Эдафические стрессовые факторы Северо-Востока европейской части России и проблемы селекции растений

Автор: Лисицын Е.М., Шихова Л.Н., Овсянкина А.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Проблемы плодородия почвы и адаптивности растений
Статья в выпуске: 3 т.39, 2004 года.
Бесплатный доступ
Представлены стрессовые факторы, наиболее типичные для почв Северо-Востока европейской части России. Приведены данные по влиянию на рост и развитие растений различных химических элементов. Рассматриваются особенности полевых оценок адаптированности растений к почвенным стрессам, а также доступность и сбалансированность элементов минерального питания. Описаны реакции растений на эдафические стрессы в зависимости от свойств почвы. Обсуждается проблема создания сортов, устойчивых к грибным болезням, в том числе к корневым гнилям.
Короткий адрес: https://sciup.org/142132987
IDR: 142132987 | УДК: 631.41/42:631.527:58.01/.07:631.811
Edaphic stressful factors of North-East European part of Russia and the problems of plant selection
The stressful factors were presented which are typical for soils of North-East European part of Russia. The data about the effect of different chemical elements on a growth and a development of plants are given. The authors consider the features of field estimations of plants' adaptivity to soil stress and also the availability and the balance of elements of mineral nutrition. The plants reactions on edaphic stresses in connection with soil properties were described. The problem of creation of varieties resistant to fungous diseases including root rots was discussed.
Текст научной статьи Эдафические стрессовые факторы Северо-Востока европейской части России и проблемы селекции растений
Поиск способов адаптивного ведения земледелия, стратегия которого строится на основе анализа адаптивного потенциала высших растений, невозможен без изучения влияния на последние лимитирующих экологических факторов, среди которых особое значение имеют почвенные (1). Северо-восточная зона европейской части России в целом характеризуется следующими условиями: пестрота почвенного покрова; занятость значительных территорий кислыми дерново-подзолистыми почвами; воздействие комплекса неблагоприятных факторов внешней среды; устойчивая тенденция к снижению плодородия почв. Так, по данным Центра агрохимической службы «Кировский», за период предпоследнего (1985-1992 годы) и последнего (1993-2000 годы) циклов обследования почв Кировской области площади кислых почв увеличились на 34,2 тыс. га, что наблюдается в основном в районах, где раньше проводили широкомасштабное известкование (2). При этом в 1995 году органических и минеральных удобрений было внесено соответственно в 4 и 11 раз меньше, чем в 1990 году. Если в 1990 году известкование было проведено на площади 218 тыс. га, то в 1995 и 1997 годах — соответственно на площади 15 и 2,8 тыс. га; фосфоритова-нию в 1997 году подвергли почву на площади 3,1 тыс. га, что составляет 1 % от ежегодной потребности (3, 4).
В этой ситуации вопрос о пластичности, адаптивности и устойчивости к стрессовым факторам среды сельскохозяйственных культур различных видов и сортов приобрел исключительную актуальность. Более 100 лет назад В.В. Докучаев писал, что наибольшую отдачу при возделывании сельскохозяйственных культур можно получить только тогда, когда растения будут приспособлены к местным почве, воде и климату (5). Рассмотрим наиболее типичные для почв Северо-Востока европейской части России стрессовые факторы.
П о в ы ш е н н а я к и с л о т н о с т ь п о ч в е н н о г о р а- с т в о р а. По данным научных исследований последних лет, снижение урожайности сельскохозяйственных культур на кислых почвах бывает чаще вызвано повышенной доступностью подвижных ионов Al3+, нежели высокой концентрацией ионов H+ (6-8). При кислой реакции среды (особенно на минеральных почвах) повышается растворимость алюминия, а следовательно, и содержание его подвижных форм в почвенном растворе. Попадая в растения, ионы алюминия, первоначально концентрируясь на поверхности кончиков корней и клеточных оболочках кортикальных и эпидермальных клеток, конкурируют с таковыми кальция, препятствуют активному поглощению фосфора корнями, ингибируют деление и растяжение клеток поглощающих органов (918).
Одним из эффективных агромероприятий на кислых почвах является их известкование и фосфоритование, которые необходимо повторять каждые 5-7 лет. Однако затраты на внесение фосфорных удобрений столь высоки, что не окупаются прибавками урожая, а известкование кислых почв резко сократилось. Подобная ситуация складывается не только в России. Считается, что кислыми являются около 3042
50 % всех пахотных земель в мире (19, 20). Токсичность подвижных ионов Al3+ является важнейшим лимитирующим фактором при выращивании на таких почвах растений многих видов (21-25).
Комплекс стрессовых факторов кислых почв, особенно токсичность для растений ионов Al и Mn, привлек к себе внимание ученых различных специальностей. Пионером в этом вопросе стала Бразилия, затем селекционные программы по повышению устойчивости к кислотному стрессу начали активно разворачиваться в США, Канаде, Австралии, Новой Зеландии, Колумбии, Мексике, Польше, СССР, некоторых странах Африки и др. Использование возможностей самих растений противодействовать стрессу в зоне корней позволило добиться значительных успехов в интенсификации сельскохозяйственного производства в таких странах, как Мексика, Бразилия и Китай. У растений различных видов и сортов выявлена существенная вариабельность по устойчивости к избытку алюминия в почвенной среде (26, 27). Показано, что эти различия контролируются на генетическом уровне (28-36).
Работы по отбору устойчивых к кислотности почв сортов зерновых культур начались в нашей стране еще в 40-50-е годы ХХ века (37, 38). Более трех десятилетий Климашевский с сотрудниками изучали различные аспекты кислотоустойчивости сортов и образцов кукурузы, гороха, пшеницы, ячменя, сои, вики, картофеля, в том числе воздействие алюминия на рост корней и эффективность усвоения последними элементов минерального питания (18). Эти работы свидетельствуют о приоритетной роли корневой системы в кислотоустойчивости. С 1984 года работы по оценке и выявлению сортов ячменя, устойчивых к повышенным кислотности почвы и содержанию подвижных ионов алюминия, ведутся в лабораториях НИИ сельского хозяйства Северо-Востока им. Н.В. Рудницкого (39-41). При этом были достигнуты определенные успехи в создании кислото- и алюмоустойчивых сортов основных сельскохозяйственных культур Северо-Востока европейской части России.
Физиологические и биохимические основы кислотоустойчивости до конца не определены, хотя и активно изучаются во всем мире (42). При воздействии на растение любого стрессового фактора наблюдаются одни и те же ответные реакции — тревога, адаптация и истощение (43-45). Естественно, что каждый тип стресса имеет и специфические черты ответа, но по мере усиления действия стрессового фактора (во времени или по интенсивности) на первый план все в большей степени выступают неспецифические изменения: проницаемости мембран клеток для ионов; подкисление цитоплазмы; снижение общей интенсивности синтеза полимерных макромолекул; синтез стрессовых белков; интенсификация синтеза компонентов клеточных стенок (лигнина, суберина, кутина, каллозы); повышение содержания абсцизовой и жасмоновой кислот; торможение фотосинтеза. Основные различия между устойчивыми и неустойчивыми растениями заключаются в степени проявления этих реакций, поэтому анализ последних позволяет выявлять потенциальные возможности растений противостоять разнообразным стрессам.
Особенности полевых оценок адаптиро- ванности р а с т е н и й к п о ч в е н н ы м с т р е с с а м. Наличие органической связи между климатом и уровнем плодородия почвы делает неэффективным анализ влияния на урожайность растений климатических условий отдельно от почвенных. Развитие растений находится в единстве с сезонной динамикой свойств почв и факторов почвообразования, причем эти показатели в отдельные периоды могут достигать критических значений. Поэтому факторы почвенного плодородия оказывают непосредственное влияние на продуктивность агроценозов, особенно на ответственных фазах онтогенеза растений.
Количественные и качественные показатели свойств почв меняются во времени неодинаково. Так, содержание подвижных форм элементов питания изменяется более существенно, чем их валовое количество (46). Значительная вариация показателей кислотности почвы в период вегетации растений зависит от характера водного режима, температуры, вида агроценоза, системы удобрений. На протяжении вегетационного периода возможны существенные колебания кислотно-щелочного соотношения в почве, что обусловлено влагообеспеченностью. В засушливые периоды воз-43
растает концентрация почвенного раствора, что сопровождается временным снижением кислотности почвы, которое может быть весьма существенным — на 0,5-1,0 рН (47). Гидролитическая кислотность повышается, как правило, в середине лета и зависит от буферных свойств почвы. Динамика изменений физико-химических свойств почвы служит показателем напряженности, интенсивности протекающих процессов (48).
Выбор участка почвы, на котором можно проводить оценку кислотоустойчи-вости растений, является трудной задачей, так как при этом необходимо учитывать возможность существования нескольких стрессовых факторов. Кроме того, следует иметь в виду, что алюминий может взаимодействовать с другими химическими элементами — кальцием, фосфором, магнием, железом и т.д. В отличие от марганца токсичность алюминия, содержащегося в почве, способствует снижению урожая без четко идентифицируемых симптомов нарушения развития надземной части растений, а проявление последних можно наблюдать на корнях.
