Явление квантированности роста в метамерной организации побега некоторых представителей семейства Аlismataceae vent

Автор: Лапиров Александр Григорьевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Материалы лекций и устных докладов
Статья в выпуске: 9, 2008 года.
Бесплатный доступ
Исследовано явление квантированности роста у частухи подорожниковой (Alisma plantago-aquatica L.) и частухи злаковой (Alisma gramineum Lej.). Показано, что у этих видов соцветие закладывается только после определенного числа не отдельных фитомеров, а целых «квантов». Данный показатель у обоих изучаемых растений колеблется в пределах 4 - 6 метамеров.
Короткий адрес: https://sciup.org/146116169
IDR: 146116169 | УДК: 582.536-14/.5
Текст научной статьи Явление квантированности роста в метамерной организации побега некоторых представителей семейства Аlismataceae vent
Институт биологии внутренних вод им. И.Д. Папанина РАН, Борок
Исследовано явление квантированности роста у частухи подорожниковой (Alisma plantago-aquatica L. ) и частухи злаковой (Alisma gramineum Lej .). Показано, что у этих видов соцветие закладывается только после определенного числа не отдельных фитомеров, а целых «квантов». Данный показатель у обоих изучаемых растений колеблется в пределах 4 – 6 метамеров.
Явление «квантированности» роста, выражающееся в виде «…тенденциозного варьирования числовых и размерных признаков, можно рассматривать в качестве … общей закономерности … роста и органогенеза у растений…» [22]. В последнее время это явление привлекает все большее внимание ботаников [14].
Исследуя закономерности роста растений, Д.А. Сабинин [16; 17] пришел к выводу о строгой ритмичности данного процесса. При этом он, формулируя понятие о ритмах роста, ссылался на высказывание W. Zimmerman (1935, цит. по: [16]) о квантированности формообразовательных процессов.
Термин «кванты роста» впервые был введен в обиход А.В. Гурским в 1948 г. (цит. по [22]), который применил его к описанию особенностей роста древесных побегов1. Автор термина писал: «…удлиненные побеги древесных растений, по числу образующихся на них метамеров, во многих случаях представляют собой комбинацию одного, двух и более укороченных побегов. Таким образом, намечается весьма важный принцип дозировки, « квантированности» образования метамеров на побегах … принцип кратных отношений роста по числу метамеров» [3, с. 19]. Позднее А.В. Гурский и соавторы [4] подчеркивали, что это явление характерно для многих видов древесных растений. Кроме того, В.М. Шмидт [22, с. 100] отмечает, что « Гурский сообщил нам, что установленный им принцип [«кратных отношений»] в более общей форме справедлив и для травянистых растений». Более того, Т.И. Серебряковой [19] при изучении семейства злаковых было описано не только явление квантированности числа метамеров побегов, но и обнаружена совершенно отчетливая связь между числом фитомеров в зрелой почке и квантом фитомеров, составляющих побег в целом. Подобное явление было отмечено и при изучении ростовых процессов у горных тундровых растений (травянистых розеткообразующих, подушковидных и кустарников) [21].
Таким образом, ритмичность, или «квантированность» в структуре побегов была изучена на древесных растениях, кустарниках, кустарничках, травянистых многолетних растениях и злаках [19; 21; 22]. В работах данных авторов, кроме того, содержатся литературные обзоры по этому вопросу. По низкотравным гелофитам подобная информация в отечественных источниках нами не обнаружена. В немногочисленной иностранной литературе квантированности роста побега у этих растений серьезного внимания не уделяется, а лишь констатируется факт наличия между соцветиями определенного числа листовых зачатков, т.е. периодичности в развитии соцветий [24 – 28].
Именно поэтому целью нашей работы явилось изучение явления «квантированности» роста в структуре побегов у частухи подорожниковой ( Alisma plantago-aquatica L.) и частухи злаковой ( Alisma gramineum Lej.).
При этом рассматривать данный вопрос мы будем, учитывая мысль O. Schüepp (1938, цит. по: [21, с. 13], что «… каждая форма должна быть понята и объяснена как результат роста» и что «…становление морфы происходит на основе ритмов роста» [21, с. 14]. Кроме того, за «квант», следуя Т.И.Серебряковой [19, с. 186], принимаем «порцию из нескольких фитомеров, количественно нередко совпадающую с емкостью закрытой или открытой почки».
Alisma plantago-aquatica обладает широкой экологической амплитудой и встречается по берегам озер, рек, на мелководьях водохранилищ, в прудах, канавах, каналах, кюветах, в местах с поверхностным и грунтовым подтоплением [5; 11; 12; 20].
