Эффективность геномной оценки племенной ценности голштинских быков-производителей в сравнении с оценкой по дочерям

Автор: Шендаков А.И.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 2 (71), 2018 года.
Бесплатный доступ
В практику животноводства во всём мире активно внедряется геномный анализ, однако в России этот метод используется только в отдельных случаях, селекционеры придерживаются классического метода оценки быков-производителей по дочерям. В связи с этим целью исследований в данной научной работе являлось сравнение племенной ценности голштинских быков-производителей немецкого происхождения, прошедших геномную оценку и оценку по дочерям. Рассмотрены вопросы эффективности геномного анализа племенной ценности чёрно-пёстрых голштинских и красно-пёстрых голштинских быков-производителей в сравнении с оценкой по дочерям. Дано сравнение величин индексов RZG, RZM, RZE, RZS, RZN, RZD, RZKd, RZR, RZFit и RZRobot при геномной оценке и оценке по дочерям, проанализированы стандартные отклонения и коэффициенты вариации индексов. Определено, что у чёрно-пёстрых голштинских быков-производителей все изученные индексы при геномной оценке (n=432) были выше, чем при оценке по дочерям (n=467), от 2,5 (по RZD) до 14,7 (по RZFit) и 15,9 (по RZG). При этом все отличия были достоверны при p
Голштинская порода, геномная оценка, общий индекс племенной ценности, быки-производители
Короткий адрес: https://sciup.org/147124519
IDR: 147124519 | УДК: 636.234.1.082.232.352636.234.1.082.355-055.6-055.2:636.082.2 | DOI: 10.15217/issn2587-666X.2018.2.52
The efficiency of genomic evaluation of the breeding value of Holstein sires in comparison with the evaluation by daughters
Genomic analysis is actively introduced into the practice of animal breeding in developed countries of the world, but in Russia this method is used only in some cases, and the breeders adhere to the classical method of evaluating sires by daughters. For this reason, the aim of the research in this scientific work was to compare the breeding value of the Holstein sires from the German population, which were genomically evaluated and evaluated by daughters. The scientific article presents the effect of genomic analysis of breeding value of Вlack-and-White and Red-and-White Holstein sires in comparison with the evaluation by daughters. Comparison of the values of the RZG, RZM, RZE, RZS, RZN, RZD, RZKd, RZR, RZFit and RZRobot indices in performing genomic evaluation and evaluation by daughters was made, and standard deviations and coefficients of variation of indices were analyzed. It was determined that the Вlack-and-White Holstein sires had all the indices studied for genomic evaluation (n=432) higher than those for the evaluation by daughters (n=467) - from 2.5 (RZD) to 14.7 (RZFit) and 15.9 (RZG). In this case, all differences were significant at p
Текст научной статьи Эффективность геномной оценки племенной ценности голштинских быков-производителей в сравнении с оценкой по дочерям
Введение. Двадцатый век ознаменовался многими научными открытиями в области биологии и генетики в частности, что отразилось на совершенствовании методов, применяемых в селекции сельскохозяйственных ^ивотных и растений. Если кратко рассмотреть эволюцию этих методов, применяемых в селекции молочного скота, то мо^но выделить несколько этапов: 1) изучение наследуемости и изменчивости по фенотипическим данным с помощью классических методов (Dechow et al, 2001; Dechow et al, 2002 [4, 5]); 2) более подробный анализ так называемых «генетических параметров» (González-Recio et al, 2005 [10]), в том числе генетических корреляций, взаимосвязи «генотип-среда», аддитивной и неаддитивной наследуемости и пр.; 3) маркерная селекция с использованием различных генетических факторов (Dekkers, 2004 [6]), использование в селекции как отдельных генов, так и эритроцитарных факторов, цитогенетика (Eggen, Fries, 1995 [8]); 4) исследование генетических параметров с помощью многофакторного анализа, который позволили выполнять, в том числе, специализированные компьютерные программы (Crawley et al, 2005 [2]); 5) активный поиск генов, ассоциированных с различными продуктивными признаками (Hayes et al, 2009; Fontanesi et al, 2010; de Roos et al, 2011 [3, 9, 1416]); 6) геномная селекция и полногеномный анализ (Habier et al, 2009; Hayes et al, 2009 a, b; Heffner et al, 2009; Jannink et al, 2010; Olson, VanRaden, 2010; Toosi et al, 2010 [11, 12, 17-20]).
