.
Штаммы азоспирилл - A. brasilense Sp7 и Sp245 интересны тем, что они относятся к наиболее изученному виду этих бактерий и отличаются стратегией поведения в процессе формирования симбиозов (Bashan et al ., 2014). В частности, штамм A. brasilense Sp7 был обнаружен только на поверхности корня, в тоже время бактерии штамма A. brasilense Sp245 способны колонизировать растительные ткани и относятся к эндофитам. Как показали исследования, проведенные с использованием метода сканирующей конфокальной микроскопии, бактерии штамма Sp245 способны к тесному контакту с растением-хозяином: они проникают в проводящую систему корня пшеницы и заполняют корневые волоски (Xie, Yokota, 2005). Эндофитные бактерии представляют особый интерес, поскольку они в меньшей степени зависят от внешних факторов среды по сравнению с другими микроорганизмами и проявляют комплекс хозяйственно полезных свойств. Эндофитные бактерии способны обеспечивать длительную защиту макроорганизма от стрессовых факторов окружающей среды (Bashan et al ., 2014).
Согласно представлениям, описанным в литературе, образование азотфиксирующих систем, подобно любым другим биологическим межклеточным взаимодействиям основано на функционировании углеводсвязывающих белков – лектинов. Долгое время главную роль узнающих молекул в системе углевод-белкового взаимодействия при формировании азотфиксирующих ассоциаций и симбиозов отводилась лектинам растений (Антонюк, Евсеева, 2006). Однако новые исследования, приведшие к появлению новых знаний относительно лектинов азотфиксирующих бактерий заставило внести заметные изменения в представления по лектин-углеводным взаимодействиям, реализуемым при становлении ассоциаций растений с азотфиксирующими бактериями с учетом роли бактериальных лектинов (Castellanos et al. , 1998; Никитина и др., 2005).
Было показано, что самые первые стадии прикрепления бактерий к корням в процессе образования ассоциации происходят за счет лиганд-рецепторного взаимодействия. Достоверно установлено, что со стороны азоспирилл в этом процессе принимают участие углеводсвязывающие гликопротеины, находящиеся на поверхности клетки (Антонюк, Евсеева, 2006). С поверхности двух штаммов бактерий – A. brasilense Sp7 и Sp245 были выделены лектины с различными молекулярными массами и углеводной специфичностью. Лектин эпифитного штамма A. brasilense обладал молекулярной массой 36 кДа и был специфичен к L-фукозе и D-галактозе (Никитина и др., 2005). Лектин A. brasilense Sp245 характеризовался молекулярной массой 67 кДа и проявлял углеводсвязывающую активность к своему полисахариду – кислому D-рамнану (Шелудько и др., 2009).
В более ранних исследованиях было показано, что лектины азоспирилл являются многофункциональными молекулами. Они принимают участие не только в адгезии бактерий к корням растений, но также модифицируют метаболизм растительной клетки – усиливают прорастание семян, обладают ферментмодифицирующей и митогенной активнос- тями (Alen’kina et al., 2017, 2018; Никитина и др., 2005), изменяют количество стрессовых веществ в растительной клетке (Alen’kina et al., 2014).
С каждым годом увеличивающееся поступление в окружающую среду тяжелых металлов приводит к загрязнению почвы. Почва же, в свою очередь, является основным источником поступления солей тяжелых металлов в растения. Накопление тяжелых металлов в растениях приводит к снижению количества и качества урожая сельскохозяйственных растений. В связи с этим становится понятной необходимость изучения механизмов их поступления в растения. Так как токсическое действие тяжелых металлов сильно сказывается на росте культурных растений, которые настоящее время часто культивируются в окрестностях крупных городов, эта проблема весьма важна. В связи с тем, что почва практически не способна произвести самоочищение и скорость этого процесса довольно низка, это создает большие проблемы для растениеводческой отрасли (Cho, Seo, 2005).
