Эдификаторная роль Filipendula ulmaria L. на лугах в дельте р. Северной Двины в пределах аллювиальной литогенной матрицы

Автор: Паринова Татьяна Александровна, Волков Алексей Геннадьевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
В статье рассматриваются эколого-флористические особенности лугов с доминированием Filipendula ulmaria, формирующихся на аллювиальных почвах в дельте р. Северной Двины. Представлены результаты, полученные на основе анализа геоботанических описаний и данных полевого опыта. Выделены ведущие экологические факторы, в которых формируются таволговые луга. Установлено распределение видов по фитоценотической значимости, представлены данные о низком видовом богатстве и видовой насыщенности, данные о количественных и качественных показателях продуктивности. Подчёркнута роль таволги, как эдификатора.
Пойменные луга, эдификатор
Короткий адрес: https://sciup.org/146279506
IDR: 146279506 | УДК: 574.4
Defining role of Filipendula ulmaria on meadows in the delta of the Northern Dvina river within the alluvial lithogenic matrix
Ecological and floristic peculiarities of meadows with Filipendula ulmaria dominant is introduced (formed on alluvial soils in the Delta of the Northern Dvina river). Results based on the analysis of geobotanical descriptions and data field experience. Leading environmental factors that affect these meadows was selected. Set the distribution of types in accordance with the phytocoenotic importance. Lowest species richness, number of species per square meter, data about the quantitative and qualitative indicators of productivity are presented. Role of meadowsweet as edificator is highlighted.
Текст научной статьи Эдификаторная роль Filipendula ulmaria L. на лугах в дельте р. Северной Двины в пределах аллювиальной литогенной матрицы
Введение. Filipendula ulmaria (L.) Maxim, (семейство Rosaceae) крупное вегетативно-подвижное поликарпическое короткокорневищное полурозеточное травянистое растение высотой до 2 м, с ползучим симподиальным корневищем, с озимыми моноциклическими надземными побегами, гемикриптофит (Шалыт, I960; Рожанская, 1984; Потапова, 2009; Комаревцева, Черёмышкина, 2016). Ареал вида простирается далеко на восток Европейской части России, и заходит далеко на север по всему Мурманскому берегу. На юге доходит до побережья Черного и Азовского морей на востоке до Енисея, проходит через Алтайский и Красноярский края и уходит в Забайкалье. В Приморье и Северной Америке встречается как заносное (Юзепчук, 1941; Ворошилов, 1982).
F. ulmaria типичный вид крупнотравья, светолюбивый мезотрофный гигромезофит, относится к растениям летно-осенней вегетации, один из видов, диагностирующих среднепойменные и собственно долгопоемные условия (Сапегин, 1985). Произрастает на сырых и заболоченных лугах, в лиственных и смешанных лесах с проточным увлажнением на глинистых и суглинистых иловатых почвах, бедных доступным калием, богатых доступным фосфором. Нередко таволгой зарастают сырые участки свежих вырубок. Подобно другим вегетативно-подвижным растениям, вид часто является доминантом, поэтому имеет большое значение для формирования растительного покрова (Котелина, 1967; Полуянова, 1981; Астрологова, Феклистов, 2002), но данные о его средообразующей роли весьма малочисленны и отрывочны (Ипатов, Кирикова и др., 2007). Известна причина отсутствия таволги на кислых, бедных и слабо дренированных торфяных почвах вследствие угнетения вегетативного роста, возобновления и отсутствия семенного возобновления из-за мощного сфагнового покрова (Рожанская, 1984).
Это растение в благоприятных для него условиях быстро захватывает территорию, вытесняя прочие виды, и прочно удерживает её за собой в течение длительного времени, существенно трансформируя надземную и подземную среду, препятствуя возобновлению древесных пород (Рожанская, 1984). Размножается семенами, переносимыми ветром или водой; одно растение даёт до 500 семян (Губанов, Киселёва, 2003).
На территории Северо-Запада Европейской России широко распространены луга, в которых доминирует F. ulmaria. Их описание в литературе берёт начало из работ А.П. Шенникова (1938). Эти сообщества имеют как первичное, так и вторичное происхождение, относятся к разнотравно-крупнотравным (Введение в ..., 1964). Таволга встречается как в более обводнённых местообитаниях в составе топяно-хвощевых, осоковых, ситниковых, канареевниковых сообществ, так и лисохвостных, болотномятликовых и щучковых лугах (Котелина, 1967; Хантимер, 1959). Таволга вязолистная доминирует, как правило, при условии ненарушенности сообщества и в определённых, довольно узких экологических условиях (Ипатов, Кирикова и др., 2007).