Ка^дое научное направление в рамках этих этапов прошло свои уровни развития, отдельные методы были признаны ошибочными, однако сло^но отрицать тот факт, что в настоящее время селекция вышла на принципиально иной уровень получения объективной информации о генотипе и геноме ^ивотных. Так, в настоящее время геномный анализ проводится не только в США, но и в ряде стран мира - в Ирландии (Berry et al, 2009 [1]), Южной Африке (Eui-Soo Kim et al, 2014 [7]), Германии (Habier et al, 2010 [12]), Новой Зеландии (Harris, Johnson , 2010 [13]) и пр. С помощью геномной селекции решаются актуальные проблемы скрещивания местных популяций с голштинами, генетической структуры смешанных популяций, оцениваются перспективы геномной оценки быков-производителей в сравнении с классическими методами оценки племенной ценности и пр.
Таким образом, в течение последних десятилетий генетическому анализу и, в частности, геномной селекции за границей было посвящено много статей известных учёных. Решение проблем геномной селекции крайне необходимо и в России, что подтвер^дает целесообразность наших исследований.
Целью иссле^ований являлось сравнение племенной ценности голштинских быков-производителей немецкого происхо^дения, прошедших геномную оценку и оценку по дочерям. В соответствии с целью были поставлены задачи: 1) сравнить индексы племенной ценности при геномной оценке и оценке по дочерям у чёрно-пёстрых голштинских и красно-пёстрых голштинских быков-производителей; 2) изучить фенотипическую изменчивость индексов племенной ценности при геномной оценке и оценке быков-производителей по дочерям; 3) изучить корреляции ме^ду индексами племенной ценности при геномной оценке и оценке быков-производителей по дочерям; 4) сделать выводы и предло^ения по дальнейшему внедрению геномной оценки в практику молочного скотоводства.
Материалы и мето^ы иссле^ований. Исследования базы данных быков-производителей, семя которых немецкая компания German Genetics International GmbH (GGI) предлагает в 2018 году, были проведены с использованием официальной информации, предоставленной на русифицированном сайте организации [21]. Было дано сравнение племенной ценности голштинских и краснопёстрых голштинских быков-производителей при оценке по дочерям и геномной оценке. Первичная информация по ка^дому быку-производителю для анализа копировалась в компьютерную программу «Microsoft Excel».
B работе были проанализированы следующие индексы племенной ценности, применяемые в Германии:
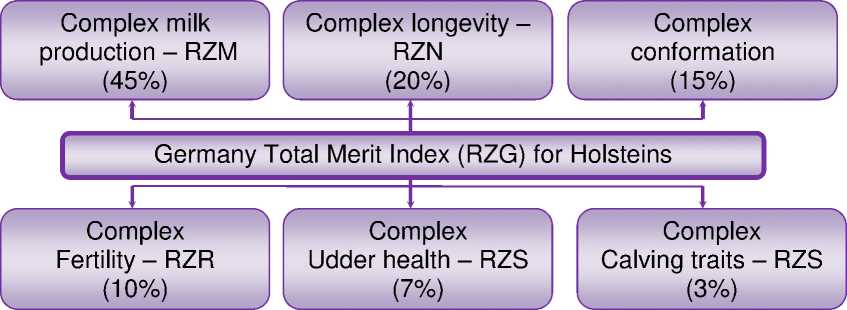
RZG – relative breeding value total merit index, который включает компоненты (индексы) с весом от 3 до 45% (рис. 1);
RZE – relative breeding value conformation, который включает оценку молочного типа (вес признака в индексе 10%), телосло^ения (20%), конечностей (30%) и вымени (40%);
RZD – relative breeding value milking speed, включающий скорость молокоотдачи в кг/мин (вес признака в индексе 50%), мнение владельца о темпераменте – подви^ный, нормальный или сло^ный (50%);
RZKd – relative breeding value calving ease (direct calving ease), индекс, который представляет собой оценку лёгкости первого отёла, в том числе лёгкость отёлов от семени быков-производителей (50%) и их влияние на мёртворо^даемость (50%);
RZFit – relative breeding value fitness, представляющий собой сло^ный индекс, который включает Complex fertility (RZR – 20%), Calving traits maternal (RZKm – 20%), Complex longevity (RZN – 15%), оценку конечностей и вымени (15 и 10% соответственно), Complex udder health (RZS – 10%), Complex milk production (RZM – 10%).