Почвы, классифицированные как алюмотоксичные, значительно различаются по содержанию обменного кальция, что в свою очередь обусловливает степень токсичности алюминия и отражается на комплексе симптомов выращенных растений. Для каждой культуры реальное содержание KCl-экстрагируемого алюминия, вызывающее определенное снижение урожайности, варьирует в зависимости от типа почвы. Например, концентрация алюминия, необходимая для ингибирования роста корней хлопчатника, колебалась от 0,1 до 2,5 мг-экв/100 г почвы соответственно из Норфолка и Бладена, а уровень рН, критический для роста культуры, составлял соответственно 5,5 и менее 5,0 (49).
Для многих типов почвы трудно выделить воздействие на рост растений отдельно алюминия или фосфора. Внесение фосфора в сильно кислые почвы может заменить известкование, в других случаях большие дозы этого элемента (224 кг/га) без известкования не способствуют развитию корневой системы даже высокоустойчивых сортов пшеницы (50).
Исследование алюмоустойчивости в полевых условиях часто приводит к неожиданным результатам. Например, при оценке устойчивости разных сортов кукурузы в полевых условиях и лаборатории были получены противоположные результаты (51). Как показал анализ, это было обусловлено неодинаковой требовательностью растений разных сортов к молибдену. Аналогичные данные получены при оценке устойчивости растений различных сортов ячменя, пшеницы, сои и кукурузы к ионам меди, железа и другим микроэлементам, содержащимся в почвенном растворе (52, 53).
Отсутствие корреляции между полевыми и лабораторными оценками степени кислотоустойчивости можно объяснить, в частности, тем, что алюмоустойчивость у растений разных сортов одной и той же культуры контролируется различными генами. Если при метаболизме происходит поглощение ионов кальция или фосфора, тогда реальная доступность этих элементов в питательном растворе (в отличие от полевых условий) может маскировать сортовые различия по реакции на алюминий. Поэтому необходимо, во-первых, оценить число стрессовых факторов, имеющихся в конкретном месте проведения полевых опытов, во-вторых, — проследить за их сезонной динамикой, в-третьих, — оценить и объяснить направленность и степень сопряженности полевой и лабораторной оценок кислотоустойчивости растений.
Д о с т у п н о с т ь и с б а л а н с и р о в а н н о с т ь э л е м е н-т о в м и н е р а л ь н о г о п и т а н и я. В сильно кислой почве процессы нитрификации ингибированы и важным источником азотного питания растений являются ионы аммония. Многие растения, адаптировавшиеся к таким почвам, обладают устойчивостью не только к ионам алюминия, но и ионам аммония (NH +) в концентрациях, которые являются токсичными для неустойчивых растений. К таким растениям относятся сахарный тростник, береза, некоторые виды трав (Paspalum notatum, Lolium rigidum), дикорастущих ягодников и кофе-арабика (54-61). У растений некоторых видов это связано с отсутствием или пониженной активностью нитратредуктазы (58, 62, 63). Соотношение между формами азота в почвенном растворе определяет скорость и направление изменений рН в ризосфере в присутствии или отсутствии алюминия. У отдельных сортов пшеницы устойчивость к алюминию определяется способностью эффективно использовать нитрат-ионы при наличии ионов аммония, в результате чего повышается рН (64).
Для многих видов растений устойчивость к алюминию тесно связана с эффективностью использования фосфора (27, 65, 66). Например, алюмоустойчивость некоторых сортов гороха обусловлена высоким содержанием фосфора в корнях растений (67). Установлено, что под влиянием ионов алюминия содержание фосфора в надземной части повышается у устойчивых, но понижается у чувствительных растений бобовых культур (68). Алюминий способствует существенному повышению ре-докс-потенциала и содержания органических форм фосфора в корнях растений гороха, а также снижению содержания высокоэнергетических соединений фосфора (69). Низкое содержание фосфора в ризосфере индуцирует активность кислой фосфатазы корней и позволяет растениям экстрагировать этот элемент из органических запасов в почве (70). В то же время показано, что фосфор, связанный с алюминием, остается обменным (71, 72). По всей видимости, влияние алюминия на процессы минерального питания растений зависит от фазы вегетации последних и отражается не только на развитии корней, но и надземных органов.
Под действием различных эдафических стрессов процессы минерального питания претерпевают значительные изменения, что не может не сказаться на построении надземной части растений. Более того, в научной литературе устойчивость растений как целостного организма к стрессовым воздействиям зачастую связывают с конкретными формами азотных удобрений, используемых в сельском хозяйстве. В частности, применение аммонийной формы азота вместо нитратной может служить причиной индуцированного алюминием недостатка магния (73). Между тем при изучении разнообразных сторон деятельности растения, например процесса фотосинтеза, в условиях вегетационных и лабораторных опытов исследователи часто не учитывают таких методических аспектов, как особенности состава среды выращивания. Так, соотношение азота, фосфора и калия (мг-атом/л) могут изменяться от 54:0:46 до 68:5:27 (стандартная смесь Хогланда-Арнона) и 87:1:12 (стандартная смесь Стейнберга) (74). При этом азот может быть представлен как нитратной, так и аммонийной формой либо их смесью в различных пропорциях (59, 75-78). Сложность оптимизации соотношения трех макроэлементов минерального питания (азота, фосфора и калия) была преодолена Омесом, предложившим метод систематических вариантов с дальнейшей математической обработкой полученных данных (79).
В наших опытах растения выращивали в условиях песчаной культуры при разных соотношениях N:P:K (70:15:15; 15:70:15; 15:15:70), но одинаковом суммарном содержании, равном 22 мг-атом/кг песка, что соответствует суммарному содержанию этих элементов в стандартной питательной среде Хогланда-Арнона. В каждом варианте опыта (контроль — рН 6,0; опыт — 1 мМ Al, рН 4,3) использовали три формы азотных удобрений (NO 3 , NH 4 и NO 3 -NH 4 ) в 3-кратной повторности. Через 30 сут вегетации определяли следующие показатели развития фотосинтетического аппарата: поверхностную плотность листьев (ППЛ); площадь листьев, приходящуюся на единицу сухой массы; долю листьев в общей массе растения; содержание фотосинтетических пигментов в листьях (табл. 1).
1. Оптимальные соотношения азота, фосфора и калия при максимальной фотосинтетической активности листьев различных сортов овса под влиянием ионов алюминия (песчаная культура, 30-е сут вегетации)
|
Сорт, вариант опыта |
Форма азотного удобрения |
Содержание хлорофилла в листьях, мг/г су-хого вещества |
Поверхностная плотность листьев, г/см2 |
Площадь листьев, см2/г сухого вещества |
Доля листьев, % от общей массы растения |
|
Аргамак: контроль (рН 6,0) |
NO 3 |
51:29:19 |
34:41:25 |
44:21:35 |
41:28:31 |
|
NH 4 |
48:30:22 |
29:46:25 |
40:19:41 |
44:30:26 |
|
|
NO 3 -NH 4 |
30:25:45 |
31:35:34 |
50:25:25 |
40:20:40 |
|
|
опыт (рН 4,3; |
NO 3 |
51:27:22 |
38:32:29 |
39:37:28 |
39:29:32 |
|
1 мМ Al) |
NH 4 NO 3 -NH 4 |
35:24:41 23:40:37 |
36:38:26 28:34:38 |
43:22:35 50:28:22 |
43:28:30 47:28:35 |
|
Чиж: контроль (рН 6,0) |
NO 3 |
44:18:38 |
31:38:31 |
55:17:28 |
55:19:26 |
|
NH 4 |
29:36:35 |
38:39:23 |
42:21:37 |
50:24:26 |
|
|
NO 3 -NH 4 |
29:29:42 |
35:42:23 |
46:18:36 |
54:23:23 |
|
|
опыт (рН 4,3; |
NO 3 |
51:17:32 |
32:30:38 |
41:28:31 |
42:27:31 |
|
1 мМ Al) |
NH 4 |
44:35:21 |
26:50:24 |
41:21:38 |
40:32:28 |
|
NO 3 -NH 4 |
32:33:34 |
25:25:50 |
65:15:20 |
58:17:25 |
Относительные потребности растений в том или ином макроэлементе существенно зависели от формы азотного удобрения. Так, на ППЛ и площадь листьев, приходящуюся на единицу сухой массы растений овса сорта Аргамак, относительно чувствительного к рН почвенного раствора, не оказывала влияния кислотность почвы, если азот поставлялся в виде смешанной нитратно-аммонийной соли (50:50). В этом же варианте у растений относительно устойчивого сорта Чиж максимальные показатели активности фотосинтеза наблюдались при противоположных изменениях состава среды: для увеличения ППЛ требовалось повысить относительную долю калия за счет азота и фосфора, а площади листьев, приходящейся на единицу сухой массы, — снизить долю калия и повысить долю азота. Однако взаимная компенсация этих параметров приводила в условиях действия стрессового фактора к стабилизации такого показателя, как «доля листьев в общей массе растения».
Аналогичным образом можно проанализировать действие кислотности почвы и отдельных форм азотных удобрений на изменение потребностей растений в макроэлементах, необходимое для максимальной активности фотосинтетического аппарата. При этом становится понятным, что уменьшение поглощения из почвы того или иного элемента минерального питания может быть обусловлено не только нарушением процессов переноса их через стенки и мембраны клеток, но и изменениями физиолого-биохимического плана, то есть переключением программ метаболизма и катаболизма на шунтовые механизмы, обеспечивающие иные потребности в элементах для достижения той же конечной цели.
