Эффект агрегаций на половозрастную структуру Xenylla obscura (Collembola, Hypogastruridae) на Витимском плоскогорье (Забайкалье)
 на Витимском плоскогорье (Забайкалье)")
Автор: Гулгенова А.Б., Потапов М.Б.
Журнал: Природа Внутренней Азии @nature-inner-asia
Рубрика: Биология
Статья в выпуске: 4 (33), 2025 года.
Бесплатный доступ
Исследование посвящено изучению сезонной динамики и половозрастной структуры многочисленного вида коллембол Xenylla obscura Imms, 1912 в биотопах юга Витимского плоскогорья (Еравнинская и Кондинская котловины). Работа основана на материале, собранном в полевые сезоны 2008–2009 гг. В результате исследования установлено, что в соотношении полов у X. obscura в большинстве случаев преобладают самки, составляя 60–70 % популяции, что характерно в целом для коллембол. Однако в некоторых сроках взятия проб в разнотравном и ерниковом лиственничниках были зафиксированы высокие доли самцов (75–76 %), что, вероятно, связано с образованием размножающихся агрегаций. На основе анализа присутствия репродуктивных самцов сделан вывод, что основной пик размножения вида приходится на первую половину лета. К концу лета наблюдается массовое появление ювенильных особей, которые к октябрю завершают рост и, вероятно, уходят на зимовку в предгенеративном состоянии. Обнаружение размножающихся самцов в августе свидетельствует о возможности растянутого репродуктивного периода.
Сезонная динамика, криоаридные котловины, агрегированность, ногохвостки, жизненный цикл, соотношение самцов и самок
Короткий адрес: https://sciup.org/148332664
IDR: 148332664 | УДК: 595.7 (571.54) | DOI: 10.18101/2542-0623-2025-4-30-39
Effect of Aggregations on Age-Sex Structure of Xenylla obscura (Collembola, Hypogastruridae) in the Vitim plateau (Transbaikalia)
We have investigated the seasonal dynamics and age-sex structure of the numerous species of springtails Xenylla obscura Imms, 1912 in the biotopes of the southern Vitim Plateau (Eravninskaya and Kondinskaya Basins). The study is based on material collected during the 2008–2009 field seasons. The sex ratio of X. obscura is predominantly female, accounting for 60–70 % of the population, which is typical for springtails in general. High proportions of males (75–76 %), however, were recorded in mixed-herb and dwarf birch larch forests, which are likely associated with the formation of breeding aggregations. Based on an analysis of the presence of reproductive males we have concluded that the main reproductive peak occurs in the first half of summer. By late summer, we observed mass hatching of juveniles, which had completed their growth by October and likely hibernate in a pre-reproductive state. The discovery of breeding males in August suggests the possibility of an extended reproductive period.
Текст научной статьи Эффект агрегаций на половозрастную структуру Xenylla obscura (Collembola, Hypogastruridae) на Витимском плоскогорье (Забайкалье)
Почвенные беспозвоночные, в частности ногохвостки, благодаря высокой численности и разнообразию, участию в процессах деструкции органического вещества являются важными компонентами функционирования наземных экосистем. Исследование сезонной динамики и структуры популяций этих организмов важно для понимания не только биологии отдельных видов, но и устойчивости сообществ в меняющихся условиях, что особенно актуально для криоаридных территорий. Xenylla obscura Imms, 1912 — вид, широко распространенный во многих биотопах юга Витимского плоскогорья, достигает высоких численностей и входит в число доминантов лесных биотопов.
В настоящей работе мы попытались изучить сезонную динамику X. obscura и связать соотношение самцов и самок с наблюдаемым в наших материалах агрегированным распределением данного вида.
Материал и методика
Объект исследования. Xenylla obscura Imms, 1912 описан из северо-восточной Индии, в массовом скоплении, с высоты около 2 100 м над уровнем моря. В настоящий момент этот вид широко отмечен в Азии, но определяется в широком понимании, т. е. «sensu lato», так как первоописание сильно устарело. Форма, изученная нами в Бурятии, соответствует описанию, данному в Определителе коллембол России и сопредельных стран [1994].