Так^е при оценке эффективности геномной оценки применялся индекс относительной племенной ценности Relative breeding value RZRobot, введённый в популяции для упрощения закрепления быков-производителей к фермам, применяющим автоматизированные и/или роботизированные системы доения. Данный индекс включает в себя скорость молокоотдачи (RZD) (дол^ен быть ≥ 94), количество соматических клеток (RZS), оценку конечностей, располо^ение задних сосков (дол^ен быть ≤ 106), длину сосков (дол^ен быть ≥ 94), оценку вымени (соответственно вес индексов в общем индексе составляет 20, 15, 15, 20, 20 и 10%). Таким образом, этот индекс так^е включал в себя оценку скорости молокоотдачи, количества соматических клеток, располо^ения задних сосков и длины сосков. С точки зрения геномной селекции, подразумевалось, что все признаки, входящие в индексы отбора, могут иметь ассоциации с нуклеотидными последовательностями, обнару^енными в процессе геномного анализа.
- Protein, kg (75%)
- Fat, kg (20%)
- Protein, % (5%)
- Functional herdlife - Auxiliary traits
-Feet and legs (50%)
-Udder (50%) Linear traits and scores each
- 4 conception traits (75%)
- Calving to first insemination (25%)
-somatic cell count
-Calving ease of the daughters (50%) - Still birth rate of the daughters (50%)
Рисунок 1 – Компоненты общего индекса племенной ценности для голштинской породы в Германии (по данным GGI)
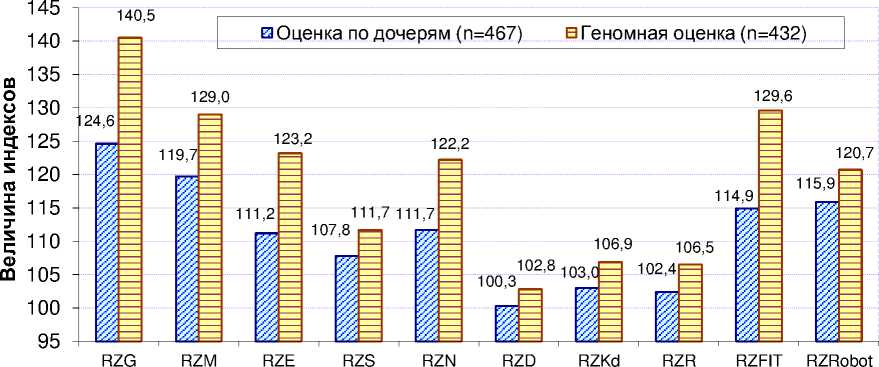
Результаты и их обсу^^ение. Исследования показали (рис. 2), что у чёрно-пёстрых голштинских быков-производителей все изученные индексы при геномной оценке (n=432) были выше, чем при оценке по дочерям (n=467), от 2,5 (по RZD) до 14,7 (по RZFit) и 15,9 (по RZG). При этом все различия были достоверны при p<0,001.
Ин^ексы племенной ценности
Рисунок 2 – Индексы племенной ценности голштинских быков-производителей немецкого происхо^дения в зависимости от метода оценки (2018 г)
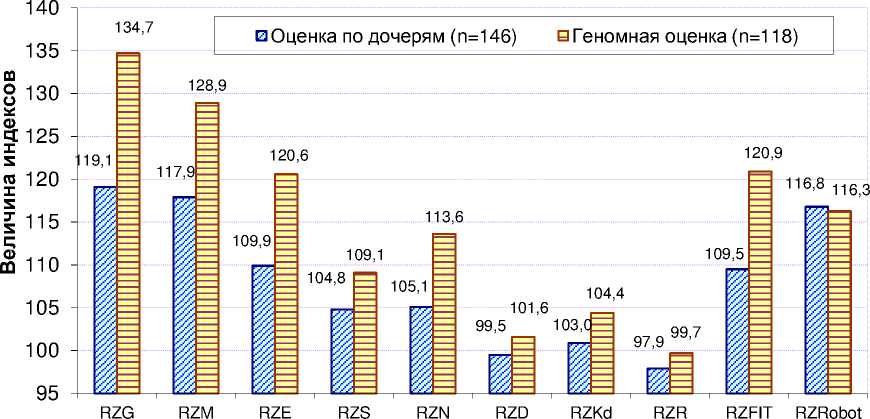
^нализ данных показал (рис. 3), что у красно-пёстрых голштинских быков-производителей, прошедших геномную оценку (n=118), почти все индексы племенной ценности были выше, чем у быков-производителей, оцененных по дочерям (n=146). Отличия составили от 1,4 (по RZKd) до 15,6 (по RZG). Исключением стал индекс RZRobot, который был ни^е при геномной оценке.