Таволга имеет большое практическое значение, в первую очередь как лекарственное растение очень широкого спектра действия (Скляров, 1990; Стрижев, 1995; Авдеева, Краснов, 2010). Пригодна для дубления и окраски кож. Ценный медонос и перганос (Губанов, Киселёва, 2003). В Швейцарии специально натирают травой лабазника ульи внутри для привлечения пчёл (Алеутский, Кашин, 1998). Летом на таволжниках пасутся лоси (Срижев, 1995). Плоды и листья лабазника вязолистного входят в рацион многих видов птиц «рябчиков, глухарей, тетеревов, уток и др. ( Кучеров и др., 1976).
Методика. Целью работы является выявление эдификаторной роли F. ulmaria на пойменных лугах в дельте Северной Двины. Объект исследования: пойменные луга с проективным покрытием F. ulmaria не менее 60 %. Предмет исследования: эколого-флористические особенности луговых сообществ с эдификатором F. ulmaria.
Проведённые исследования основаны на полевых работах в Приморском районе Архангельской области, в островной части дельты реки Северной Двины на пяти пойменных лугах. Анализировали данные, полученные на 16 геоботанических пробных площадях по 100 м2, которые закладывали на островных лугах на наиболее характерных участках луга в пределах контура растительности и 10 опытных площадках по 1 м2. На стометровых площадях осуществляли полное геоботаническое описание по стандартной методике (Лайдинен, Ларионова и др., 2001).
Рассчитывали индекс видового сходства сообществ П. Жаккара, фитоценотический индекс В.М. Понятовской - И.В. Сырокомской (Баканов, 1987), определяли тип стратегии для видов по двумерной системе Л.Г. Раменского-Дж. Грайма (Раменский, 1935; Grime, 1979). Оценка условий среды по экошкалам проведена при помощи пакета Ecoscale (Грохлина, Ханина, 2015). Для ординации использовали алгоритм неметрического многомерного шкалирования (Kruskal, Wish, 1978), который использует градиентные методы оптимизации эвристических функционалов качества. Его главным преимуществом является то, что он не требует от исходных данных никаких априорных предположений (Шитиков, Розенберг, 2013). В качестве меры различий выбрано Эвклидово расстояние. Для расчетов использовали R и пакет vegan (Oksanen, 2017). Определяли продуктивность травостоев. В пределах каждой стометровой площади определяли валовую продуктивности укосным методом на площадках 0,25 м2, которые закладывали методом случайного отбора в десятикратной повторности (Паринова, Волков, 2017). Для оценки кормовой ценности применяли 10-бальную шкалу Э. Клаапа (1961).
На десяти метровых площадках был заложен двухлетний полевой опытов с целью определения фитоценотической роли таволги вязолистной на островных лугах. В опыте три варианта:
-
- Вариант опыта «В1»: десять площадок размером 1 м2 на лугу с доминированием таволги вязолистной.
-
- Вариант опыта «В2»: на этих же десяти площадках размером 1 м2 таволга аккуратно скошена.
-
- Вариант опыта «ВЗ»: на этих же десяти площадках размером 1 м2 таволга аккуратно выкопана и в следующий полевой сезон площадки обследованы вновь.
На всех площадках проводился учёт в момент максимальной фазы вегетации F. ulmaria по следующим параметрам: полный флористический состав; проективное покрытие каждого вида; фенофаза всех видов; жизненность всех видов; возрастные состояния всех видов; освещённость; температура почвы и воздуха. Освещённость и температура в десятикратной повторности на каждой площадке. Для сравнительного анализа заявленных параметров, были высчитаны их значения параметров с ошибкой (х±тх).
Результаты и обсуждение. На лугах в пойме Северной Двины F. ulmaria редко встречается как одиночное растение. Чаще всего она имеет групповое или контагиозное либо равномерное (отрицательно контагиозной) размещение в сообществах (Уйтеккер, 1980). Исследования проводили в сообществах только со сплошным размещением таволги.
В соответствии с «Кодексом фитосоциологической номенклатуры» (Weber, Moravec and other, 2000) рассматриваемые луга со сплошным размещением F. ulmaria относятся к АССОЦИАЦИИ Filipenduletum ulmariae (ВАРИАНТ Alopecurus pratensis и ВАРИАНТ Elitrtgia repens) —> ПОДСОЮЗУ Filipendulenion (Lorn, in Oberd. et al 1967) Bal.-Tul. 1978 ^ СОЮЗУ CalthonR. Tx. 1937 em Bui. - Tul. 1978 ^ ПОРЯДОКУ Molietalia W. Koch 1926 -^ КЛАСС Molino-Arrhenathereta R. Tx. 1937 em R. Tx 1970.