Район исследования. Сбор материала проводился в полевые сезоны 2008– 2009 гг. на территории Еравнинской и Кондинской котловин, расположенных в южной части Витимского плоскогорья (Восточная Бурятия). Для этих котловин характерен криоаридный резко континентальный климат, наличие сезонной и многолетней мерзлоты, расчлененный рельеф и значительное разнообразие типов почв [Бадмаев и др., 2006]. Отбор проб проводился бурением в диаметре 5 см на глубину 15–20 см. В ходе полевых исследований 2008 г. в 13 биотопах юга Витимского плоскогорья [Чимитова, Чернова, Потапов, 2010] был проведен двукратный отбор проб (по 10 образцов в июне и августе — сентябре). В 2009 г. работа была продолжена в 6 биотопах, где пробы отбирались четыре раза за сезон (по 15 образцов в июне, июле, августе и сентябре). В Кондинской котловине исследовано три биотопа: разнотравный и ерниковый лиственничники, а также ерник осоковый с пятилистником. В Еравнинской котловине — степи кобрезиево-типчаково-полын-ная, эдельвейсово-кобрезиевая и эдельвейсово-овсянициево-кобрезиевая [Чими-това, 2010]. Выгонку коллембол проводили по стандартной методике до полного высыхания субстрата с помощью воронок Тулльгрена [Гиляров, 1975].
Измерение длины головной капсулы у разных полов и возрастов. В связи с тем, что у коллембол нет выраженного метаморфоза, линьки у них происходят в течение всей жизни (т. е. и после достижения половозрелости), для определения возраста у них измеряют длину тела [Takeda, 1984]. В нашем исследовании мы с помощью рисовального аппарата проводили промеры головных капсул коллембол, потому что посчитали этот параметр менее зависимым от фиксации особи в препарате, чем общая длина тела. В каждом из сроков в биотопах, где обнаружена X. obscura, были промерены и рассмотрены по 50 экземпляров этого вида, если же общее количество было меньше, то измеряли все экземпляры.
Определение половозрастной структуры. У коллембол отмечено, что условно взрослые особи могут быть незрелой (предгенеративной) и зрелой (генеративной) стадии [Takeda, 1984]. Эти стадии отличаются по открытости полового отверстия — оно закрыто у предгенеративной и открыто у генеративной. Согласно Петерсену [Petersen, 1978] предгенеративная стадия не просто сменяется генеративной, а эти стадии чередуются друг с другом. В нашем исследовании были выделены следующие группы: ювенилы, самки, предгенеративные самцы и генеративные самцы. Мелкие особи с неразвитым генитальным полем, т. е. где пол коллемболы было невозможно определить, были отнесены к ювенилам. Самцов и самок различали по форме полового отверстия. Из-за сложности определения половозрело-сти у самок Poduromorpha (куда относится X. obscura ) самок мы не разделяли на предгенеративные и генеративные, а рассматривали как одну группу. Самцов разделили на две группы: предгенеративные (subadult) и генеративные (adult), особи последней группы имеют хорошо выраженный «ductus ejaculatorius»).
Результаты исследования
Размер головной капсулы у разных возрастов и полов. На основе проведенных промеров головных капсул были выделены следующие размерные классы: 1 (0–0,1 мм), 2 (0,1–0,14 мм), 3 (0,15–0,17 мм) (табл. 1). Из всех рассмотренных ювенилов 94 % приходится на первый размерный класс и только 6 % — на второй. Подавляющее большинство взрослых особей (76 %) отнесено ко второму, 18 % — к первому и 6 % — к третьему классу. Значительных отличий между размерами головных капсул у предгенеративных и генеративных самцов не выявлено. Самки по размерам близки к самцам.