Р е а к ц и я р а с т е н и й н а э д а ф и ч е с к и й с т р е с с в з а в и с и м о с т и о т с в о й с т в п о ч в ы. По данным оценки агрохимических свойств почвы на участках Фаленской селекционной станции НИИСХ Северо-Востока (80), выявлена различная вариабельность эдафических факторов по горизонтам кислой и нейтральной почвы (табл. 2).
2. Оценка агрохимических свойств кислой и нейтральной почвы на участках Фа-ленской селекционной станции НИИСХ Северо-Востока им. Н.В. Рудницкого (1999-2000 годы)
|
Глубина почвенного горизонта, см |
Влажность, % |
Обменная кислотность, мг-экв/100 г почвы |
Содержание, мг/100 г почвы |
рН |
Содержание, мг/кг |
Срок отбора пробы |
||
|
Н+ |
1 А13+ |
К 2 О 1 |
Р 2 О 5 |
|||||
|
Ап, 0-20 |
16,1 |
Н е й т 0,45 |
р а л ь н а я п о ч в а 0,45 0 |
5,97 |
214 |
124 |
Май |
|
|
21,6 |
0,18 |
0,18 |
0 |
6,48 |
314 |
100 |
Июнь |
|
|
13,6 |
0,18 |
0,18 |
0 |
6,71 |
253 |
83 |
Июль |
|
|
21,0 |
0,18 |
0,18 |
0 |
6,64 |
287 |
123 |
Август |
|
|
А2В, 20-40 |
21,3 |
23,28 |
23,10 |
0,18 |
3,87 |
126 |
27 |
Май |
|
19,2 |
7,69 |
7,65 |
0,04 |
4,22 |
91 |
38 |
Июнь |
|
|
24,1 |
1,40 |
1,35 |
0,05 |
4,72 |
180 |
43 |
Июль |
|
|
24,1 |
2,07 |
2,02 |
0,05 |
4,36 |
195 |
51 |
Август |
|
|
В, 40-60 |
23,8 |
43,50 |
43,21 |
0,29 |
3,71 |
111 |
14 |
Май |
|
24,8 |
23,81 |
23,67 |
0,14 |
3,86 |
129 |
27 |
Июнь |
|
|
22,2 |
25,01 |
24,80 |
0,21 |
3,89 |
163 |
31 |
Июль |
|
|
26,6 |
27,29 |
27,13 |
0,16 |
3,79 |
226 |
43 |
Август |
|
|
Ап, 0-20 |
16,0 |
К и с л а я 24,15 0,03 |
п о ч в а 24,12 |
3,98 |
84 |
35 |
Май |
|
|
19,6 |
20,93 |
0,05 |
20,88 |
3,93 |
64 |
44 |
Июнь |
|
|
21,6 |
20,73 |
0,03 |
20,70 |
3,84 |
62 |
47 |
Июль |
|
|
20,0 |
20,24 |
0,08 |
20,16 |
3,84 |
68 |
55 |
Август |
|
|
А2В, 20-40 |
21,3 |
23,55 |
0,06 |
23,49 |
3,93 |
100 |
21 |
Май |
|
20,7 |
20,84 |
0,05 |
20,79 |
3,91 |
82 |
35 |
Июнь |
|
|
17,5 |
23,75 |
0,08 |
23,67 |
3,90 |
76 |
56 |
Июль |
|
|
19,4 |
25,46 |
0,08 |
25,38 |
3,81 |
85 |
49 |
Август |
|
|
В, 40-60 |
24,0 |
19,84 |
0,13 |
19,71 |
3,95 |
109 |
17 |
Май |
|
22,6 |
20,25 |
0,09 |
20,16 |
3,93 |
94 |
21 |
Июнь |
|
23,5 |
19,13 |
0,12 |
19,01 |
3,91 |
81 |
52 |
Июль |
|
29,2 |
21,51 |
0,93 |
20,58 |
3,82 |
121 |
56 |
Август |
На участке с кислой почвой вариабельность эдафических факторов по сезонам года была невысокой; как по срокам отбора проб, так и по горизонтам почвы не отмечено вариабельности таких показателей, как обменная кислотность, рН и содержание подвижных ионов алюминия. При увеличении глубины горизонта кислотность почвы нейтрального участка (обменная кислотность, содержание ионов водорода и алюминия, рН почвенного раствора) возрастала, варьируя по срокам отбора образцов. Следует отметить, что содержание токсичных для растений ионов алюминия было высоким до конца июня в подпахотном слое, который играет существенную роль в формировании продуктивности сельскохозяйственных культур (81).
Если учесть, что корни яровых зерновых культур в почвах исследуемой зоны располагаются в основном в слое 0-40 см и при этом растут вертикально вниз, то становится ясно, что еще до закладки узловых корней эти растения испытывают на себе действие стрессового фактора (82, 83). Постепенно действие алюминия уменьшается, рН почвенного раствора несколько повышается. При этом метаболизм корней растений, находящихся в условиях напряженности стрессового фактора, должен постоянно меняться. Коэффициенты парных корреляций между урожаем зерна с 1 м2 и числом зерен в метелке для растений 26 сортов овса, оцениваемых в течение 5 лет, на кислых почвах были статистически достоверными по годам исследований (Р < 0,05 и выше), а на нейтральных почвах — существенно колебались и далеко не всегда были достоверными (84). Коэффициенты парных корреляций между урожайностью и массой 1000 зерен в те же годы существенно варьировали как по абсолютной величине, так и по направленности на обоих участках, хотя в вариантах на кислой почве почти всегда были выше, чем на нейтральной. В то же время на число зерен в метелке почвенные факторы оказывали очень сильное влияние, так как семенные бугорки на конусе нарастания метелки закладываются на ранних этапах развития растений.
Число зерен в метелке и масса 1000 зерен — это две составляющие урожая растений овса, которые формируются в онтогенезе не одновременно, причем сила и направление их взаимодействия могут быть использованы для предварительной оценки характера взаимодействия стрессовых факторов среды (85). Коэффициенты парных корреляций позволяют предположить наличие двух из пяти предложенных ситуаций (86): один лимитирующий фактор при увеличении жесткости действует в течение всего вегетационного периода на два признака, формирующиеся не одновременно; при аналогичной ситуации распределение действия последующего лимитирующего фактора зависит от предыдущего. В первой ситуации коэффициенты корреляции теоретически должны быть очень низкими, вплоть до отрицательных значений. Однако в полевых условиях Фаленской селекционной станции нам не удалось отметить существенного увеличения кислотности почвенного раствора и содержания подвижных ионов алюминия в период вегетации. Во втором случае если распределение действия второго фактора положительно зависит от такового первого, то коэффициенты корреляций будут также положительными, и наоборот.
В нашем случае коэффициенты парных корреляций между числом зерен в метелке и массой 1000 зерен были отрицательными в девяти случаях из десяти (исключение — кислая почва, 1997 год). Это подтверждает наличие в течение 5 лет в полевых условиях нескольких лимитирующих рост факторов, сменяющих друг друга в онтогенезе растений овса.
Суммарное влияние стрессов различных типов и их сочетания на каждый отдельно взятый генотип можно оценивать на основе графического отображения работы соответствующих генетических систем (87). Для этого рассмотрим распределение сортов овса в предлагаемых автором двумерных признаковых координатах масса соломины—масса зерна для главного стебля или в пересчете на 1 м2 (рис.). Эти признаковые координаты будут отражать изменение важнейшего свойства растения — аттракции пластических веществ к растущим семенам, что обусловливает массу 1000 зерен (крупность зерна). Известно, что аттрагирующая способность существенно 47
различается у разных генотипов в зависимости от условий выращивания (88). Исходя из этих графиков, можно рассчитать количественный сдвиг в работе системы адаптивности каждого сорта и проранжировать рассматриваемые сорта в порядке уменьшения (увеличения) адаптивного потенциала.
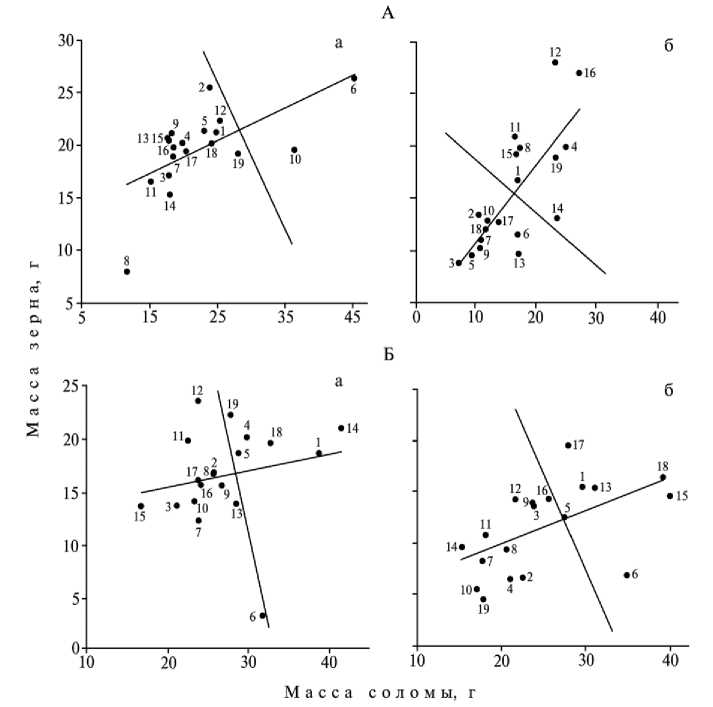
Распределение различных сортов овса (1-19) в двумерных признаковых координатах (масса соломы— масса зерна) по годам исследования и в зависимости от кислотности почвы: А и Б — соответственно 1999 и 2000 годы; а и б — соответственно нейтральная и кислая почвы; 1 — 1L86-5698, 2 — Centinial, 3 — Dolphin, 4 — Hja 78152, 5 — Meguin, 6 — Orlando, 7 — Shiromi, 8 — Гарант, 9 — Дорис, 10 — И-1987, 11 — И-2190, 12 — И-2363, 13 — И-2478, 14 — И-2492, 15 — Овен, 16 — Траверс, 17 — Улов, 18 — Факир, 19 — Чиж.