Эти луга на островных лугах в дельте Северной Двины формируются на почвах аллювиального типа, по международной классификации - Fluvisol (WRB, 2014), но почвы отличаются по степени богатства-засоления и увлажнению. Показатели богатства колеблется от 9 (почвы не богатые — мезотрофные со слабокислой реакцией среды) до 13 (почвы довольно богатые со слабокислой и нейтральной реакцией среды). Небогатые почвы формируются на участках таволговых лугов близких к берегу островов и в рельефных депрессиях их центральной части. Луга с таволгой в центре островов отличаются более богатыми почвами. Различия в богатстве почв на изученных участках связаны, в первую очередь, с разной интенсивностью аллювиально-пойменных процессов. Условия увлажнения на исследуемых площадках колеблются в достаточно больших пределах от 66,5 (влажнолуговое, почвы обычно без признаков оглеения или со слабым оглеением) до 83,5 (сыролуговое, почвы с явными признаками оглеения и признаками начальных стадий оторфовывания) баллов.
Различия в увлажнении исследованных таволговых лугов объясняются разностью высоты берега над меженью воды (чем ниже -тем интенсивнее влияние полых вод и тем сырее), различиях в залегании грунтовых вод, удалённостью от центрального русла и проток, микропонижениями и микроповышениями рельефа.
Пастбищная дигрессия на пробных площадках выражена слабо (от полного отсутствия выпаса до сенокосной стадии). Отсутствие или слабое влияние выпаса на изученных таволговых лугах объясняется тем, что их основной доминант таволга - практически не поедаемый вид высокого, крупнолистного, грубостебельного разнотравья. Этот вид не поедается скотом как в свежем виде, так и в сене, поэтому на таких участках травостой обычно остаётся нетронутым человеком и животными. Такие луга не используются ни как пастбища, ни как сенокосы (Ржанская 19 84; Василевич В.И., Беляев Е.А., 2005).
Изученные таволговые луга невозможно строго отнести ни к одной из стадий развития луга по соотношению жизненных форм (Дербин, 1948). Так как, с одной стороны, в травостое преобладает короткокорневищная F. ulmaria, с другой стороны, в примесь к ней идут виды, способные выдерживать её мощное эдификаторное воздействие - это длиннокорневищные Elytrigia repens (L.) Nevski и Vicia cracca L., V. septum L., иногда рыхлокустовой Alopecurus pratensis L. Кроме того, некоторые участки лугов разной степени закочкаренности, это и кочки плотнодерновинного злака Deschampsia cespitosa (L.) Beauv. и плотнодерновинной осоки Carex cespitosa L. Поэтому обозначим стадию развития таволговых лугов по соотношению жизненных форм как смешанную.
Общее видовое богатство исследуемых фитоценозов представлено 52 видами, что составляет 28 % от общего флористического списка лугов островной поймы в дельте С. Двины, на которых отмечено 186 видов сосудистых растений (Паринова, Наквасина, 2013). Из чего можно сделать вывод, что флористический состав сравнительно не богат. Видовая насыщенность на 100 м2 от 7 до 23 видов, в среднем 15 видов, не соответствует средним показателям для пойменных лугов таёжной зоны, для которых она составляет в среднем около 30 видов (Луговодство в ..., 1955).
Низкие показатели видового богатства и видовой насыщенности объясняются мощными эдификаторными свойствами таволги. Как указывает Ипатов, Кирикова и др. (2007), в результате перехвата светового потока и развития плотной дернины он негативно влияет на развитие сопутствующих видов, для которых экотоп в месте произрастания таволги благоприятен. В этом процессе принимают участие и содоминанты, находящиеся под пологом таволги, которые в свою очередь испытывают её отрицательное влияние. Таволговый опад, как поставщик органики в почву, благоприятен для всех видов, формирующих сообщество, но влияние режима освещённости и плотная дернина нивелируют его влияние. Такое соседство выдерживают не многие виды.
Не очень высокие индексы сходства видового состава таволговых сообществ в пойме по коэффициенту Жаккара (от 5 до 94%, в среднем 27%) могут быть объяснены тем, что таволговые луга занимают относительно не большие площади, со всех сторон окружены другими луговыми фитоценозами с различным видовым составом, что не может не отражаться на видовом состоянии. Также, относительно широкий диапазон экологических условий таволговых лугов даёт вариации сопутствующих видов при сохранении доминирующих позиций F. ulmaria. Да и в целом, пойменные луга в низовьях р. Северной Двины отличаются высоким видовым разнообразием и пестротой.