Таблица 1
Процентное соотношение размерных классов по длине головных капсул у разных половозрастных групп X. obscura (по всем промеренным экземплярам)
|
Половозрастные группы / размерный класс |
1 (0–0,1 мм) |
2 (0,1–0,14 мм) |
3 (0,15–0,17 мм) |
|
ювенилы (n=278) |
94 |
6 |
0 |
|
cамки предгенеративные и генеративные (n=124) |
23 |
71 |
6 |
|
самцы предгенеративные (n=119) |
15 |
79 |
6 |
|
самцы генеративные (n=23) |
6 |
88 |
6 |
Средние показатели половозрастной структуры. Ювенилы составляли от 2 до 100 % общего числа коллембол в каждом из сроков. Соотношение самцов и самок определяли их сравнением в общей популяции взрослых особей. Самцы составляли от 14 до 81 % взрослого населения (табл. 2). В двух случаях процент самцов был значительно ниже или выше средних значений: в сосняке рододендроновом в августе 2008 г. — 14 %, в лиственничнике разнотравном в июне 2009 г. — 81 % самцов. Такие крайние значения, вероятно, случайны, так как связаны с небольшим количеством экземпляров в выборках: 55 и 18 соответственно. Если не учитывать эти два случая, самцы составляли в среднем 48 % популяции, но в большинстве сроков — 30–40 %. Доля самцов в лиственничнике разнотравном в июне 2008 г. и в лиственничнике ерниковом в сентябре 2009 г. была достоверно выше: 75 % и 76 % соответственно.
Таблица 2
Доля взрослых особей в популяции и доля самцов среди взрослых особей у X. obscura в различных биотопах юга Витимского плоскогорья
|
лиственничник разнотравный |
лиственничник ерниковый |
§ S ° о Й S ° 4 w s S H & к W О |
о к ч Оч н и со О С- о |
эВ со к ^ ч |
о о 4 S “ 1 Я Я Ц Оч 1 (Й о н И О О >> S о Оч ° О ю |
||||||
|
июн. 08 |
июн. 09 |
сент. 09 |
авг. 08 |
июн. 09 |
авг. 09 |
сент. 09 |
июн. 08 |
авг. 08 |
авг. 08 |
июн. 08 |
|
|
общее кол-во, экз. |
4 739 |
55 |
59 |
4 402 |
29 |
871 |
431 |
419 |
86 |
18 |
57 |
|
процент взрослых особей |
96 |
35 |
2 |
14 |
52 |
64 |
98 |
90 |
65 |
39 |
54 |
|
процент самцов |
75 |
81 |
0 |
43 |
40 |
56 |
76 |
27 |
32 |
14 |
39 |
Сезонные изменения половозрастной структуры в различных биотопах
Из рассмотренных в 2008 г. 13 биотопов X. obscura встречается в 11, в 6 из них он входит в число доминантов (при пороге доминирования 10 %): лиственничники ерниковый и разнотравный, сосняк рододендроновый, березово-лиственничный лес с шиповником, ерник осоковый с пятилистником, разнотравно-осоковый луг. При этом в одном и том же биотопе в разные сроки на данный вид приходится от 1,4 до 93 % общего обилия ногохвосток.