Как было отмечено выше, воздействие тяжелыми металлами является одним из разновидностей абиотических стрессов, оказывающих влияние на растения. В результате данного стресса в растительном организме происходит излишнее накопление активных форм кислорода, которые в свою очередь способны вызывать повреждение нуклеиновых кислот, перекисное окисление липидов, окисление белков, инактивацию ферментов. У высших растений имеются сложные системы антиоксидантной защиты растительных клеток, важнейшей частью которых являются супероксиддисмутазы, каталазы, некоторые пероксидазы (Cho, Seo, 2005). Супероксиддисмутаза - фермент, относящийся к классу оксидоредуктаз, осуществляющий разложение супероксид-радикала с образованием перекиси водорода. Этот фермент обнаружили у всех живых организмов, но все три изоформы этого фермента имеются лишь в растениях. Каталаза и пероксидаза являются одними из наиболее важных ферментов антиоксидантной защиты, способных детоксифицировать перекись водорода. Они присутствуют в различных органоидах растительных клеток (Fo er, Noctor,
2015). Многими исследованиями показано, что изменение активности этих ферментов в условиях воздействия ионов тяжелых металлов связано с развитием устойчивости растений к данному стрессу. В настоящее время, одним из перспективных направлений повышения устойчивости растений к воздействию различных стрессовых факторов является внедрение экологически чистых технологий и методов, основанных на применении как эпифитных, так и эндофитных штаммов азоспирилл, обладающих ростстимулирующей активностями (Xie, Yokota, 2005). Многими исследователями показано, что азоспириллы могут проявлять антистрессовый эффект в условиях воздействии на растения различных абиотических стрессов (Bhattachar a, Jha, 2012). Однако сведения о механизмах данного эффекта, в том числе и при воздействии на растения тяжелых металлов, в литературе отсутствуют.
Цель нашей работы состояла в выявлении способности лектинов двух штаммов азоспирилл – A. brasilense Sp7 и A. brasilense Sp245 оказывать регулирующее влияние на активность пероксидазы, каталазы и супероксиддисмутазы в корнях проростков пшеницы в условиях загрязнения солями тяжелых металлов – CoSO4, ZnSO4, Pb(CH3COO)2 и CuSO4.
MATERIALS AND METHODS
Объектом исследования служили два штамма бактерий рода Azospirillum – эпифитный штамм A. brasilense Sp7 и эндофитный штамм A. brasilense Sp245 из коллекции микроорганизмов ИБФРМ РАН .
Выделение изучаемых белков с клеточной поверхности бактерий проводили ранее описанным методом (Никитина и др., 2005). Лектиновую активность определяли с помощью реакции гемагглютинации с использованием 2%-ной суспензии трипсинизированных кроличьих эритроцитов.
Семена пшеницы Triticum aestivum L. сорта «Саратовская 29» (ГНУ НИИ Сельского хозяйства Юго-Востока РСХА, Саратов, Россия) были поверхностно стерилизованы в 70% (v/v) этаноле 1 мин, затем многократно отмыты стерильной водой. Для получения этиолированных корней проростков семена были выращены в чашках Петри на стерильной дистиллированной воде при 25ºC. В экспериментах были использованы четырехдневные проростки.
Для постановки экспериментов корни в течение двух часов подвергали совместному воздействию изучаемых лектинов в концентрациях от 5 до 40 мкг/ мл и солей тяжелых металлов – CoSO4, ZnSO4, Pb(CH COO) , CuSO в концентрации 10-3 мМ. 32 4
Корни подвергали гомогенизированию в 0.15 М фосфатно–солевом буфере с рН 7.8. Гомогенат центрифугировали при 7000g 10 мин, супернатант использовали для определения ферментативной активности.
Концентрацию белка определяли по методу Бредфорд (Bradford, 1976). Активность пероксидазы (ЕС 1.11.1.7) определяли с использованием микрометода, основанного на окислении о -фенилендиамина (Хайруллин и др., 2001). Для этого к 50 мкл надосадочной жидкости, предварительно разбавленной фосфатно–солевым буфером (рН 5.6) в 20 раз добавляли 25 мкл раствора ОФД в концентрации 0.5 мг/мл. Затем через 2 мин вносили 25 мкл 0.43 мМ Н2О2. Реакцию останавливали добавлением 50 мкл 4N серной кислоты. Оптическую плотность образцов измеряли при длине волны 492 нм. Активность ферментов выражали в единицах поглощения на 1 г сырой массы корней. Для сравнения вариантов активность выражали в относительных единицах.