Один и тот же сорт в разные годы может испытывать различные по жесткости и продолжительности действия стрессы, что и приводит к сдвигам в работе сложной генетической системы адаптивности. Ввиду различий на генетическом уровне одни и те же параметры среды выращивания для одних сортов будут стрессовыми, для других — нет. Иными словами, условия года будут способствовать либо проявлению адаптивного потенциала сорта, либо — наоборот. Те же рассуждения можно применить и к перераспределению биомассы между колосом и вегетативной частью растения. Как видно из графиков, условия одного года могут способствовать перекачке пластических веществ в колос, а условия другого — препятствовать им (см. рис.).
Из всех рассмотренных методов оценки реакции растений на стрессовые эдафические факторы нам представляется наиболее подходящим и простым в использовании именно графический метод, предложенный Драгавцевым. С одной стороны, это объясняется тем, что между величиной сдвига в работе генетических систем, оцениваемых по распределению биомассы между частями растений, и всеми 13
элементами структуры урожая существует достоверная корреляционная зависимость в оба года исследований, причем в большей степени это относится к генетической системе адаптивности растений, чем системе аттракции пластических веществ. С другой стороны, при Р < 0,05 можно получить уравнения множественной регрессии, которые бы связывали воедино показатели сдвигов в работе генетических систем пластичности и адаптивности растений, элементов структуры урожая и лабораторной оценки кислотоустойчивости испытанных сортов овса (ИДК): I — величина сдвига в работе генетических систем адаптивности равна –210,7 – 0,83 (длина метелки) + 0,81 (высота растений) + 1,62 (число зерен в метелке) – 1,39 (масса зерна с одной метелки) + 1,16 (масса 1000 зерен) + 0,41 (число растений ко времени уборки) – 0,21 (число стеблей) + 0,74 (масса снопа) – 0,46 (масса зерна с одного снопа) + 0,54 (ИДК); II — величина сдвига в работе генетических систем пластичности равна 97,83 – 0,61 (длина метелки) + 0,21 (число зерен) – 0,46 (масса зерна с одной метелки) – 0,09 (число растений ко времени уборки) – 0,37 (масса снопа) + 0,43 (масса зерна с одного снопа) + 0,25 (урожай с 1 м2) – 0,28 (ИДК). Уравнение I позволяет проанализировать до 97,5 % изменчивости результирующего признака, уравнение II — до 85,6 %.
В то же время между показателями сдвигов в работе генетических систем адаптивности и пластичности и ИДК конкретных сортов овса наблюдается далеко неоднозначная взаимосвязь. При расчете коэффициентов парных корреляций между рассматриваемыми параметрами выявлено отсутствие статистически значимых взаимосвязей. Это свидетельствует о существовании как сортов, у которых повышение адаптивности и пластичности, оцениваемой по развитию надземных органов, коррелирует с повышенной устойчивостью корневых систем к действию стресса, так и сортов, у которых наблюдается обратная зависимость между устойчивостью надземных и подземных органов, а также сортов, у которых их развитие находится в относительной независимости друг от друга.
Содержание в почве различных химичес-ких элем е н т о в. Кроме кислотности почвы, связанной в первую очередь с высоким содержанием подвижных форм алюминия, необходимо остановиться и на таком аспекте проблемы, как наличие в почве других химических микроэлементов, которые могут оказывать существенное влияние на рост и развитие растений. В настоящее время ни у кого уже не возникает сомнений в том, что антропогенные факторы в биогеохимическом круговороте многих элементов стали сопоставимы с природными, а порой и превосходят их. Например, накопление тяжелых металлов в верхних горизонтах почвы промышленных регионов мира может значительно превышать фоновые концентрации в десятки и сотни раз (6, 89). Ежегодное техногенное поступление наиболее опасных тяжелых металлов — свинца и кадмия — в окружающую среду Кировской области составляет соответственно 451,64 и 4,5 т (90). Почва служит мощным аккумулятором тяжелых металлов и практически не теряет их со временем. Особенно прочно фиксируют тяжелые металлы гумусосодержащие почвенные горизонты, то есть наиболее плодородный слой. Опасность усугубляется тем, что химическое загрязнение может не проявляться длительное время в силу высокой буферной способности почвы.
Многие относительно «чистые» территории Кировской области (лесные, луговые и пахотные почвы северных, восточных и южных районов), несмотря на свою удаленность от промышленных объектов, содержат в почве подвижные фракции некоторых тяжелых металлов в количествах, превышающих ПДК. Хотя среднее содержание подвижных форм тяжелых металлов не вызывает беспокойства, максимальные показатели свидетельствуют об опасности загрязнения (табл. 3). При расчете коэффициентов парных корреляций между содержанием в почве подвижных форм тяжелых металлов, с одной стороны, рН сол. и содержанием общего органического углерода (С общ. ), с другой, не выявлено каких-либо четких закономерностей. Только для Ni отмечены достоверные отрицательные взаимосвязи между содержанием в почве и рН ( r = –0,53), а также Собщ. ( r = –0,35).
3. Содержание подвижных форм тяжелых металлов в гумусовых горизонтах различных почв Кировской области (мг/кг почвы)
|
Микроэлемент |
Дерново-подзолистая почва |
Серая лесная почва |
Пойменная почва |
|||
|
среднее |
^ от-до |
среднее |
от-до |
среднее 1 |
от-до |
|
|
Cu |
2,02 |
0,70-4,15 |
2,15 |
0,99-5,10 |
1,31 |
0,58-2,10 |
|
Ni |
0,39 |
0,11-2,44 |
0,32 |
0,17-0,80 |
0,53 |
0,30-0,78 |
|
Cd |
0,10 |
0,02-0,29 |
0,22 |
0,03-1,48 |
0,18 |
0,08-0,35 |
|
Pb |
1,86 |
0,45-3,10 |
1,48 |
0,42-3,10 |
1,11 |
0,75-2,11 |
|
Zn |
3,17 |
0,85-6,11 |
3,04 |
1,25-6,10 |
4,58 |
3,04-7,20 |
|
Mo |
0,26 |
0,09-0,56 |
0,15 |
0,05-1,41 |
0,25 |
0,12-0,60 |
|
Fe |
158,86 |
11,70-284,20 |
250,74 |
129,17-369,50 |
133,66 |
68,20-250,10 |
|
Cr |
2,62 |
0,18-10,50 |
2,66 |
0,60-9,88 |
0,66 |
0,20-1,35 |
|
Mn |
43,86 |
7,60-135,40 |
59,85 |
21,60-91,50 |
90,64 |
22,40-130,20 |
Достоверная положительная корреляция наблюдалась между содержанием в почве отдельных металлов по группам Cu-Cr-Mn-Cd и Cd-Pb-Cr, что позволяет говорить о сходстве подвижных форм этих элементов. По нашим предварительным данным, в зависимости от вида и типа стратегии адаптации распределение и накопление тяжелых металлов в биомассе растений происходит по разным схемам. Например, для клевера лугового получены достоверные корреляции между содержанием Cr и Mn в почве и растении — соответственно r = 0,93 и r = –0,82, а содержание Cd и Pb зависело от Собщ. — соответственно r = 0,78 и r = –0,67; для растений бодяга полевого таких взаимосвязей не обнаружено.
Взаимосвязь между содержанием различных химических элементов и Собщ. служит показателем гумусного состояния почвы. Известно, что гумус в значительной мере является источником питания для растений и энергетической основой биологических процессов. Гуминовые кислоты, выступающие в роли комплексообразовате-лей, способствуют перераспределению веществ в почвенном профиле, во многом определяя направление почвообразовательного процесса. В течение года содержание, а главное, состав гумуса могут значительно изменяться (91). Пономарева отмечала, что сезонные колебания содержания гумуса не так значимы, как изменения его качественного состава (92). Содержание подвижных форм органического вещества почвы варьирует в течение вегетационного периода, обусловливая динамику ряда других показателей (93).
По нашим данным, на полях НИИСХ Северо-Востока (суглинистая дерновослабоподзолистая почва) содержание общего углерода не различается по горизонтам почвы в зависимости от дозы внесенных удобрений (контроль, N60P60K60 и N100P100K100 под посев ячменя). В пахотных горизонтах практически отсутствует сезонная динамика по содержанию органического углерода. По мнению ряда авторов, внесение минеральных удобрений способствует стабилизации содержания гумуса, а распашка почвы без внесения удобрений ведет к минерализации последнего (94).
Максимальное содержание лабильных гумусовых веществ (С лаб. ) наблюдается в пахотных горизонтах осенью. Очевидно, минерализация органического вещества, идущая в первую очередь за счет лабильных форм последнего, компенсируется поступлением органических соединений при гумификации пожнивных и корневых остатков. Содержание Слаб. в подпахотных горизонтах практически не отличалось по вариантам внесения удобрений. Полученные данные свидетельствуют о подкисляющем действии удобрений на пахотные горизонты.