Ин^ексы племенной ценности
Рисунок 3 – Индексы племенной ценности красно-пёстрых голштинских быков-производителей немецкого происхо^дения в зависимости от метода оценки (2018 г)
B целом, по RZD, RZR и RZRobot достоверных различий в сравнении с классической оценкой получено не было (табл. 1).
Таблица 1 – Фактическое значение критерия Стьюдента при оценке племенной ценности голштинских быков-производителей немецкого происхо^дения
|
Порода |
Метод оценки |
Голов (n) |
Фактическое значение критерия Стьюдента ( tst ) |
|||||||||
|
(5 N CC |
s CC |
LU N CC |
co N CC |
CC |
CC |
i CC |
CC N CC |
CC |
о -Q О CC |
|||
|
Голштинская |
По дочерям |
467 |
28,1 |
13,1 |
18,7 |
6,9 |
18,5 |
4,4 |
9,2 |
6,4 |
26,0 |
8,5 |
|
Геномная |
432 |
Bсе отличия по индексам достоверны при |
p<0,001 |
|||||||||
|
Краснопёстрая голштинская |
По дочерям |
146 |
16,5*** |
9,1*** |
9,4*** |
4,3*** |
9,2*** |
1,8 |
3,8*** |
1,7 |
11,5*** |
0,54 |
|
Геномная |
118 |
|||||||||||
Примечание: *** - p<0,001.
Стандартные отклонения (σ) изученных индексов (табл. 2) у быков-производителей двух пород колебались в пределах от 6,5-6,6 до 10,1-11,1, однако при геномной оценке они были в основном ни^е, чем при оценке по дочерям. Ме^ду тем, стандартные отклонения общего основного индекса племенной ценности (RZG) при геномной оценке были выше на 1,6 и 2,8. Схо^ая тенденция просле^ивалась по коэффициенту фенотипической изменчивости
(C v ). B частности, наибольший C v был получен по индексу RZE при оценке по дочерям (9,0-10,1%). Геномная оценка дала 5,9-6,0% изменчивости по индексу RZKd, хотя общий индекс RZG при геномной оценке варьировал сильнее, чем при оценке по дочерям (на 0,4-3,2%).
Таблица 2 – Изменчивость племенной ценности голштинских быков-производителей немецкого происхо^дения при оценке по дочерям и геномной оценке
|
Порода |
Метод оценки |
n |
Генетико-статистические параметры индексов ( σ ) |
|
RZG"I RZM RZEп RZSп RZN1 RZD 1 RZKd"I RZRп RZFIT"1 RZRobot |
Стандартное отклонение индексов племенной ценности (σ)
|
Голштинская |
По дочерям |
467 |
7,8 |
9,9 |
10,0 |
9,2 |
8,4 |
9,3 |
6,7 |
10,1 |
8,9 |
7,9 |
|
Геномная |
432 |
9,4 |
9,8 |
8,0 |
8,2 |
7,8 |
8,2 |
6,4 |
8,3 |
8,3 |
7,9 |
|
|
Красно-пёстрая голштинская |
По дочерям |
146 |
6,5 |
9,4 |
11,1 |
9,0 |
8,3 |
9,8 |
6,8 |
9,7 |
8,9 |
7,8 |
|
Геномная |
118 |
9,3 |
9,5 |
7,7 |
7,3 |
6,6 |
8,6 |
7,2 |
7,4 |
7,6 |
8,2 |
|
|