Определение активности каталазы (EC 1.11.1.6) проводили по методу Aebi (Aebi, 1984). Снижение количества перекиси водорода измеряли при длине волны 240 нм. За единицу активность принимали мкM H2O2, разлагаемой в минуту на г сырой массы корней (коэффициент экстинкции 39.4 мМ-1 см-1). Для сравнительного анализа опытных вариантов активность выражали в относительных единицах.
Активность супероксиддисмутазы (EC 1.15.1.11) в системе НАДН и феназинметасульфата определяли по ингибированию скорости восстановления нитросинего тетразолия (Alscher et al., 2002). Спектрофотометрическое определение концентрации формазана проводили при 560 нм и использовали для расчета активности фермента. Результаты выражали в относительных единицах.
Для статистической обработки результатов использовали критерий Стьюдента. Результаты приведены как средние арифметические значения по пяти опытам, проведенным в 5-кратной биологической повторности с их стандартными ошибками. Обсуждаются величины, достоверные при P<0.05.
RESULTS AND DISCUSSION
В результате проведенных нами опытов было установлено, что комбинированное воздействие лектинов A. brasilense Sp7 и A. brasilense Sp245 c CoSO , ZnSO , Pb(CH COO) приводило к 4 4 32
повышению активности пероксидазы в корнях проростков пшеницы. Активность фермента в случае с лектином A. brasilense Sp245 максимально возрастала после 15-минутной экспозиции с корнями в присутствии CoSO4, ZnSO4 и 30-минутной экспозиции в присутствии Pb(CH3COO)2. В варианте солью цинка наибольшую эффективность лектин проявлял в минимальной концентрации (5 мкг/мл), с солью кобальта – 10 мкг/мл, с солью свинца – 10 мкг/ мл. В случае с лектином эпифитного штамма эффект был отмечен после часа инкубации с корнями и концентрации лектина 20 мкг/мл для всех вариантов. Воздействие обоих лектинов в присутствии СuSO4 не оказывало эффекта на активность пероксидазы (рис. 1).