Среди разнотравья по индексу фитоценотической значимости, бесспорным лидером является таволга вязолистная (индекс равен 114), в соответствии с рисунком 1.
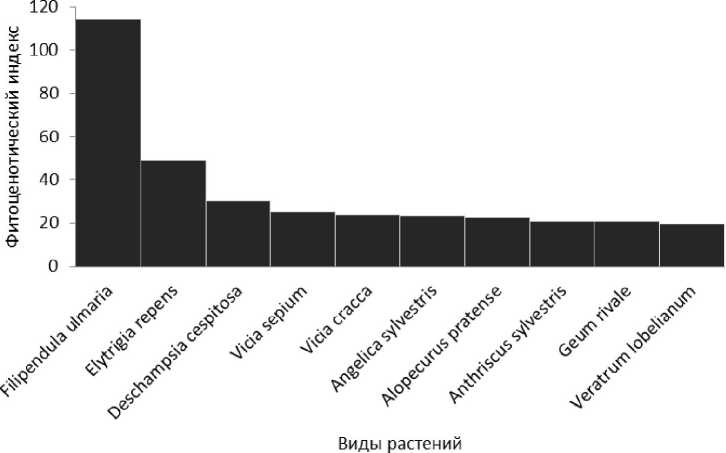
Рис. 1 . Распределение видов в травостое таволговых лугах с наибольшими значениями индексов фитоценотической значимости
Диаграмма индексов наибольшей фитоценотической значимости показывает, что наибольшее значение индекса после F. ulmaria имеет Е. repens. Это длиннокорневищный высокорослый вид, с преимущественно вегетативным размножением. Растение широкой экологии: имеет гигромезофильные, мезофильные, галомезофильные и ксеромезофильные экотипы; поймовыносливо. Благодаря своей вегетативной и экологической пластичности пырей один из немногих злаков способен выдерживать такого мощного эдификатора как F. ulmaria. Высокий индекс имеет D. cespitosa - плотнодерновинное растение с высокой семенная продуктивность (от 1600 зерновок на 1
м2), способное долго удерживать своих позиций в травостое, формируя мощные фитогенные поля, устойчиво к затенению. По индексам фитоценотической значимости также лидируют V. сгасса и V. septum. Это длиннокорневищные мезофиты с широкой экологической амплитудой, теневыносливые растения. На корнях горошков поселяются клубеньковые бактерии, которые обеспечивают их дополнительным питанием, что очень важно в условиях сожительства с таволгой, которая перехватывает питательные вещества из почвы.
Анализ видового состава по двумерной системе Л.Г. Раменского - Дж. Грайма (Grime, 1979) первичных и вторичных типов стратегий (рис. 2) показал преобладание видов со вторичным типом стратегии CSR. Это луговые растения, которые сочетают в своём поведении черты виолетности, патиентности и эксплерентности. Сама F. ulmaria по функциональному типу является фитоценотическим патиентом с выраженными виолентными признаками, а содоминант Е. repens - типичный эксплерент.
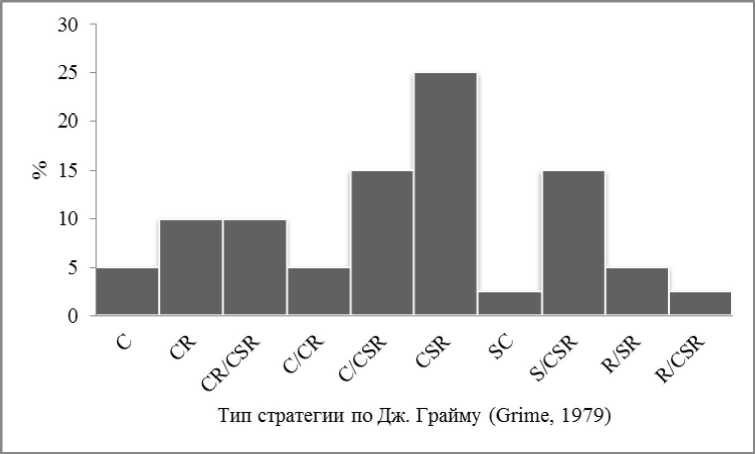
Рис. 2. Распределение видов в травостое таволговых лугов по функциональным типам в системе Л.Г. Раменского-Дж. Грайма
Экологическое пространство, в котором формируются таволговые луга дельты Северной Двины, представлено на рисунке 3, как результат неметрического многомерного шкалирования таволговых лугов на основе меры сходства в виде Евклидова расстояния, которая выбрана с учётом отражения покрытия видов и сильным акцентом на роли доминантов. Ядро описаний представляют площадки, на которых наибольший фитоценотический индекс имеют выделенные ранее F. ulmaria, Е. repens, У. septum. Рисунок отражает данные, как флористической неоднородности изучаемых лугов, так и диапазон экологических условий, в которых они формируются. Векторы факторов по шкалам Л.Г. Раменского отражают разброс единичных площадок, в частности, по пастбищной дигрессии. В целом наибольшей значимостью в распределении всех площадок обладают векторы, которые отражают «богатство почв», «кислотность почв», их «плодородие», в меньшей степени, чем «увлажнение почв» (таблица 1). Для получения более полной и детальной картины необходимо продолжить сбор соответствующих описаний.