Alisma gramineum — вид, интенсивно распространяющийся в бассейне Волги [12]. Встречается на мелководьях водохранилищ, озер и рек, песчаных отмелях, у воды и в воде на глубине до одного метра, в стоячих водах, и ее встречаемость и размеры популяции изменяются от года к году. [11; 13; 15]. Данный вид интенсивно расселился по всему Рыбинскому водохранилищу, образуя чистые или почти чистые заросли на песчаных отмелях, заходя в воду на глубину до 30 – 80 см, формируя две хорошо различающиеся формы – надземную и полностью погруженную.
Для проведения исследований A. plantago-aquatica и A. gramineum были собраны в природных местообитаниях, характерных для данных видов. Частуха подорожниковая – на мелководье на левом берегу Рыбинского водохранилища напротив д. Глебово и увлажненном понижении вблизи копани в д. Григорово, частуха злаковая – в порту близ поселка Борок и на левом берегу канала у ихтиологического корпуса (мелководье Рыбинского водохранилища), а также на реке Сутка (близ Спицинского ручья), впадающей в данное водохранилище.
Для понимания сути вопроса, рассматриваемого нами в данной статье, необходимо прежде всего отметить, что до момента образования соцветий главный побег у этих растений нарастает моноподиально. У частухи подорожниковой этот процесс охватывает два вегетационных сезона (т.е. главный побег является дициклическим), а у частухи злаковой – один. С момента заложения генеративных органов нарастание главного побега двух видов частухи становится симподиальным. Некоторые авторы [23; 25; 28] считают, что это результат раздвоения апикальной меристемы, которое характерно для многих, если не всех таксонов Alismatidae [28]. Однако наши исследования [8; 9], а также рисунки и описание этого процесса у предыдущих авторов, материалы Н.П. Савиных, Е.В. Лелековой [10; 18] показывают, что в данном случае мы имеем дело с акросимподиальным нарастанием (в понимании Л.Е. Гатцук [1]), которое происходит за счет одной из верхних пазушных почек. Как отмечает Л.Е. Гатцук [1, стр. 87] « если оно связано с образованием цветка или соцветия (в самом узком смысле – без ассимилирующих листьев) на верхушке моноподиального побега, это самая верхняя почка вегетативной сферы».
Данный процесс, ранее обнаруженный нами у частухи подорожниковой и частухи злаковой [8; 9], детально описан Е.В. Лелековой [10] у A. plantago-aquatica и заключается в том, что одновременно с началом цветения главного побега развивается пазушная почка самого верхнего метамера вегетативной сферы, образуя побег замещения n+1 порядка. Утолщаясь, его базальные метамеры сдвигают верхушечное соцветие первого порядка в сторону (создается впечатление как бы пазушного его расположения). Подобный «сценарий» развития продолжается и далее, формируя последовательные терминальные соцветия и побеги замещения n+2 и последующих порядков из соответствующих пазушных почек самых верхних метамеров (рисунок). Такой же путь формирования побеговой системы выявлен нами и у частухи злаковой.
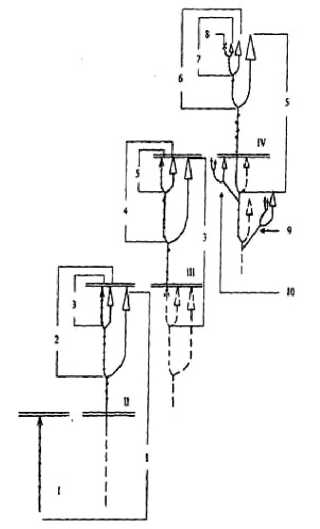
Формирование побеговой системы особи Alisma plantago-aquatica семенного происхождения до естетственного отмирания верхушечной почки побега последнего порядка (листья не показаны, узлы расставлены) (по: [10])
I – V – годы жизни особи; 1 – 7 – побеги последовательных порядков симподиальной оси; 8 – верхушечная почка побега последнего порядка; 9 – 10 – боковые симподии; = -окончание вегетационного периода. Остальные пояснения – в тексте.