Рассмотрение антиоксидантной системы невозможно без оценки функционирования фермента детоксикации образовавшейся Н2О2 – каталазы. Активность растительной каталазы часто рассматривается как показатель загрязнения среды, в которой развивается данное растение. При изучении воздействия изучаемых белков на корни в присутствии всех изучаемых солей тяжелых металлов происходило уменьшение активности каталазы, причем это понижение происходило на фоне повышения активности этого фермента при воздействии изучаемых солей тяжелых металлов на корни проростков пшеницы.
В случае с лектином A. brasilense Sp7 максимальное уменьшение происходило после 30
мин инкубации с корнями в присутствии всех металлов, затем происходило снижение эффекта и к часу инкубации лектинов с корнями она достигала уровня воздействия одними лектинами. Максимальный эффект был отмечен в варианте с Pb(CH3COO)2 при концентрации лектина – 10 мкг/мл.
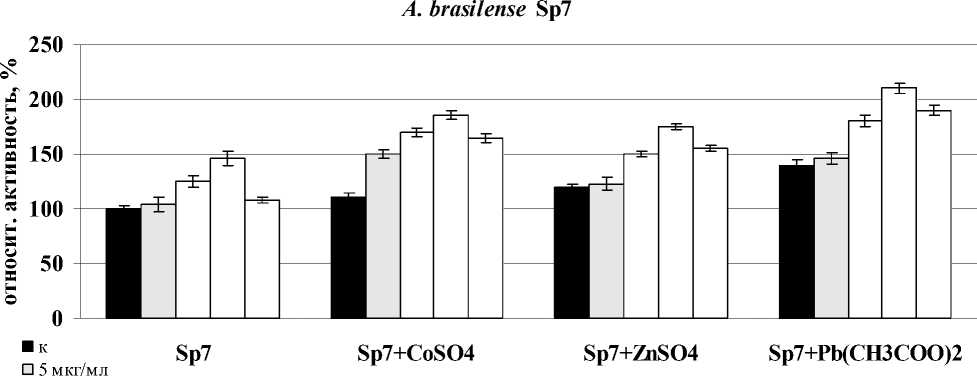
□ 10 мкг/мл
□ 20 мкг/мл
□ 40 мкг/мл
время инкубации, мин
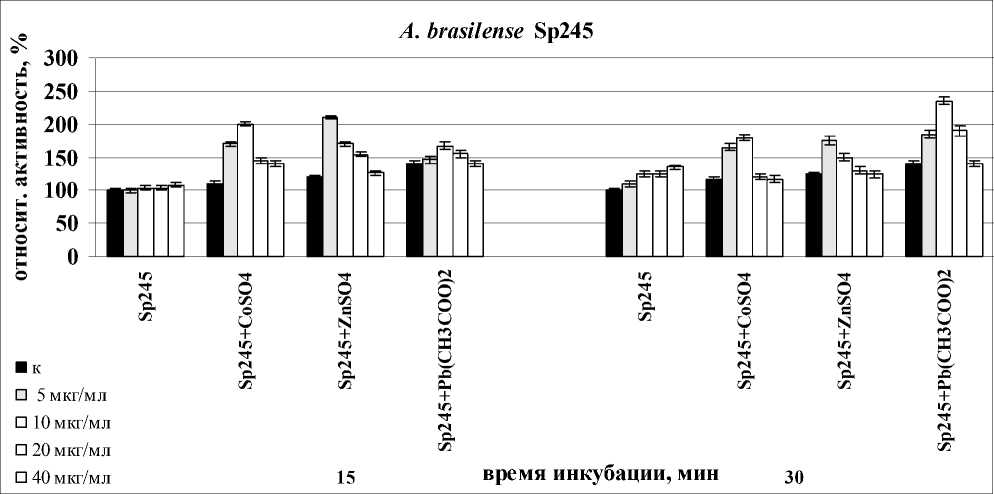
Figure 1. Активность пероксидазы в корнях проростков пшеницы в присутствии лектинов. Результаты представлены как средние арифметические значения со стандартной ошибкой ( n =5). Все различия достоверны ( p <0.05).
A. brasilense Sp7
□ 10 мкг/мл
□ 20 мкг/мл
время инкубации, мин
□ 40 мкг/мл
A. brasilense Sp245
□ 5 мкг/мл
□ 10 мкг/мл
□ 20 мкг/мл
□ 40 мкг/мл
время инкубации, мин
Figure 2. Активность каталазы в корнях проростков пшеницы в присутствии лектинов. Результаты представлены как средние арифметические значения со стандартной ошибкой ( n =5). Все различия достоверны ( p <0.05).
Для лектина A. brasilense Sp245 уменьшение активности происходило после 15 мин совместного воздействия с солями тяжелых металлов. Максимальный эффект был отмечен при концентрации лектина – 5 мкг/мл для варианта с Pb(CH3COO)2 (рис. 2).
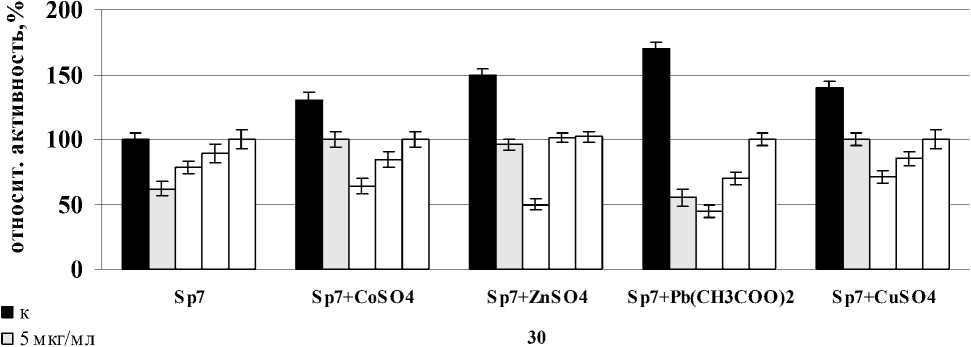
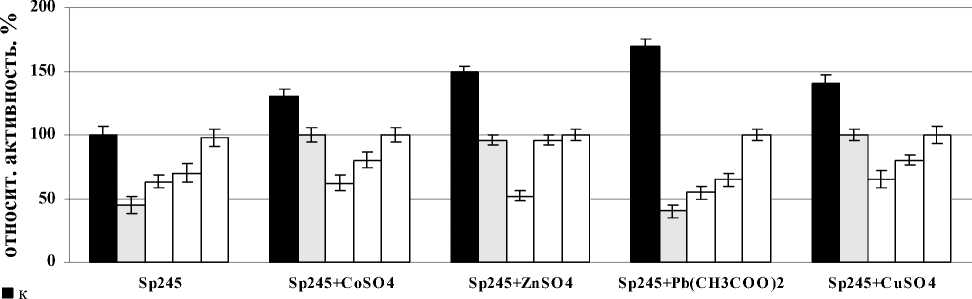
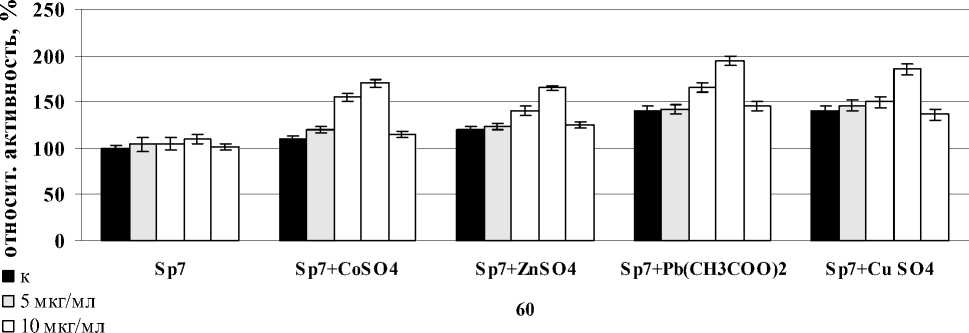