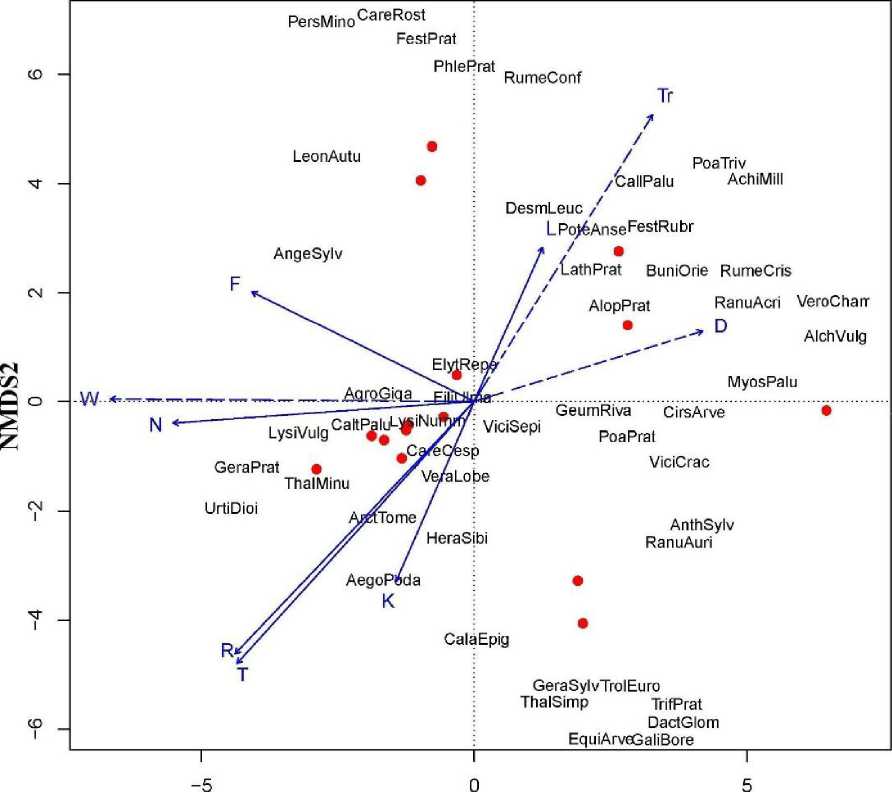
NMDS1
Рис. 3 . Результат неметрического многомерного шкалирования таволговых лугов: Шкалы Г. Элленберга: F - увлажнение почв; N - богатство почв азотом; R - кислотность почв; L - освещенность/затенение; Т - термоклиматическая; К - континентальность климата. Шкалы Л.Г. Раменского: W - шкала увлажнения; Тг - шкала активного богатства и засоления почвы; D - шкала пастбищной дигрессии.
Таблица 1
Результаты расчета экологических векторов
|
Обозначение на графике |
Экологическая шкала |
Показатель |
Коэффициенты корреляции с осями |
R2 |
Р |
|
|
NMDS1 |
NMDS2 |
|||||
|
W |
Раменского |
Влажность |
-0,999 |
0,006 |
0,53 |
0,006 |
|
Т |
Элленберга |
Термоклиматическая |
-0,672 |
-0,740 |
0,50 |
0,012 |
|
R |
Элленберга |
Кислотность почв |
-0,688 |
-0,724 |
0,48 |
0,008 |
|
Тг |
Раменского |
Богатство почв |
0,528 |
0,848 |
0,46 |
0,021 |
|
N |
Элленберга |
Богатство почв азотом |
-0,997 |
-0,071 |
0,36 |
0,052 |
|
F |
Элленберга |
Увлажнение почв |
-0,896 |
0,443 |
0,25 |
0,160 |
|
D |
Раменского |
Пасбищная дегрессия |
0,955 |
0,294 |
0,23 |
0,175 |
|
К |
Элленберга |
Континентальность климата |
-0,399 |
-0,916 |
0,15 |
0,334 |
|
L |
Элленберга |
Освещенность/затенение |
0,405 |
0,914 |
0,11 |
0,471 |
Примечание: Жирным шрифтом выделены векторы, значимые, при р<0,05.