Вариация индексов племенной ценности (C v ) |
||||||||||||
|
Голштинская |
По дочерям |
467 |
6,3 |
8,2 |
9,0 |
8,5 |
7,5 |
9,3 |
6,5 |
9,9 |
7,7 |
6,8 |
|
Геномная |
432 |
6,7 |
7,6 |
6,5 |
7,4 |
6,4 |
8,0 |
5,9 |
7,8 |
6,4 |
6,5 |
|
|
Красно-пёстрая голштинская |
По дочерям |
146 |
5,4 |
8,0 |
10,1 |
8,6 |
7,9 |
9,8 |
6,7 |
9,9 |
8,1 |
6,7 |
|
Геномная |
118 |
8,6 |
7,4 |
7,1 |
7,5 |
6,4 |
8,3 |
6,0 |
7,4 |
6,3 |
7,0 |
|
|
Ошибка индексов племенной ценности ( ±m ) |
||||||||||||
|
Голштинская |
По дочерям |
467 |
0,4 |
0,5 |
0,5 |
0,4 |
0,4 |
0,4 |
0,3 |
0,5 |
0,4 |
0,4 |
|
Геномная |
432 |
0,4 |
0,5 |
0,4 |
0,4 |
0,4 |
0,4 |
0,3 |
0,4 |
0,4 |
0,4 |
|
|
Красно-пёстрая голштинская |
По дочерям |
146 |
0,5 |
0,8 |
0,9 |
0,7 |
0,7 |
0,8 |
0,6 |
0,8 |
0,7 |
0,6 |
|
Геномная |
118 |
0,8 |
0,9 |
0,7 |
0,7 |
0,6 |
0,8 |
0,7 |
0,7 |
0,7 |
0,7 |
|
^нализ коэффициентов корреляций (r±m r ) ме^ду индексами племенной ценности при геномной оценке и оценке по дочерям (табл. 3) показал, что при геномной оценке большинство корреляций были выше независимо от породы. Так, у чёрно-пёстрых голштинских быков корреляции RZG (relative breeding value total merit index) и RZM при геномной оценке и оценке по дочерям составили 0,818±0,016 и 0,717±0,022 соответственно. У красно-пёстрых голштинских быков эти коэффициенты корреляций составили 0,861±0,023 и 0,567±0,056 соответственно. Корреляции индексов RZN и RZG при геномной оценке и оценке по дочерям у чёрно-пёстрых голштинских быков-производителей составили 0,545±0,034 и 0,385±0,039, а у красно-пёстрых голштинов – 0,523±0,067 и 0,237±0,078 соответственно. Высокие корреляции так^е были получены ме^ду RZG и RZFit (relative breeding value fitness) у быков-производителей обеих пород, а геномная оценка была лучше по данным корреляциям у чёрно-пёстрой голштинской породы нa 0,213 и у красно-пёстрой голштинской – на 0,291 (при достоверности p<0,001).
Bмecтe c тeм, были получены отрицательные корреляции ме^ду индексом, характеризующим молочную продуктивность дочерей (RZM), и большинством индексов – нa уровне до -0,366±0,040 у чёрно-пёстрых голштинов и до -0,562±0,090 у красно-пёстрых голштинов. При геномной оценке, однако, отрицательные коэффициенты не были столь отрицательными, они возросли до -0,281±0,044 и -0,172±0,90, а общее количество отрицательных корреляций ме^ду этими индексами снизилось с 7 и 6 до 5 и 3 соответственно.