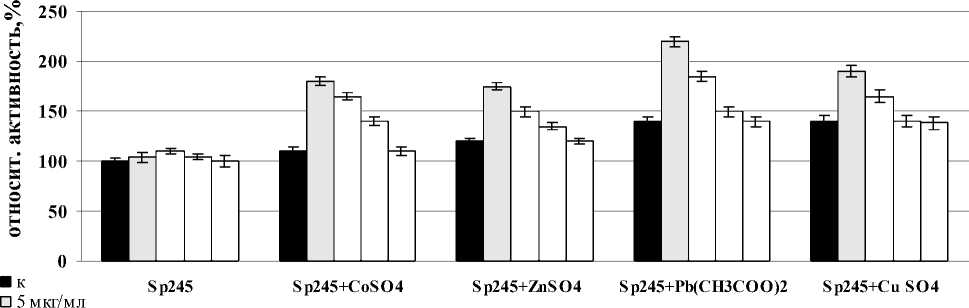
Было показано, что лектины A. brasilense Sp7 и Sp245 вызывали индукцию активности СОД в корнях проростков пшеницы после совместного воздействия со всеми солями тяжелых металлов после часа инкубации с корнями. В случае лектина A. brasilense Sp7 наибольший эффект в отношении фермента был отмечен для всех вариантов солей при концентрации лектина – 20 мкг/мл, но максимальное значение было отмечено в варианте с Pb(CH3COO)2. Сравнивая между собой изменения активности СОД в корнях проростков при совместном воздействии лектина A. brasilense Sp245 и солей – CoSO4, ZnSO4, Pb(CH3COO)2, CuSO4, было показано, что максимальное повышение активности фермента во всех случаях наблюдалось при концентрации лектина – 5 мкг/мл. Так же, как и в случае с лектином A. brasilense Sp7, максимальное значение было отмечено в варианте с Pb(CH3COO)2 (рис. 3). Можно предположить, что усиленная ответная реакция при воздействии Pb(CH3COO)2 объясняется тем, что свинец не относится к необходимым питательным элементам растений.
A. brasilense Sp7
□ 20 мкг/мл время инкубации, мин
□ 40 мкг/мл
A. brasilense Sp245
время инкубации, мин
□ 10 мкг/мл
□ 20 мкг/мл
□ 40 мкг/мл
Figure 3. Активность СОД в корнях проростков пшеницы в присутствии лектинов. Результаты представлены как средние арифметические значения со стандартной ошибкой ( n =5). Все различия достоверны ( p <0.05).
Для всех изучаемых ферментов лектин A. brasilense Sp7 проявлял максимальный эффект в отношении фермента при более высоких концентрациях, чем лектин A. brasilense Sp245 и уровень эффекта для лектина эндофитного штамма был значительно выше по сравнению с лектином другого штамма. Вероятной причиной отличающейся функциональной активности лектинов могут быть различия в углеводной специфичности, структуре белков, и как следствие, неодинаковое взаимодействие с поверхностью растительной клетки, что является определяющим фактором для включения последующих этапов.