Таблица 2
Экологические параметры на пробных площадках полевого опыта
|
Параметры (ср. значения с ошибкой Х±Щх) |
Вариант опыта В1 контроль |
Вариант опыта В2 таволга скошена |
Вариант опыта Вз таволга выкопана в пр. году |
|
Освещённость, % |
21,66±2,085 |
45,27±0,026 |
36,14±0,058 |
|
Температура воздуха, °C |
15,04±0,026 |
15,00±0,000 |
15,5 НО,024 |
|
Температура почвы, °C |
14,00±0,084 |
14,2 НО,064 |
13,20±0,905 |
|
Общее проективное покрытие, % |
98,00±1,106 |
38,00±5,540 |
43,20±3,560 |
|
Высота травостоя, см |
150,6Н4,580 |
52,69±6,880 |
6,60±4,200 |
|
Видовая насыщенность (на 1 м2) |
7, НО,64 |
7, НО,64 |
10,НО,5 |
|
Общий флористический список для 10 площадок |
13 |
13 |
26 |
|
Преобладающие возрастные состояния |
средневозрастные генеративные |
средневозрастные генеративные |
ДО генеративные |
Среднее арифметическое урожая сена изученных таволговых лугов составляет 47,6 ц/га, что соответствует средним значениям продуктивности для неиспользуемых сенокосных пойменных лугов Архангельской области (Паринова, Наквасина и др., 2013). Основную долю в формировании высоких показателей продуктивности вносит разнотравье, в первую очередь таволга - крупногабитусный вид широколистного и грубостебельного разнотравья. Доля видов остальных агроботанических групп незначительна. Так как при высоких количественных показателях продуктивности качественный состав сена может сильно различаться, была проведена оценка кормовой ценности травостоев. Средний кормовой оценочный балл для таволговых лугов 3,45 (по 10-балльной шкале от -1 до 8). В целом, таволговые луга характеризуются высокими количественными показателями продуктивности, при низких значениях кормовой ценности.
В таблице 2 отражены параметры, полученные в ходе полевого опыта на таволговом лугу.
Полевой опыт показал следующие результаты, дополняющие и подтверждающие другие исследования:
-
- Фактор температуры почвы и воздуха в опыте не проявил себя. Требуются дополнительные исследования. Температура в большей степени определяется общими погодными условиями.
-
- При удалении F. ulmaria из травостоя (скашивании, выкапывании) резко возрастает освещённость, что благоприятствует развитию сопутствующих видов, это проявляется в увеличении их проективного покрытия на фоне общего снижения покрытия травостоя).
-
- Полное аккуратное удаление F. ulmaria из травостоя приводит к увеличению общего числа видов и видовой насыщенности. Не только увеличивается покрытие ранее обитавших с ней видов, но и происходит внедрение в травостой новых видов. За счёт почвенного банка семян появляются: Роа pratensis L., Cirsium arvense (L.) Scop., Stellaria longifolia Muehl. ex Willd., Atriplex patula L., Polygonum aviculare L., Persicaria maculata (Rafin.) A. & D. Love, Myosotis palustris (L.) L., Lepidotheca suaveolens (Pursh) Nutt. За счёт вегетативного размножения из окружающего площадки травостоя: Amoria repens (L.) С. Presl, Ranunculus acris L.
-
- На площадках происходит смена доминантных видов и, прежде всего, усиливают свои позиции высокорослые грубостебельные виды Cirsium arvense arvense (L.) Scop, (абсолютный трудноискоренимый сорняк) и Heracleum sibiricum L. (факультативный сорняк). Крайняя степень засилья травостоя этими видами может выражаться в формировании одноярусных сообществ без мохового покрова, с деградирующей дерниной и практически полным отсутствием злаков и бобовых (Паринова, Наквасина и др., 2013).
-
- Травостой без F. ulmaria становится более «молодым», среди особей
всех видов травостоя догенеративные растения преобладают над генеративными.