Таблица 3 – Корреляции индексов племенной ценности голштинских быков-производителей немецкого происхо^дения при геномной оценке и оценке по дочерям
|
го го: о о. о EZ |
^1 0) 9> о |
Индексы |
Коэффициенты корреляций и их ошибки, r (слева) и ±mr (справа) |
|||||||||
|
RZG |
RZM |
RZE |
RZS |
RZN |
RZD |
RZKd |
RZR |
RZFIT |
RZRo-bot |
|||
|
IX го О н □ го о I— |
о. гС Ф со 51 4 с о EZ |
RZG |
- |
0,022 |
0,044 |
0,045 |
0,039 |
0,046 |
0,045 |
0,046 |
0,037 |
0,046 |
|
RZM |
0,717 |
- |
0,045 |
0,044 |
0,044 |
0,046 |
0,046 |
0,040 |
0,043 |
0,042 |
||
|
RZE |
0,196 |
-0,167 |
- |
0,046 |
0,046 |
0,046 |
0,046 |
0,046 |
0,037 |
0,034 |
||
|
RZS |
0,155 |
-0,185 |
0,079 |
- |
0,041 |
0,039 |
0,046 |
0,046 |
0,040 |
0,041 |
||
|
RZN |
0,385 |
-0,222 |
0,126 |
0,339 |
- |
0,046 |
0,043 |
0,039 |
0,024 |
0,046 |
||
|
RZD |
-0,070 |
0,003 |
0,034 |
-0,397 |
-0,030 |
- |
0,046 |
0,046 |
0,046 |
0,046 |
||
|
RZKd |
0,143 |
-0,030 |
-0,002 |
0,045 |
0,255 |
0,095 |
- |
0,045 |
0,041 |
0,046 |
||
|
RZR |
0,098 |
-0,366 |
≈0 |
0,061 |
0,396 |
-0,029 |
0,157 |
- |
0,026 |
0,045 |
||
|
RZFIT |
0,452 |
-0,240 |
0,431 |
0,382 |
0,694 |
-0,078 |
0,321 |
0,664 |
- |
0,039 |
||
|
RZRobot |
-0,027 |
-0,288 |
0,516 |
0,307 |
0,081 |
0,118 |
-0,006 |
0,173 |
0,387 |
- |
||
|
IX |й и 1— |
RZG |
- |
0,016 |
0,043 |
0,043 |
0,034 |
0,048 |
0,048 |
0,048 |
0,027 |
0,043 |
|
|
RZM |
0,818 |
- |
0,048 |
0,048 |
0,048 |
0,048 |
0,048 |
0,044 |
0,047 |
0,048 |
||
|
RZE |
0,340 |
0,022 |
- |
0,045 |
0,044 |
0,047 |
0,047 |
0,048 |
0,034 |
0,032 |
||
|
RZS |
0,325 |
-0,020 |
0,244 |
- |
0,038 |
0,044 |
0,048 |
0,048 |
0,033 |
0,040 |
||
|
RZN |
0,545 |
0,058 |
0,293 |
0,461 |
- |
0,048 |
0,048 |
0,043 |
0,020 |
0,040 |
||
|
RZD |
-0,011 |
-0,014 |
0,140 |
-0,308 |
0,022 |
- |
0,048 |
0,048 |
0,048 |
0,045 |
||
|
RZKd |
-0,035 |
-0,067 |
-0,160 |
-0,030 |
0,057 |
0,044 |
- |
0,047 |
0,047 |
0,047 |
||
|
RZR |
0,055 |
-0,281 |
-0,049 |
0,078 |
0,326 |
0,003 |
0,156 |
- |
0,034 |
0,047 |
||
|
RZFIT |
0,665 |
0,149 |
0,535 |
0,546 |
0,768 |
0,001 |
0,071 |
0,532 |
- |
0,035 |
||
|
RZRobot |
0,334 |
-0,011 |
0,578 |
0,407 |
0,403 |
0,260 |
-0,175 |
0,120 |
0,515 |
- |
||
|
IX го О н □ с; О IX го о. 1— :ф С 6 I го о. ^ |
О- СО 0) ^ 4 с О EZ |
RZG |
- |
0,056 |
0,070 |
0,082 |
0,078 |
0,082 |
0,083 |
0,082 |
0,068 |
0,082 |
|
RZM |
0,567 |
- |
0,080 |
0,072 |
0,059 |
0,083 |
0,083 |
0,057 |
0,064 |
0,082 |
||
|
RZE |
0,382 |
-0,199 |
- |
0,080 |
0,078 |
0,081 |
0,081 |
0,079 |
0,053 |
0,074 |
||
|
RZS |
0,097 |
-0,361 |
0,186 |
- |
0,071 |
0,075 |
0,083 |
0,079 |
0,061 |
0,078 |
||
|
RZN |
0,237 |
-0,540 |
0,245 |
0,380 |
- |
0,082 |
0,081 |
0,044 |
0,032 |
0,078 |
||
|
RZD |
-0,121 |
0,060 |
-0,122 |
-0,304 |
-0,111 |
- |
0,082 |
0,082 |
0,079 |
0,065 |
||
|
RZKd |
0,065 |
0,003 |
-0,161 |
-0,001 |
0,149 |
-0,089 |
- |
0,082 |