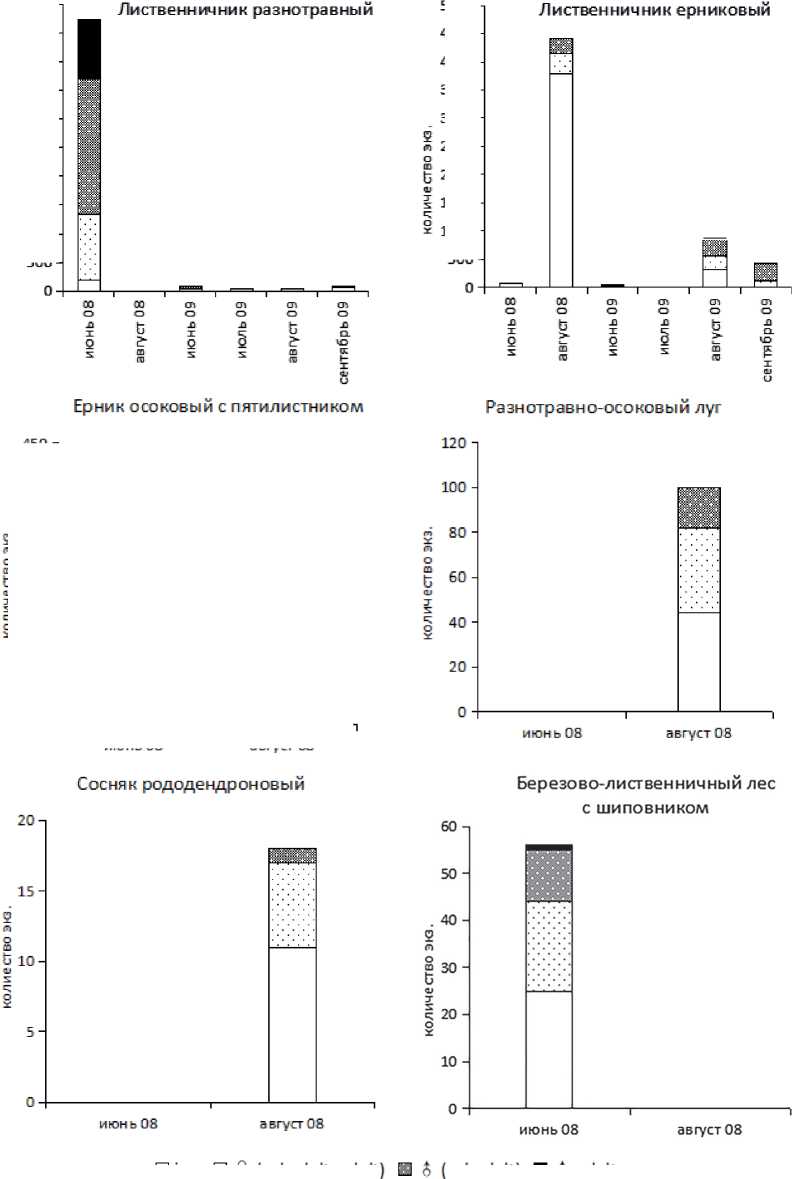
Половозрастная структура данного вида в районе исследования рассмотрена в 6 биотопах. Сезонную динамику рассматривали в основном в лиственничниках, поскольку в части биотопов вид встречается только в одном из сроков (рис. 1).
Точные сроки размножения и возраст зимующих особей данного вида остаются до конца невыясненными, так как четких фенологических закономерностей выявить не удалось. Сезонная динамика численности данного вида отличается как между биотопами, так и в разные годы в одном биотопе. Так, в разнотравном лиственничнике в 2008 г. X. obscura дала вспышку численности в июне, в ернико-вом — в конце августа, в 2009 г. резких изменений численности не обнаружено. При этом в пробах со вспышками численности отмечена высокая агрегированность.
5000 п
5000-1
4500-
4500-
4000-
4000-
3500-
3500-
3000-
3000-
2500-
2500- b
2000-
2000-
1500-
1500-
1000-
1000-
500-
500-
□ juv. □ ¥ (subadult; adult)
(subadult) ■ $ adult
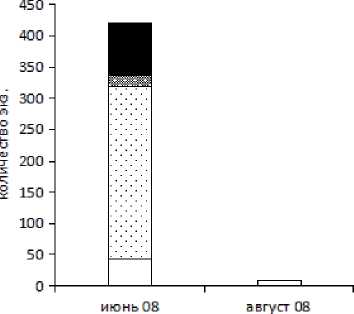
Puc. 1 . Половозрастная структура популяции.^ obscura в различных биотопах (juv. — ювенильные особи; subadult — предгенеративные; adult — генеративные особи)
В ерниковом лиственничнике индекс агрегированности Лексиса (λ) у X. obscura в 2008 г. увеличился с 8,0 в июне до 59,5 в августе, в разнотравном лиственничнике снизился с 52,4 в июне до 1,7 в августе.
В начале лета, в июне 2008 г. в разнотравном лиственничнике популяция X. obscura состоит в основном из взрослых особей с малой долей ювенилов и преобладанием в популяции самцов, свыше 30 % которых являлись генеративными. В ернике осоковом с пятилистником наблюдали схожую картину с большей долей самок, при этом большая часть самцов (80 %) также генеративная. В ерниковом лиственничнике обнаружено небольшое число особей этого вида, все ювенилы. В березово-лиственничном лесу также преобладают ювенилы, самцов 39 %, среди которых есть небольшой процент генеративных.
К концу августа в ерниковом лиственничнике анализ массовых скоплений показал, что ювенилы становятся преобладающей группой и составляют 86 % популяции. Среди взрослых особей на самцов приходится 43 % популяции, при этом генеративных особей среди них не обнаружено. В разнотравно-осоковом лугу около половины популяций представлено ювенилами, из взрослых особей 32 % приходится на самцов, генеративных среди них нет.