При анализе лабильного органического вещества суглинистой дерновослабоподзолистой почвы оказалось, что содержание Ca, Mg и Al зависит в наибольшей степени от срока отбора проб и почвенного горизонта, а также сочетания этих факторов, а содержание Ni, Zn и Cu — от срока отбора проб.
При расчете коэффициентов парных корреляций между содержанием микроэлементов, составом и содержанием органического вещества, а также кислотностью почвы выявлен ряд закономерностей. Так, между С общ. и содержанием Cu нет взаимосвязи, а между последним, рНсол. и Слаб. наблюдалась прямая корреляция, которая ослабевала при увеличении дозы удобрений. Коэффициенты корреляции между содержанием Zn и Ni, с одной стороны, и Слаб., с другой, оказались несущественными; между рНсол. и содержанием Ni составляли соответственно 0,70, 0,82 и 0,54, а для Zn 50
-
— 0,42, 0,38 и 0,24 в зависимости от дозы удобрений (контроль, N 60 P 60 K 60 и N 100 P 100 K 100 ). Корреляции между содержанием Al и величиной обменной кислотности были незначительными и повышались при увеличении дозы удобрений; в то же время между содержанием Al и Слаб. наблюдалась высокая корреляционная зависимость — r = 0,70.
Остальные микроэлементы по своим свойствам либо обнаруживали сходство (Са с Cu; Mg с Zn и Ni), либо существенные различия (Ca с Zn и Ni). Очевидно, это было связано с влиянием на подвижность элементов наличия подстилающих карбонатных пород на опытных полях.
К о р н е в ы е г н и л и к а к э д а ф и ч е с к и й с т р е с- с о в ы й ф а к т о р. Почву заселяют различные микроорганизмы, в том числе микромицеты, играющие существенную роль в процессах почвообразования, вызывая заболевания многих растений. В почвах агроэкосистем в отличие от естественных ценозов численность патогенных популяций превышает пороги вредоносности, причем вредители обладают высокой изменчивостью (95). В экстремальных условиях среды, обусловленных действием как абиотических, так и биотических факторов, возникают изменения генотипической структуры микроорганизмов естественных популяций. При этом возрастает вероятность появления мутантов, обладающих большей паразитической специализацией, агрессивностью и вредоносностью, чем исходные формы (96). В последние десятилетия прошлого столетия корневые гнили повсеместно наносили ощутимый вред сельскому хозяйству: ежегодные потери урожая достигали 30 % (97-101).
Для оздоровления почв от возбудителей корневых гнилей разработан ряд агротехнических методов: фитосанитарные севообороты, что исключает повторное возделывание восприимчивых культур; правильный подбор фитосанитарных предшественников; сбалансированное внесение органических и минеральных удобрений. Однако взаимодействие растение-хозяин—патоген—среда имеет более сложный характер, причем корневые гнили выступают и как эдафический стрессовый фактор.
Корневые гнили — общее наименование группы распространенных болезней растений, вызываемых, как правило, комплексом полупаразитарных микроорганизмов и имеющих сходные внешние симптомы поражения (99). Возбудителями корневой гнили являются грибы, принадлежащие к следующим родам: Fusarium, Bipolaris, Gaemonnomyces, Cercosporella, Typhula, Rhizoctonia, Aureobasidium, Alternaria, Py-thium . По частоте встречаемости и вредоносности приоритет принадлежит грибам рода Fusarium и Bipolaris . Заболевание может проявляться в течение всего периода вегетации растений, вызывая гибель всходов, отставание растений в росте, щуплость зерна и отмирание стеблей. Грибы — возбудители корневой гнили обладают широким набором различных ферментов, разрушающих ткани растений, что обусловливает гибель последних (102). Для большинства патогенов этой группы характерны высокая генетическая изменчивость и вся гамма переходных форм от сапрофита до факультативного паразита (103).
Патогены, вызывающие корневую гниль, поражают не только подземные, но и приземные органы злаков (98). Болезнь внешне проявляется в виде побурения корней, подземного междоузлия, узла кущения, основания стебля и влагалища нижних листьев, которые теряют прочность, становятся рыхлыми, хрупкими и обламываются при выдергивании растений из почвы (103). При этом корни растений, как правило, сильнее поражаются фузариозными, а основания стеблей — гельминтоспориозными грибами. В агробиоценозах в ризосфере корней растений локализуются возбудители корневой гнили, принадлежащие к разным видам (смешанная инфекция) (104).
К возбудителям гельминтоспориозной корневой гнили относятся виды грибов, входящие в прежний сборный род Helminthosporium Lk. Fr. Из этого комплексного формального рода выделилось несколько самостоятельных родов — Drechslera, Bipolaris, Curvularia . Возбудителями фузариозной гнили являются грибы рода
Fusarium Lk. Fr., которые заселяют различные растительные остатки, ризосферу и корни, а также активно размножаются на мертвых корнях, выдерживая конкуренцию при заселении субстрата другими грибами, бактериями и актиномицетами (105). Эти 51
возбудители способны существовать в почве в сапротрофной форме и накапливаться на растительных остатках, переходя при определенных условиях к паразитной форме существования, что делает борьбу с этой болезнью весьма сложной.
Адаптационный потенциал растений к корневым гнилям недостаточно исследован. Как отмечал Н.И. Вавилов, источники иммунитета и устойчивости к грибным болезням следует выявлять в местах сопряженной эволюции растения-хозяина и паразита (106). По мнению А.А. Гончаренко, основой стратегии селекции зерновых культур должно быть создание не высокоустойчивых, а толерантных к корневой гнили сортов (107). Необходима длительная и целенаправленная селекция на создание исходного материала и сортов, способных формировать урожай при поражении патогенами. Приоритетным направлением селекции считается создание сортов, экологически устойчивых к нерегулируемым факторам внешней среды и наиболее вредоносным заболеваниям, максимально адаптированных к почвенно-климатическим особенностям Северо-Восточного региона России (108).
В результате многолетних обследований посевов зерновых культур и изучения структуры популяций возбудителей корневой гнили, проводи-
4. Частота встречаемости наиболее распространенных возбудителей корневой гнили на различных зерновых культурах (%)
|
Культура |
Fusarium culmorum |
Fusarium oxysporum |
Fusarium heterosporum |
Bipolaris sorokiniana |
|
Пшеница |
42,2 |
27,0 |
22,3 |
1,2 |
|
Рожь |
36,5 |
8,5 |
12,1 |
1,5 |
|
Ячмень |
25,1 |
16,6 |
7,5 |
32,7 |
мых сотрудниками ВНИИ фитопатологии в СевероВосточном регионе, показано, что среди патогенов, вызывающих заболевания озимой ржи, доминирующее положение занимают грибы рода Fusarium : F. culmorum, F. oxysporum, F. heterosporum, F.
sporotrichiella, F. nivale, F. gibbosum, F. avenaceum, F. moniliforme, F. sambucinum, F. semitectum, F. solani , а также Bipolaris sorokiniana . Наибольшая доля в патогенных комплексах зерновых культур принадлежит видам F . culmorum , F. oxysporum , F. heterosporum и B. sorokiniana (табл. 4).
Соотношение видов в популяции вредителей меняется по годам и зонам. Например, в посевах озимой ржи, занимающих около 30 % всех площадей зернового клина в Кировской области, в 1995, 1998, 2001 и 2002 годах наряду с F. culmorum широкое распространение получили возбудители F. oxysporum , в 1996 году — F. culmorum и F. heterosporum, в 1997 году — F. culmorum, B. sorokiniana . Различалось также соотношение представителей разных видов возбудителей корневой гнили по зонам Кировской области: на севере преобладали популяции F. oxysporum , F. sporotrichiella , B. sorokiniana и F. culmorum ; в центральной части — F. culmorum , F.
oxysporum и F. heterosporum (табл. 5).
По данным лабораторных исследований, пластичность и совместимость видов между собой обусловливают высокую адаптационную способность вредителей (109-112). Показано, что штаммы внутри каждого вида характеризуются разной способностью к токсинообразованию. При достаточной влажности и высокой температуре изоляты фузариозных грибов ряда видов ( F. graminearum, F. avenaceum, F. culmorum ) продуцируют дезоксиниваленол (ДОН) и его производные (3-ацетил-ДОН, 15-ацетил-ДОН, 7-дезокси-ДОН), а также токсины из группы бутенолидов — 5-ацетамидо-2(5Н)-фуранона-(5-ААФ).
Для первичной оценки образцов на устойчивость к отдельным видам возбудителей используют модифицированный экспресс-метод Бенкена (113), позволяющий проводить массовый скрининг и отбор слабопоражаемых генотипов. В основу метода положена различная реакция сортов на споровую суспензию или фильтрат культуральной жидкости (ФКЖ) в период прорастания семян. Образцы дифференцируют по доле проросших семян, числу корней, размеру проростков и корней. Устойчивость сортообразцов ржи к споровой суспензии определяют в лабораторных условиях, споровую нагрузку подбирают для каждого вида возбудителя отдельно (114, 115). По данным этих опытов, под действием токсинов уменьшается доля проросших семян и число корней, замедляется рост первичного корня и побегов.