Нами показано, что при этом соцветие закладывается только после определенного числа не отдельных фитомеров, а целых «квантов». Данный показатель у обоих изучаемых растений колеблется в пределах 4 – 6 метамеров. В этом случае мы говорим о вегетативных метамерах, формирующих последовательные побеги замещения в результате акросимподиального нарастания главного побега. Кроме того, в составе базального метамера каждого из побегов замещения содержится профилл – редуцированный двухкилевой пленчатый лист. В типичном случае (при закладке генеративных органов) наличие профилла является не только показателем перехода к симподиальной форме роста. Он также разграничивает «кванты роста», которые, по сути, являются отдельными элементарными побегами (термин И.А. Грудзинской [2]). A.J. Eames (1961, цит. по: [28]) указывал, что профиллы покрытосеменных это– «листья редуцированной формы» («leaves of reduced form») и «слабо развитые» («weakly developed»). W.A. Charlton (1973, цит. по [28]) для Alisma plantago-aquatica L. они трактуются как профиллы побегов возобновления на вертикальной оси (prophylls of renewal shoots on upright axes). Однако, как справедливо считают Н.П. Савиных и Е.В. Лелекова [18], с. 101], у частухи подорожниковой «…Побеги замещения различны по своему происхождению: если первые два закладываются в год, предшествующий цветению, и, по сути, могут быть охарактеризованы как побеги возобновления, то следующие – типичные силлептические побеги ветвления, развивающиеся из почек обогащения….». В отличие от этого, у частухи злаковой все побеги замещения силлептические. У частухи подорожниковой в течение вегетационного периода формируется до четырёх побегов замещения, у частухи злаковой – до трёх. Такое беспрерывное образование за вегетационный период нескольких, следующих друг за другом побегов замещения, формирующих скелетную ось растения, представляет собой итеративное ветвление (термин Troll, 1964; цит. по: [6]). Учитывая то, что пазушная почка у Alisma plantago-aquatica L., которая закладывается в конце вегетационного периода, в виде побега замещения следующего порядка содержит от 12 – 18 зачатков, справедливо утверждение, что «…итеративное ветвление характерно уже для почек этого растения…» [18], с. 101].
Необходимо также отметить, что, как и у злаков [19], если соцветие не заложилось, то «кванты» фитомеров соцветия становятся вегетативными. Так, у двух видов частухи нами были обнаружены растения, содержащие (8) 10 – 12 метамеров (2 кванта по (4) 5 – 6 метамеров) до первого соцветия, тогда как до второго и последующих соцветий сохранялось от 4 до 6 метамеров. По данным Н.П. Савиных и Е.В. Лелековой [18], шесть вегетативных метамеров обнаружено между последовательными соцветиями у Alisma plantago-aquatica L в Кировской области.
Кроме того, заметим, что результаты наших исследований не подтверждают данные G.J. Wydler (1863, цит. по: [25, с. 2349], что «…все более и более меньшее количество листьев образуется между соцветиями в течение вегетационного периода… (… that increasingly fewer leaves were produced between inflorescences over the growing season in Alisma plantago-aquatica. ) » . Это подтверждает мысль S.M. Liev [25], с. 2349] о том, что «… определенная вегетативная стадия может иметь иные различающиеся характеристики между европейским Alisma plantago-aquatica и североамериканским Alisma triviale, иногда рассматриваемыми как один и тот же вид» («… a definite vegetative phase may be another distinguishing characteristic between the European A. plantago-aquatica and North American A. triviale, sometimes considered the same species») (Hendricks, 1957; цит. по: [25], с. 2349].
По-видимому, решающую роль в формировании побегов замещения (их количестве и степени развития) у двух видов частухи играют условия вегетационного периода, особенно температура и водность года. Кроме того, Н.В. Шилова [21] считает, что большое приспособительное значение квантированность (порционность) и ритмичность ростовых процессов играют при сокращении вегетационного периода.
Говоря о низкотравных гелофитах, интересно отметить, что, по данным зарубежных исследователей, у Alisma triviale (вида, близкого к Alisma plantago-aquatica L.) между соцветиями формируется 6 или, иногда, 5 листьев (включая мембранный профилл – [25]); 5 листьев обнаружено у Alisma subcordatum [24]. У Butomus umbellatus между соцветиями обнаружено нечетное количество листьев (9, 7 – Irmisch, 1850; Buchenau, 1903 – все (цит. по: [27]) или 5 [27]. У Sagittaria latifolia, по данным S.M. Liev [25; 26], формируется 3, а по данным R.B. Kaul [24], – 5 листьев между соцветиями, так же как и у Sagittaria cuneata и Sagittaria brevirostra [24].
М.М. Магомедмирзаев ([14]: 10) справедливо заметил, что «…число и размер элементов морфологической организации растений выступают как следствие фундаментального свойства высшего растительного организма – метамерности развития…», под которой понимают « последовательное повторное онтогенетическое заложение однозначных органов или оформленных отчленений» [7], с. 18]. При этом «… числа метамеров 2, 3, 5 характеризуют « архетипы» растений, повторяясь с удивительным постоянством в структуре самых разных представителей растительного мира…» [14], с. 118].
The phenomenon of quantum growth in Alisma plantago-aquatica L. and Alisma gramineum Lej. is investigated. It is shown, that in these species the inflorescence is laid only after a certain number of not just separate phytomeres but whole "quanta". This parameter in both the studied plants ranges from 4 – 6 metameres.