В 2009 г. в лиственничниках по сравнению с 2008 г. отмечены следующие изменения: агрегации вида в наших пробах не обнаружены, а половозрастная структура в целом отличалась большей стабильностью в течение сезона и не имела резких изменений соотношений групп. В августе в ерниковом лиственничнике 64 % приходится на взрослых особей, доля самцов составила 56 %, из которых 5 % генеративные. При этом к концу сезона в этом биотопе доля ювенилов значительно снизилась и популяция была представлена в основном взрослыми особями с большой долей самцов (76 %), среди которых генеративных не обнаружено. В остальных биотопах к концу лета зафиксирован рост доли ювенильных особей [Чимитова, 2010].
О периодах размножения X. obscura можно делать выводы, если в этих сроках в половозрастной структуре присутствуют репродуктивные самцы. На сводном графике по всем биотопам можно увидеть более наглядно изменение соотношений различных половозрастных групп в течение сезона (рис. 2). По нашим данным, пик размножения вида наблюдается в первой половине лета. На это указывает преобладание в июне в половозрастной структуре взрослых особей (94 %), большая часть которых (77 %) самцы, при этом 27 % из них репродуктивные. К концу лета в большом количестве появляются ювенильные особи. В августе 2008 г. популяции X. obscura в основном представлены ювенилами, в половозрастной структуре на них приходится 85 %. Самцы составляют в этот срок 42 %, при этом генеративных самцов не обнаружено. К концу сентября (2009 г.) доля ювенилов значительно снижается (14 %), при этом большая часть (75 %) взрослых особей — предгенера-тивные самцы. По-видимому, появившиеся в большом количестве в августе юве-нилы к октябрю завершают рост и уходят на зимовку в предгенеративном состоянии. Достигшие половозрелости к весне или началу следующего лета, особи приступают к размножению. Находка размножающихся самцов в августе 2009 г. в ерниковом лиственничнике позволяет предположить возможность растянутого репродуктивного периода, что требует дальнейших исследований.
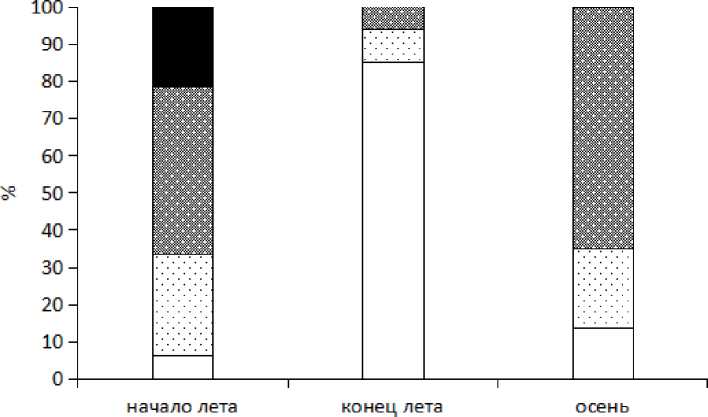
□ juv □ ^ (subadult, adult) И J (subadult) ■ ф adult
Рис. 2 . Половозрастная структура популяций X. obscura юга Витимского плоскогорья (сводная по всем биотопам) (начало лета объединяет данные всех биотопов по июню 2008 г., конец лета — по августу 2008 г., осень — по сентябрю 2009 г.)
Результаты и обсуждение
Среди ногохвосток есть обоеполые виды, партеногенетические виды, состоящие только из самок, и виды, которые представлены и партеногенетическими, и обоеполыми популяциями [Hopkin, 1997]. Исследованию соотношению самцов и самок посвящен ряд работ [Petersen, 1978; Takeda, 1984; Niclasson, Petersen, Parker, 2000; Chahartaghi, Scheu, Ruess, 2006 и др.]. Факторы, от которых зависит это соотношение, не ясны, но предполагается, что оно хотя бы частично связано с абиотическими факторами [Hopkin, 1997]. В целом для обоеполых видов, которым принадлежит X. obscura , характерно преобладание самок [Takeda, 1984; Chahartaghi et al., 2006 и др.]. Например, в сосновых лесах в Японии общее соотношение полов составило 3,1:1 для самок и самцов всех коллембол соответственно [Takeda, 1984]. В дубово-буковых лесах южной Германии самки составляли 65 % взрослых особей у обоеполых коллембол [Chahartaghi et al., 2006].