При сравнительном анализе устойчивости различных сортов злаков к споровой суспензии возбудителя F. culmorum и ФКЖ выявлена высокая корреляционная связь между показателями, полученными соответственно в полевых и лабораторных условиях ( r = 0,65). Для оценки и отбора сортов, устойчивых к возбудителям корневой гнили, можно использовать споровую суспензию наиболее патогенных штаммов. При сравнительном анализе влияния токсинов ФКЖ и споровой суспензии грибов F. culmorum на развитие проростков озимой ржи ряда сортов (Татарская 1, Кировская 89, Вятка 2, Крона, Дымка, Радуница и Восход, популяция Ф14-92), разли-
-
5. Видовой состав возбудителей корневой гнили озимой ржи по годам исследования и зонам возделывания культуры в Кировской области (%)
Год
Fusarium culmorum
Fusarium oxysporum
Fusarium heterosporum
Fusarium avenaceum
Fusarium gibbosum
Fusarium sporotrihiella
Fusarium sambucinum
Fusarium semitectum
Fusarium moniliforme
Fusarium nivale
Bipolaris soro-kiniana
1995
1996
1997
1998 Всего
10,5
40,0
5,0
С е в е р н а я з о н а в о з д е л ы в а н и я 12,5 13,5 50 12,5
0,2
12,5
4
2,5
13,6
13,6
9,0
–
–
4,5
–
–
–
40
18,2
9,9
34,7
8
6,2
6,7
27,2
6,2
0,1
6,2
22
10,5
1995
13
29
18
Ц е н
5
т р а л ь н а я 6
з о н а в о з д е л ы в а н и я
6 12
–
5
–
6
1996
59,6
2,3
12,8
3,5
3,5
3,2
1,8
–
–
–
8,6
1997
38,4
10,5
7,6
11,5
13,5
13,5
0,9
–
–
5,2
10,6
1998
30,6
20,9
10,7
10,8
11,8
11,8
2
1,1
–
–
9,8
Всего
35,4
15,7
12,3
7,7
8,6
3,5
4,1
0,3
1,3
1,3
8,7
1995
78,3
15,8
1,1
Ю ж н а я з о н а в о з д е л ы в а н и я 1,5 5,4 0,4 1,8
–
0,7
–
–
1996
64,9
0,5
25,7
1
0,5
–
2,6
–
–
–
3,4
1997
50
–
–
50
–
–
–
–
–
–
–
1998
41,5
18,1
17,5
4,3
4,1
1,5
2,2
0,4
–
–
10,6
Всего
50,8
10
14,2
14,2
2,1
0,4
1,6
0,2
0,2
–
3,5
П р и м е ч а н и е. Прочерк означает отсутствие возбудителей заболевания.
чающихся по устойчивости к этому возбудителю, была выявлена аналогичная реакция растений по основным показателям.
Регулярные обследования посевов и анализ видового разнообразия комплексов патогенов и степени агрессивности последних позволяют составлять фитопатологические прогнозы и предотвращать эдафические стрессовые факторы, вызываемые корневыми гнилями.
Заключение
Данные литературы и результаты наших собственных исследований свидетельствуют о том, что наиболее существенное влияние на продуктивность сельскохозяйственных растений на Северо-Востоке европейской части России оказывают следующие факторы: повышенная кислотность почвы; большое количество подвижных ионов алюминия; несбалансированность минерального питания растений; повышенная подвижность ионов тяжелых металлов; значительное распространение корневых гнилей растений. Формы азотных удобрений, соотношение N:P:К, сезонная динамика физико-химических свойств почвы значительно влияют на полевую оценку степени адаптированности растений к кислым почвам региона. Особенно наглядно это проявляется по качественным и количественным сдвигам в работе соответствующих генетических систем растений. По данным, полученным в различных научных учреждениях мира, в том числе и России, растения, хорошо приспособленные к условиям внешней среды, оказываются более устойчивыми и к болезням. Сочетание в сортах устойчивости к биотическим и абиотическим стрессовым факторам — один из возможных способов создания долговременных источников устойчивости (116).
Таким образом, при селекции на эдафическую устойчивость растений необходимо: во-первых, выявлять количество полевых стрессовых фaкторов и оценивать их сезонную динамику; во-вторых, при выборе полевых участков (для контроля и опыта) использовать методически правильный подход; в-третьих, параллельно оценивать устойчивость селекционного материала к нескольким основным стрессовым факторам в контролируемых условиях лабораторных и вегетационных опытов; в-четвертых, усилить работы по экологической генетике основных сельскохозяйственных культур Северо-Востока европейской части России.
Л И Т Е Р А Т У Р А
-
1. Ж у ч е н к о А.А. Проблемы адаптации в современном сельском хозяйстве. С.-х. биол., 1993, 5: 3-35.
-
2. Б е л я к о в В.С., М о л о д к и н В.Н. Состояние плодородия почв сельскохозяйственного назначе
ния Кировской области. В сб.: Повышение устойчивости земледелия в современных условиях. Киров, 2001.
-
3. К о п ы с о в И.Я. Изменение качества почв под влиянием длительного сельскохозяйственного использования в условиях Кировской области. В сб.: Повышение устойчивости земледелия в современных условиях. Киров, 2001.
-
4. О состоянии окружающей природной среды Кировской области в 1997 году. Регион. докл. Киров, 1998.
-
5. Д о к у ч а е в В.В. К учению о зонах природы. СПб, 1899.
-
6. К а б а т а - П е н д и а с А., П е н д и а с Х. Микроэлементы в почвах и растениях. М., 1989.
-
7. Р о д и н а Н.А. Оценка исходного материала ячменя на устойчивость к повышенной кислотности и
алюминию. Тез. докл. V съезда ВОГиС. М., 1987, 4, 2: 123.
-
8. I n o u e K. Active Al and Fe components in acid soils and related soils. Grans. 14 Int. Cong. Soil Sci., Kyoto, 1990, 7: 153-158.
-
9. R e n g e l Z., E l l i o t t D. Mechanism of aluminum inhibition of net 45Ca2+ uptake by Amaranthus protoplasts. Plant Physiol., 1992, 98, 3: 632-638.
-
10. R y a n P.R., D i t o m a s o J.M., K o c h i a n L.V. Aluminum toxicity in roots: an investigation of spatial sensitivity and the role of the root cap. J. Exp. Bot., 1993, 44, 2: 437-446.
-
11. W a g a t s u m a T. Characterization of absorption sites for aluminum in the roots. Soil Sci. Plant Nutr., 1983, 29, 4: 499-515.
-
12. H u a n g J.W., G r u n e s D.L., K o c h i a n L.V. Aluminum effects on calcium (45Ca2+) translocation in aluminum-tolerant and aluminum-sensitive wheat ( Triticum aestivum L .) cultivars. Differential responses of the root apex versus mature root regions. Plant Physiol., 1993, 102, 1: 85-93.
-
13. R e n g e l Z. Disturbance of cell Ca2+ homeostasis as a primary trigger of Al toxicity syndrome. Plant Cell Environ., 1992, 15, 4: 931-938.
-
14. R y a n P.R., K o c h i a n L.V. Aluminum differentially inhibits calcium uptake into the root apex of near-isogenic lines of wheat: a possible mechanism of toxicity. Plant Physiol., 1993, 102, 4: 975-982.
-
15. A l v a A.K., E d w a r d s D.G., A s h e r C.J. e.a. Effect of phosphorous/aluminum ratio and calcium concentration on plant response to aluminum toxicity. Soil Sci. Soc. Am. J., 1986, 50, 1: 133-137.
-
16. B l a m e y F.P.C., E d w a r d s D., A s h e r C. Effects of aluminum, OH:Al and P:Al molar ratios, and ionic strength on soybean root elongation in solution culture. Soil Sci., 1983, 136, 4: 197-207.
-
17. M a r w a h a B.C., S o o d R. Effect of partially acidulated rock phosphate in minimizing the P-fixing capacity of an acid affisol. J. Indian Soc. Sci., 1989, 32, 2: 333-336.
-
18. К л и м а ш е в с к и й Э.Л. Генетический аспект минерального питания растений. М., 1991.
-
19. H a u g A. Molecular aspects of aluminum toxicity. C. R. C. Crit. Rev. Plant Sci., 1984, 1, 1: 345-373.
-
20. V o n U e x k u l l H.R., M u t e r t E. Global extend, development and economic impact of acid soils. Plant
Soil, 1995, 171, 1: 1-15.
-
21. F o y C.D. Effect of aluminum on plant growth. In: Plant root and its environment. Charlottesville, 1973.
-
22. K a m p r a t h E.J., F o y C.D. Lime-fertilizer-plant interactions in acid soils. Fertilizer technology and use.
-
23. M c L e a n E.O. Chemistry of soil Al. Commun. Soil Sci. Plant. Anal., 1976, 7, 3: 619-636.
-
24. O l m o s I.L.J., C a m a r g o M.N. Incidence of aluminum toxicity in Brazilian soils: its characterization
and distribution. Ciencia e Cultura, 1976, 28, 1: 171-180.
-
25. S i l v a A.R. Melhoramento genetica para resistencia a toxidez de aluminio e manganes no Brazil; Antece-dentes, necessidade e possibilidades. Topica para discussao e pesquisas. Cienc. Cult (San Paulo), 1976, 28, 1: 147-149.
-
26. П а й в и н С.Г., Н о в о с е л о в М.Ю. Некоторые результаты селекции клевера лугового на устойчивость к ионам алюминия. Сел. и сем., 1997, 2: 23-25.