Заключение. F. ulmaria поселяется на пойменных лугах чаще в прирусловой части поймы или в других частях, но вблизи проток или в понижениях микрорельефа (на аллювиальных почвах с достаточным или избыточным увлажнением). Закрепившись в сообществе, таволга начинает проявлять мощное эдификаторное влияние на все соседствующие виды. Только виды, обладающие определёнными экологическими и морфологическими свойствами способны сосуществовать с ней. Таволговые пойменные луга в дельте Северной Двины не однородны по своим эколого-флористическим характеристикам: изученные пробные площади дистанцируются в пространстве экологических факторов и по видовому сходству. Видовое богатство представлено 52 видами. Видовая насыщенность 23 вида. Флористический состав таволговых лугов сравнительно не богат. Данные показатели значительно ниже известных средних данных для пойменных лугов в целом. В травостое преобладают виды со смешанным типом стратегий по Л.Г. Раменскому-Дж. Грайму. С выпадением из сообщества таволги вязолистной для других растений становятся доступны в большем количестве свет и элементы питания в травостое, увеличивается видовой состав и видовая насыщенность.
Отдельно необходимо подчеркнуть, что без мощного действия таволги вязолистной, на пойменных лугах могут произрастать многие виды. В первую очередь быстро начинают заселять территорию однолетние и многолетние «луговые сорняки», распространение которых ранее сдерживала таволга. Поэтому умеренное распространение таволги в виде полос вдоль используемых пойменных лугов может выступать естественным сдерживающим барьером для распространения «сорных» видов. Однако полное отсутствие сенокошения на пойменных лугах в совокупности с изношенной системой осушения приводит к массовому распространению таволги по всем лугам. Данный процесс наблюдается в дельте последние десятилетия, особенно на лугах левобережной поймы под г. Архангельском. Это негативно сказывается не только на хозяйственных характеристиках травостоев, но и на биологическом разнообразии (как флористическом, так и фаунистическом) пойменных лугов.
DEFINING ROLE OF FILIPENDULA ULMARIA ON MEADOWS IN
THE DELTA OF THE NORTHERN DVINA RIVER WITHIN THE ALLUVIAL LITHOGENIC MATRIX
T.A. Parinova, A.G. Volkov
Lomonosov Northern (Arctic) Federal University, Archangelsk
Ecological and floristic peculiarities of meadows with Filipendula ulmaria dominant is introduced (formed on alluvial soils in the Delta of the Northern Dvina river). Results based on the analysis of geobotanical descriptions and data field experience. Leading environmental factors that affect these meadows was selected. Set the distribution of types in accordance with the phytocoenotic importance. Lowest species richness, number of species per square meter, data about the quantitative and qualitative indicators of productivity are presented. Role of meadowsweet as edificator is highlighted. Keywords: Filipendula ulmaria, meadows, edificator.
Об авторах:
Паринова Т.А. Эдификаторная роль Filipendula ulmaria L. на лугах в дельте р. Северной Двины в пределах аллювиальной литогенной матрицы / Т.А. Паринова, А.Г. Волков // Вести. ТвГУ. Сер. Биология и экология. 2018. № 2. С. 181-194.
Список литературы Эдификаторная роль Filipendula ulmaria L. на лугах в дельте р. Северной Двины в пределах аллювиальной литогенной матрицы
- Авдеева Е.Ю., Краснов Е.А. 2010. Биологическая активность Filipendula ulmaria (Rosaceae)//Растительные ресурсы. Т. 46. №3. С. 123-130.
- Алеутский Н.Н, Кашин В.И. 1998. Северная кладовая целебных трав.: Архангельск «Правда Севера». 414 с.
- Астрологова Л.Е, Феклистов П.А. 2002. Лекарственные растения Севера. Архангельск: Изд-во АГТУ. 156 с.
- Баканов А.И. 1987. Количественная оценка доминирования в экологических сообществах. Борок: ВИНИТИ. 63 с.
- Василевич В.И., Беляев Е.А. 2005. Таволговые луга Северо-Запада Европейской России//Бот. Журнал. Т. 90. № 12. С. 1801-1813.
- Введение в геоботанику. 1964/отв. ред. А.П. Шенников. Л.: изд-во Ленинградского ун-та. 446 с.
- Ворошилов, В.Н. 1982. Определитель растений советского Дальнего Востока. М.: Наука. 672с.
- Грохлина Т.И., Ханина Л.Г. 2015. О компьютерной обработке геоботанических описаний по экологическим шкалам//Математическое моделирование в экологии: материалы Четвертой Национальной научной конференции с международным участием, 18-22 мая 2015 г. Пущино. ИФХиБПП РАН. С. 63-64.
- Губанов И.А., Киселёва К.В., Новиков В.С., Тихомиров В.Н. 2003. Иллюстрированный определитель растений Средней России. Т. 2: Покрытосеменные (двудольные: раздельнолепестные). М.: Т-во научных изданий КМК, Ин-т технологических исследований. 665 с.