Полученная в наших исследованиях средняя доля самцов (48 %) несколько выше этих значений, но все же в большинстве сроков на самцов приходится 30–40 %, что типично для гемиэдафических коллембол.
В двух случаях нами обнаружен феномен резкого увеличения доли самцов. В июне 2008 г. (лиственничник разнотравный) увеличение численности самцов (75 %), треть которых генеративная, сопряжено с очень высокой численностью (всего поймано 4 739 экземпляров) и явной агрегированностью этого вида (индекс Лексиса (λ) = 52,4). В сентябре 2009 г. (ерниковый лиственничник) доля самцов поднялась до 76 %, численность была относительно высокая (всего пойман 431 экземпляр), хотя агрегированность была ниже (индекс Лексиса (λ) = 14,3).
Формирование агрегаций известно для многих почвенных членистоногих. Возникновение их у коллембол может объясняться как внешними, так и внутренними факторами [Hopkin, 1997]. Во многих исследованиях выявлено, что внешние факторы, вызывающие скопления ногохвосток, — благоприятные условия влажности, температуры, наличие пищи и т. д. [Christiansen, 1970; Joosse, 1970, 1971; Usher, Hider, 1975; Verhoef, Nagelkerke, 1977 и др.]. Внутренними факторами агрегаций могут выступать размножение и синхронные линьки, имеющие феромонный механизм [Joosse, 1966, 1970; Joosse, Verhoef, 1974; Mertens, Bourgoignie, 1977; Verhoef et al., 1977; Manica et al., 2001 и др.].
Мы предполагаем, что в наших случаях локальное резкое повышение доли самцов в сочетании с высокой численностью популяции (что является по сути агрегацией) может быть связано с размножением. Особенно это проявилось в июне 2008 г. в лиственничнике разнотравном, где была обнаружена феноменально высокая численность самцов в генеративном состоянии. В качестве рабочей гипотезы можно принять, что самцы активнее реагируют на половые феромоны и быстрее скапливаются в местах размножения, чем самки. Данная гипотеза требует более прямых подтверждений, например в лабораторных условиях.
Нам известна только одна работа, оценивающая соотношение полов у коллем-бол в зависимости от плотности популяций. Джуссе [Joosse, 1966] исследовала агрегации Anurida maritima на побережье Нидерландов. Этот вид концентрируется в вертикальных глиняных стенках, выполняющих функции убежищ («гнезд») от набегающей волны. Как в «гнездах», так и вне их самок было больше, чем самцов (75 % и 89 % соответственно). Небольшое увеличение доли самцов в «гнездах» автор объяснял тем, что пища, которая больше необходима самкам, добывается в основном вне «гнезд». В свете наших данных возможна и другая интерпретация — как и в нашем случае, роль может играть групповое размножение.
Обнаруженные в наших исследованиях агрегации X. obscura в ерниковом лиственничнике в августе 2008 г., которые в основном состоят из ювенилов, возможно, связаны с массовой кладкой яиц и последующим вылуплением из них ювенильных особей, имеющих очень медленную скорость перемещения в пространстве.
Заключение
Проведенное исследование позволило выявить некоторые особенности половозрастной структуры и сезонной динамики массового для лесных биотопов юга Витимского плоскогорья вида коллембол — Xenylla obscura .
Хотя по результатам промеров головных капсул подавляющее большинство ювенилов преобладает в наименьшем размерном классе (0–0,1 мм), а большинство взрослых особей относится к среднему классу (0,1–0,14 мм), все же значительный процент взрослых особей перекрывается в 1-м классе с ювенильными особями.
На основании проведенных исследований можно сделать вывод, что основной репродуктивный период у X. obscura приходится на первую половину лета.
Соотношение полов в популяциях X. obscura в большинстве случаев соответствует характерному для коллембол преобладанию самок (30–40 % самцов), однако в отдельные сезоны в разнотравном и ерниковом лиственничниках наблюдались высокие доли самцов (75–81 %). Феноменально высокая численность самцов в генеративном состоянии в агрегациях X. obscura предположительно связана с размножением. Этот аспект требует дальнейшего изучения.