-
27. F o y C.D. Effects of soil calcium on plant growth. In: Plant root and its environment. Charlottesville, 1974.
-
28. A n i o l A., G u s t a f s o n J.P. Chromosome location of genes controlling aluminum tolerance in wheat,
rye, and triticale. Can. J. Genet. Cytol., 1984, 26, 4: 701-705.
-
29. B u s s G.R., L u t z J.A., H a w k i n s G.W. Effect of soil pH and plant genotype on element concentra
tion and uptake by alfalfa. Crop Sci., 1975, 15, 3: 614-617.
-
30. C o n n e r A.J., M e r e d i t h C.P. Strategies for the selection and characterization of aluminum-resistant variants
from cell cultures of Nicotiana plumbaginifolia. Planta, 1985, 166, 3: 466-473.
-
31. C u l v e n o r R.A., O r a m R.N., F a z e k a s d e St. e.a. Variation in tolerance in Phalaris aquatica L.
-
32. D e v i n e T.E., F o y C.D., F l e m i n g A.L. e.a. Development of alfalfa strains with differential toler
ance to aluminum toxicity. Plant Soil, 1976, 44, 1: 73-79.
-
33. H a n s o n W.D., K a m p r a t h E.J. Selection for aluminum tolerance in soybeans based on seedling-root growth. Agron. J., 1979, 71, 4: 581-586.
-
34. R e i d D.A. Genetic control of reaction to aluminum in winter barley. Barley Genetics II – Proc. 2nd Int. Barley Genetics Symp., Pullman, Wash., 1971.
-
35. R h u e R.D., G r o g a n C.O., S t o c m e y e r E.W. e.a. Genetic control of aluminum tolerance in corn. Crop Sci., 1978, 18, 6: 1063-1065.
-
36. S t o l e n O., A n d e r s e n S. Inheritance of tolerance to low soil pH in barley. Hereditas, 1978, 88, 1: 101-105.
-
37. Б у т к е в и ч В.В. Приспособление растений к химическому составу среды. Сел. и сем., 1947, 6: 6376.
-
38. К о р н и л о в М.Ф., Б о р и с о в а Е.М., Т р у н и н а З.В. Известкование почвы и сорт (из работ Ленинградского отделения ВИУА). В сб.: Известкование дерново-подзолистых почв. Тр. ВИУА. М., 1955, 31: 202-250.
-
39. Б е р е с н е в а О.Л., Н и к и ф о р о в а Е.В., Т у м а с о в а М.И. Создание исходного материала клевера лугового с повышенной устойчивостью к кислотности почв. В сб.: Новые методы селекции и создания адаптивных сортов сельскохозяйственных культур: результаты и перспективы. Киров, 1998.
-
40. Р о д и н а Н.А. Направления адаптивной селекции и семеноводства в Северо-Восточном регионе РФ. В сб.: Научные основы стратегии адаптивного растениеводства Северо-Востока европейской части России. Киров, 1999, 2: 3-13.
-
41. Р о д и н а Н.А., С о л о д я н к и н а М.М. Реакция генотипов ячменя на стрессовые условия дерново-подзолистых почв. В сб.: Новые методы селекции и создания адаптивных сортов сельскохозяйственных культур: результаты и перспективы. Киров, 1998.
-
42. K o c h i a n L.V. Cellular mechanisms of aluminum toxicity and resistance in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 1995, 46, 2: 237-260.
-
43. Т а р ч е в с к и й И.А. Катаболизм и стресс у растений. 52-е Тимирязевское чтение. М., 1993.
-
44. У д о в е н к о Г.В. Механизмы адаптации растений к стрессам. Физиол. и биохим. культ. раст., 1979, 11, 2: 99-107.
-
45. L e s h e m Y.Y., K u i p e r P.J.C. Is there a GAS (general adaptation syndrome) response to various types of environmental stress? Biol. Plant., 1996, 38, 1: 1-18.
-
46. С а в и ч В.И. Некоторые закономерности ионного обмена в почвах. В сб.: Современные почвенные процессы. М., 1971.
-
47. П е р е в е р з е в В.Н. Сезонная динамика почвенных процессов в лесах Кольского полуострова. Тез. докл. Второго съезда общ-ва почвоведов. СПб, 1996.
-
48. С н а к и н В.В., К е с о в Е.Н. Биотический круговорот и процессы почвообразования. Мат. Междунар. симп. стран-членов СЭВ. Пущино, 1984.
-
49. A d a m s F., L u n d Z.F. Effect of chemical activity of soil solution aluminum on cotton root penetration of acid subsoils. Soil Sci., 1966, 101, 2: 193-198.
-
50. F o y C.D. General principles involved in screening plants for aluminum and manganese tolerance. In: Proc. workshop on plant adaptation to mineral stress in problem soils. Ithaca, N.Y., 1976.
-
51. B r o w n J.C., C l a r k R.B. Differential response of two maize inbreds to molybdenum stress. Soil Sci. Soc. Amer. Proc., 1974, 38, 2: 331-333.
-
52. A r m i g e r W.H., F o y C.D., F l e m i n g A.L. e.a. Differential tolerance of soybean varieties to an acid soil high in exchangeable aluminum. Agron. J., 1968, 60, 1: 67-70.
-
53. B r o w n J.C. Differential uptake of Fe and Ca by two corn genotypes. Soil Sci., 1967, 103, 2: 331-338.
-
54. M e d a p p a K.G., D a n a M.N. Tolerance of cranberry plants to manganese, iron, and aluminum. J. Am.
-
55. P r e s a d M. Nitrogen nutrition and yield of sugarcane as affected by N-Serve. Agron. J., 1976, 68, 2: 343346.
-
56. R o r i s o n I.H. The effect of extreme soil acidity on the nutrient uptake and physiology of plants. In: Acid sulphate soils. Wageningen, Netherlands, 1972.
-
57. S a n t a m a r i a P., E l i a A., P a p a G. e.a. Nitrate and ammonium nutrition in chicory and rocket salad plants. J. Plant Nutrit., 1998, 21, 9: 1779-1789.
-
58. T o w n s e n d L.R., B l a t t C.R. Lowbush blueberry: Evidence for the absence of a nitrate reduction system. Plant Soil., 1966, 25, 3: 456-459.
-
59. V a a s t P., Z a s o s k i R.J., B l e d s o e C.S. Effects of solution pH, temperature, nitrate/ammonium ratios, and inhibitors on ammonium and nitrate uptake by Arabica coffee in short-term solution culture. J. Plant Nutrit., 1998, 21, 7: 1551-1564.
-
60. V a n T u i l H.D.W. Organic salts in plants in relation to nutrition and growth. Agric. Res. Rep. Inst. Biol. Chem. Res. Field Crops and Herbage, Wageningen, Netherlands, 1965, 657: 1-52.
-
61. W i l t s h i r e G.H. Response of grasses to nitrogen source. J. Appl. Ecol., 1973, 10, 4: 429-435.
-
62. H a v i l l D.C., L e e J.A., S t e w a r t D.R. Nitrate utilization by species from acidic and calcareous soils.
-
63. G i g o n A., R o r i s o n I.H. The response of some ecologically distinct plant species of nitrate and to ammonium nitrogen. J. Ecol., 1972, 60, 1: 93-102.
-
64. F o y C.D., F l e m i n g A.L. The physiology of plant tolerance to excess available aluminum and manganese in acid soils. In: Crop tolerance to suboptimal land conditions. Madison, USA, 1978.
-
65. C l a r k R.B., B r o w n J.C. Differential phosphorus uptake by phosphorus-stressed corn inbreds. Crop Sci., 1974, 14, 4: 505-508.
-
66. F o y C.D., F l e m i n g A.L., S c h w a r t z J.W. Opposite aluminum and manganese tolerances in two wheat varieties. Agron. J., 1973, 65, 1: 123-126.
-
67. К л и м а ш е в с к и й Э.Л., Д е д о в В.М. Осаждение тканями корней Al – одна из причин генотипической специфики устойчивости растений к его токсичности. Докл. ВАСХНИЛ, 1977, 4: 7-9.
-
68. A n d r e w C.S., V a n d e n B e r g P.J. The influence of aluminum on phosphate sorption by whole plants and excised roots of some pasture legumes. Austr. J. Agric. Res., 1973, 24, 3: 341-351.
-
69. Д е д о в В.М., К л и м а ш е в с к и й Э.Л. О механизме генотипической специфики устойчивости растений к токсичности Al3+. Сиб. вест. с.-х. науки, 1976, 3: 13-16.
-
70. B i e l e s k i R.L. Enzyme changes in plants following changes in their mineral nutrition. Plant Anal. Fert. Probl. Abstr. 6th Int. Colloq., Tel Aviv, Israel, 1971.
-
71. A n d r e w C.S., J o h n s o n A.D., S a n d l a n d R.L. Effect of aluminum on the growth and chemical composition of some tropical and temperate pasture legumes. Austr. J. Agric. Res., 1973, 24, 3: 325-339.
-
72. C l a r k s o n D.T. Interactions between aluminium and phosphorus on root surfaces and cell wall material. Plant Soil, 1967, 27, 4: 347-356.
-
73. T a n K., K e l t j e n s W.G., F i n d e n e g g G.R. Evaluating the contribution of magnesium deficiency in the aluminum toxicity syndrome in twelve sorghum genotypes. Plant Soil, 1993, 149, 2: 255-261.