- Дербин JI.A. 1948. Стадии развития луга//Улучшение и использование лугов и пастбищ на Севере. Архангельск: ОГИЗ Архангельское издательство. С. 35-45, 39-45.
- Ипатов, В.С., Кирикова, Л.А., Майор, И.А. 2007. Влияние Filipendula ulmaria (Rosaceae) насопутствующие виды в луговыхсообществах//Бот. журн. Т. 92. № 7. С. 1046-1053.
- Клаап Э. 1961. Сенокосы и пастбища. М.: Изд-во сельхоз. лит-ры, журналов и плакатов. 613 с.
- Комаревцева Е.К., Черёмушкина В.А. 2016. Развитие Filipendula ulmaria (Rosaceae) и онтогенетическая структура его ценопопуляций на юге Западной Сибири//Растительный мир Азиатской России. № 1 (21). С. 35-41.
- Котелина Н.С. 1967. Динамика луговой растительности долины реки Вычегды. Л.: Наука. 85 с.
- Кучеров Е.В., Байков К.К., Гуфранова И.Б. 1976. Полезные растения Южного Урала. М: Наука. 264 с.
- Лайдинен, Г.Ф., Ларионова Н.П., Лантратова А.С. 2001. Геоботаническое изучение луговой растительности//Методы полевых и лабораторных исследований и растительных сообществ. Петрозаводск: ПетрГУ. 296 с.
- Луговодство в поймах рек. 1955/Под ред. С.П. Смелова, Н.С. Конюшкова. М.: Госуд. изд-во с/х лит-ры. 256 с.
- Паринова Т.А., Волков А.Г. 2017. Методы изучения луговых экосистем. Учебное пособие. Архангельск: Кира. 142 с.
- Полуянова В.И. 1981. Исследования морфологической структуры ценопопуляций вегетативно-подвижных растений: Автореф. дис. … канд. биол. наук. Свердловск. 19 с.
- Потапова М.С. 2009. Особенности онтоморфогенеза Filipendula ulmaria (Rosaceae) (L.) Maxim. в различных экологических условиях//Вестник Российского университета дружбы народов. Серия: Экология и безопасность жизнедеятельности. № 3. С. 37-45.
- Рожанская О.А. 1984. Экологические и биологические свойства лабазника вязолистного Filipendula ulmaria (L.) Maxim. Диссертация на соискание учёной степени кандидата биологических наук. М. 2015 с.
- Сапегин Л.М. 1985. Пойменные луга юго-востока БССР, их рациональное использование, улучшение и охрана. Минск: издательство "Университетское". 1985. 100 с.
- Скляров Э.Л. 1990. Съедобные и лекарственные дикие растения Русского Севера. Архангельск. 272 с.
- Стрижев А.Н. 1995. Русское разнотравье. М.: Дрофа. 575 с.
- Уиттекер Р. 1980. Сообщества и экосистемы/ред. Т.А. Работнова. М.: Прогресс. 315 с.
- Хантимер И.С. 1959. Материалы к изучению лугов поймы р. Печоры//Луга Республики Коми. М.-Л. С. 175-265.
- Шалыт М.С. 1960. Методика изучения морфологии и экологии подземной части отдельных растений и растительных сообществ//Полевая геоботаника. Т.2. М. Л. С. 369-447.
- Шенников А.П. 1938. Луговая растительность СССР//Растительность СССР. Т.1. М.-Л. С. 429-638.
- Шитиков В.К., Розенберг Г.С. 2013. Рандомизация и бутстреп: статистический анализ в биологии и экологии с использованием R. Тольятти: Кассандра. 314 с.
- Юзепчук C.B. 1941. Сем. Rosaceae//Флора СССР. Т. 10. М. Л. 284 с.
- Grime J.P. 1979. Plant Strategies and Vegetation Processes. John Wiley & Sons, Ltd., Chichester. New York-Brisbane-Toronto. 222 pp.
- Oksanen J., Blanchet F., Friendly M. 2017. Vegan: Community Ecology Package. R package version 2.4-4.
- Kruskal J.B., Wish M. 1978. Multidimensional scaling. Sage University Paper series on Quantitative Applications in the Social Sciences, number 07-011. Sage Publications, Newbury Park, CA. 93 p.
- Weber H.E., Moravec J., Theurillat J.-P. 2000. International Code of Phytosociological Nomenclature 3 rd edition//J.Veg. Sci. V. 11. № 5. P. 739-768.
- World reference base for soil resources 2014 (WRB, 2014). 2015. International soil classification system for naming soils and creating legends for soil maps. Rome. 192 p.