Представленные данные подтверждают результаты других авторов, которые отмечают способность азоспирилл изменять активность антиоксидантных ферментов в растениях при различных абиотических стрессах (Bhattachar a, Jha, 2012). Одной из возможных причин повышения активности пероксидазы, СОД и угнетения каталазной активности может быть влияние салициловой кислоты, индукцию синтеза которой вызывают лектины азоспирилл (Alen’kina et al., 2014).
CONCLUSION
Таким образом, лектины азоспирилл можно отнести к регуляторам роста растений, обладающих не только росторегулирующим, но и антистрессовым и иммуностимулирующим действием, что может явиться основой разработки новых технологий повышения продуктивности растений.
Список литературы Изучение возможного участия лектинов Azospirillum в механизмах адаптивного ответа растений, подвергшихся воздействию солей тяжелых металлов
- Антонюк Л.П., Евсеева Н.В. (2006). Лектин пшеницы как фактор растительно-микробной коммуникации и белок стрессового ответа. Микробиология, 75, 544-549
- Никитина В.Е., Пономарева Е.Г., Аленькина С.А. (2005) Лектины клеточной поверхности азоспирилл и их роль в ассоциативных взаимоотношениях с растениями//Молекулярные основы взаимоотношений ассоциативных микроорганизмов с растениями/Под ред. В.В. Игнатова. М.: Наука, 70-97
- Шелудько, А.В., Пономарева Е.Г., Варшаломидзе О.Э., Ветчинкина Е.И., Кацы Е.И., Никитина В.Е. (2009) Гемагглютинирующая активность и подвижность бактерий Azospirillum brasilense в присутствии разных источников азота. Микробиология, 78, 749-756
- Хайруллин Р.M, Яруллина Л.Г., Трошина Н.Б., Ахметова И.Э. (2001) Активация хитоолигосахаридами окисления орто-фенилендиамина проростками пшеницы в присутствии щавелевой кислоты. Биохимия, 66, 354-358
- Aebi H. (1984) Catalase in Vitro. In Packer, L., (Ed.), Methods in Enzymology/H. Aebi -Academic Press, San Diego. 105, 121-126
- Alen’kina S.A., Bogatyrev V.A., Matora L.Yu., Sokolova, M.K., Chernysheva M.P., Trutneva K.A., Nikitina V.E. (2014) Signal effects of the lectin from the associative nitrogen-fixing bacterium Azospirillum brasilense Sp7 in bacterial-plant root interactions. Plant and Soil, 381, 337-349
- Alen’kina S.А., Nikitina V.Е. (2017) Change in the ratio of the activities of different types of proteases and their inhibitors in plant roots exposed to Azospirillum lectins. J Plant Regulation, 36, 522-527
- Alen’kina S.А., Romanov N.I., Nikitina V.Е. (2018) Regulation by Azospirillum lectins of the activity of antioxidant enzymes in wheat seedling roots under short-term stresses. Brazilian Journal of Botany, 41, 579-587
- Alscher R.G., Erturk N., Heath L.S. (2002) Role of superoxide dismutases (SODs) in controlling oxidative stress plants. J. Exp. Bot., 53, 1331-1341
- Bashan Y., de-Bashan L.E., Prabhu S.R., Hernandez J.-P. (2014) Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspectives (1998-2013). Plant Soil, 378, 1-33
- Bhattacharyya P.N., Jha D.K. (2012) Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J. Microbiol. Biotechnol., 28, 1327-1350
- Bradford M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72, 248-254
- Castellanos T., Ascencio F., Bashan Y. (1998) Cell-surface lectins of Azospirillum spp. Curr. Microbiol., 36, 241-244
- Cho U.H., Seo N.H. (2005) Oxidative Stress in Arabidopsis thaliana Exposed to Cadmium Is due to Hydrogen Peroxide Accumulation. Plant Sci., 168, 113-120
- Foyer C.H., Noctor G. (2015) Defining robust redox signalling within the context of the plant cell. Plant Cell and Environment., 38, 239-239
- Xie C.H., Yokota A. (2005) Azospirillum oryzae sp. nov., a nitrogen-fixing bacterium isolated from the roots of the rice plant Oryza sativa. Int. J. Syst. Evol. Microbiol., 55, 1435-1438