-
74. S o m e r s D.J., G u s t a f s o n J.P. The expression of aluminum stress induced polypeptides in a population segregating for aluminum tolerance in wheat ( Triticum aestivum L.). Genome, 1995, 38, 6: 1213-1220.
-
75. T a y l o r G.J. Aluminum tolerance is independent of rhizosphere pH in Triticum aestivum . Commun. Soil Sci. Plant Anal., 1988, 19, 6: 1217-1227.
-
76. W h e e l e r D.M. Differences in aluminum tolerance of ryegrass and white clover plants grown from seed or propagated vegetatively. J. Plant Nutrit., 1995, 18, 5: 949-954.
-
77. D u r i e u x P.P., J a c k s o n W.A., K a m p r a t h E.J. e.a. Inhibition of nitrate uptake by aluminium in maize. Plant Soil, 1993, 151, 1: 97-104.
-
78. S c h i e r G.A., M c Q u a t t i e C.J. Effect of water stress on aluminum toxicity in pitch pine seedlings. J. Plant Nutrit., 2000, 23, 5: 637-647.
-
79. В а х м и с т р о в Д.Б. Раздельное определение оптимумов суммарной дозы N+P+K и соотношения N:P:K в удобрении. Сообщ. 1. Постановка проблемы. Агрохимия, 1982, 4: 3-12.
-
80. Ш и х о в а Л.Н., Л и с и ц ы н Е.М. Динамика эдафических факторов на полях Фаленской селекционной станции. В сб.: Селекция, семеноводство и сортовая технология на Северо-Востоке европейской части России. Киров, 2001.
-
81. Л е б е д е в а Л.А., Ю л у ш е в И.Г., К а б а л и н а Л.Н. Значение подпахотного слоя в плодородии и продуктивности дерново-подзолистых почв, сформированных на элюво-делювии пермских карбонатных пород. В сб.: Повышение роли химизации при возделывании зерновых культур по интенсивной технологии. Киров, 1987.
-
82. К а л и н и н А.И., К у з н е ц о в Н.К. Селекция растений в связи с особенностями водного и химического состава дерново-подзолистых суглинистых почв восточной части ЕТС. В сб.: Селекция зер-
новых культур на устойчивость к болезням и неблагоприятным факторам среды в Волго-Вятском регионе. Тр. НИИСХ Северо-Востока. Киров, 1986.
-
83. Р о д и о н о в а Н.А., С о л д а т о в В.Н., М е р е ж к о В.Е. и др. Культурная флора. Овес. М., 1994, 2, 3: 1-367.
-
84. B t a l o v a G., L i s i t s y n E.M. Genotypic correlations in oat breeding for acid-resistance. Agriculture. Scientific Articles, 2002, 78, 2: 104-110.
-
85. Н е ч и п о р е н к о Н.Н., Д р а г а в ц е в В.А. О возможности прогноза уровней и знаков коэффициентов экологической корреляции. Генетика, 1986, 22, 4: 616-623.
-
86. Б а т а л о в а Г.А., Л и с и ц ы н Е.М. Генотипическая корреляция в селекции овса на кислотоустой-чивость. Докл. РАСХН, 2002, 4: 6-9.
-
87. Д р а г а в ц е в В.А. Алгоритмы эколого-генетической инвентаризации генофонда и конструирования сортов сельскохозяйственных растений по урожайности, устойчивости и качеству. СПб, 1997.
-
88. У д о в е н к о Г.В. Определение аттрагирующей способности зерна и ее компонентов у колосовых злаков. СПб, 1992.
-
89. Ч е р н ы х Н.А., М и л а щ е н к о Н.З., Л а д о н и н В.Ф. Экотоксикологические аспекты загрязнения почв тяжелыми металлами. М., 1999.
-
90. Б у р к о в Н.А. Оценка поступления ртути, свинца и кадмия в окружающую среду. В сб.: Региональные и муниципальные проблемы природопользования. Кирово-Чепецк, 1996.
-
91. О с и п о в А.И., С о к о л о в О.А. Роль азота в плодородии почв и питании растений. СПб, 2001.
-
92. П о н о м а р е в а В.В. Теория подзолообразовательного процесса. Л., 1964.
-
93. Щ е р б а к о в А.П., Р у д а й И.Д. Плодородие почв, круговорот и баланс питательных веществ. М., 1983.
-
94. О р л о в Д.С., Б и р ю к о в а О.Н., С у х а н о в а Н.И. Органическое вещество почв Российской Федерации. М., 1996.
-
95. Ч у л к и н а В.А., Т о р о п о в а Е.Ю., Ч у л к и н Ю.И. и др. Агротехнический метод защиты растений (экологически безопасная защита растений). М., 2000.
-
96. Ж у ч е н к о А.А. Экологическая генетика культурных растений. Самара, 2003.
-
97. Н е с т е р о в А.Н. Проблема корневой гнили зерновых колосовых культур в интенсивном земледелии России и Северного Казахстана. Тез. докл. Всер. съезда по защите растений «Защита растений в условиях реформирования агропромышленного комплекса: экономика, эффективность, экологичность». СПб, 1995: 132-133.
-
98. Х а ц к е в и ч Л.К., Н е с т е р о в А.Н. Патогенный комплекс возбудителей корневой гнили яровой пшеницы на южном Урале. Микол. и фитопатол., 1994, 28, 6: 16-19.
-
99. Б у г а С.Ф. Фитопатологическое состояние посевов зерновых культур и его особенности в современных условиях ведения сельского хозяйства. В сб.: Эколого-экономические основы усовершенствования интегрированной системы защиты растений от вредителей, болезней и сорняков. Минск, 1996, 1: 5-6.
-
100. С а н и н С.С., Н а з а р о в а Л.Н., С о к о л о в а Е.А. и др. Здоровье зернового поля. Защита и карантин растений, 1999, 2: 28-31.
-
101. Ж а л и е в а Л.Д., К и р и е н к о в а А.Е. Гнили озимой пшеницы в Краснодарском крае. Защита и карантин растений, 2001, 9: 14.
-
102. К у п р е в и ч В.Ф. К физиологии больного растения: Физиологические данные о вредоносности некоторых грибных и вирусных болезней культивируемых растений. Л., 1934.
-
103. К о р ш у н о в а А.Ф., Ч у м а к о в а А.Е., Щ е к о ч и х и н а Р.И. Защита пшеницы от корневых гнилей. Л., 1976.
-
104. В е т р о в Ю.Ф. Этиология корневой гнили пшеницы в Иркутской области и обоснование методов борьбы с заболеванием. Автореф. канд. дис. Иркутск, 1970.
-
105. Б и л а й В.И. Фузарии. Киев, 1977.
-
106. В а в и л о в Н.И. Теоретические основы селекции растений. М.-Л., 1935.
-
107. Г о н ч а р е н к о А.А., Ш а д у р о С.И. Иммунологическая и селекционная оценка озимой ржи на провокационных и инфекционных фонах. Науч.-техн. бюл. ВИР, 1990, 201: 6-10.
-
108. С ы с у е в В.А. Основные направления и приоритеты научных исследований в Северо-Восточном регионе РФ. Мат. Междунар. науч.-практ. конф. «Озимая рожь: селекция, семеноводство, технологии и переработка». Киров, 2003: 204.
-
109. П о л о в и н к о Г.П. Токсичность фитопатогенных грибов рода Fusarium Lk. Et Fr. Деп. ВНИИТЭИ, 4051-81. М., 1981.
-
110. P a c k a D. Cytogenetic changes in plant cells as influenced by mycotoxins. Mycotoxin Res., 1991, 7A: 150-155.
-
111. M e n k e - M i l c z a r e k I., Z i m n y J. Phytotoxity of deoxynivalenol to wjeat calli. Mycotoxin Res., 1991, 7A: 146-149.
-
112. Л е о н о в А.Н., М а л и н о в с к а я Л.С., С о б о л е в а Н.А. и др. Токсиногенность изолятов Fusarium graminearum Schw. из зерна фузариозной пшеницы в Краснодарском крае. Докл. ВАСХНИЛ, 1990, 3, 11: 40-45.
-
113. Б е н к е н А.А., Х р и с т о в с к а я В.Н. Лабораторная оценка болезнеустойчивости растений и паразитических свойств возбудителей обыкновенной корневой гнили злаков. Тр. ВИЗР, 1977, 56: 913.
-
114. G r e e n h a l g h R., N e i s h G.A., M i l l e r J.D. Mycotoxin formation by Canadian isolates of Fusarium grameniarum on solid culture. Appl. Env. Microbiol., 1983, 46(3): 625-629.
-
115. M i l l e r J.D., B l a c k w e l l B.A. Biosynhesis of 3-acetyldeoxynivalenol and other metabolites by Fusarium culmorum HLX 1503 in a stirred jar fermentor. Can. J. Botany, 1986, 64(1): 1-5.
-
116. Ж у ч е н к о А.А. Стратегия адаптивной интенсификации сельского хозяйства (концепция). Пущино, 1994.
Soil Sci. Soc. Amer., Madison, 1971, 1: 105-151.
and a related species to aluminum in nutrient solution and soil. Austr. J. Agric. Res., 1986, 37, 2: 383-395.
Soc. Hortic. Sci., 1970, 95, 1: 107-110.
New Phytol., 1974, 73, 1: 122-123.
Всероссийский НИИ сельского хозяйства Северо-Востока им.