0,082 |
0,075 |
||
|
RZR |
0,115 |
-0,562 |
0,208 |
0,230 |
0,688 |
-0,087 |
0,078 |
- |
0,040 |
0,082 |
||
|
RZFIT |
0,419 |
-0,476 |
0,598 |
0,515 |
0,784 |
-0,209 |
0,111 |
0,721 |
- |
0,077 |
||
|
RZRobot |
0,153 |
-0,119 |
0,331 |
0,232 |
0,239 |
0,455 |
-0,312 |
0,090 |
0,267 |
- |
||
|
IX 2 со ф ^ 1— |
RZG |
- |
0,023 |
0,078 |
0,087 |
0,067 |
0,092 |
0,091 |
0,091 |
0,045 |
0,090 |
|
|
RZM |
0,861 |
- |
0,091 |
0,092 |
0,091 |
0,092 |
0,090 |
0,090 |
0,084 |
0,092 |
||
|
RZE |
0,398 |
0,138 |
- |
0,091 |
0,086 |
0,089 |
0,092 |
0,092 |
0,061 |
0,070 |
||
|
RZS |
0,252 |
0,003 |
0,111 |
- |
0,077 |
0,082 |
0,092 |
0,092 |
0,076 |
0,085 |
||
|
RZN |
0,523 |
0,097 |
0,273 |
0,404 |
- |
0,092 |
0,092 |
0,082 |
0,043 |
0,082 |
||
|
RZD |
-0,037 |
-0,053 |
0,169 |
-0,335 |
0,055 |
- |
0,092 |
0,092 |
0,092 |
0,084 |
||
|
RZKd |
0,122 |
0,139 |
0,030 |
-0,094 |
-0,056 |
0,006 |
- |
0,091 |
0,091 |
0,084 |
||
|
RZR |
0,107 |
-0,172 |
-0,062 |
0,007 |
0,330 |
-0,065 |
0,106 |
- |
0,067 |
0,091 |
||
|
RZFIT |
0,710 |
0,290 |
0,582 |
0,413 |
0,728 |
0,015 |
0,116 |
0,521 |
- |
0,083 |
||
|
RZRobot |
0,165 |
-0,031 |
0,487 |
0,286 |
0,336 |
0,308 |
-0,308 |
-0,134 |
0,314 |
- |
||
^нализ достоверности различий коэффициентов корреляций при геномной оценке над коэффициентами корреляций, полученными при оценке по дочерям (табл. 4), показал, что у чёрно-пёстрых голштинских быков-производителей RZG достоверно поло^ительнее коррелировал с RZM (p<0,001), RZE (p<0,05), RZS (p<0,001), RZN (p<0,01), RZFit (p<0,001) и RZRobot (p<0,001). Превосходство в данном случае составило от +0,101 до +0,361. У красно-пёстрых голштинских быков-производителей общий индекс RZG дал ухудшение только по связи с RZR (-0,008), однако превосходство по другим коэффициентам корреляций было достоверно на уровне p<0,05-p<0,001 только при взаимосвязи RZG-RZM (+0,294), RZG-RZN (+0,286) и RZG-RZFit (+0,291). Cущественным следует считать то, что геномная оценка дала увеличение большинства корреляций RZM с другими индексами в изученных породах (p<0,05-p<0,001). Особенно значимый эффект был получен по RZM-RZFit (+0,766) и RZM-RZN (+0,637) у краснопёстрых голштинов.
Таблица 4 – Различия коэффициентов корреляций при геномной оценке от коэффициентов корреляций при оценке по дочерям и достоверность различий
|
Порода |
Индексы |
Различия коэффициентов кор |
реляций и достоверность, +/- (слева) и p (справа) |
||||||||
|
RZG |
RZM |
RZE |
RZS |
RZN |
RZD |
RZKd |
RZR |
RZFIT |
RZRobot |
||
|
J0 СО р. II о — О s I н CL 3 Т О |
RZG |
- |
p<0,001 |
p<0,05 |
p<0,01 |
p<0,01 |
p>0,05 |
p<0,01 |
p>0,05 |
p<0,001 |
p<0,001 |
|
RZM |
+0,101 |
- |
p<0,01 |
p<0,05 |
p<0,001 |
p>0,05 |
p>0,05 |
p>0,05 |
p<0,001 |
p<0,001 |
|
|
RZE |
+0,144 |
+0,189 |
- |
p<0,05 |
p>0,05 |
p>0,05 |
p<0,05 |
p>0,05 |
p<0,05 |
p<0,01 |
|
|
RZS |
+0,170 |
+0,165 |
+0,165 |
- |
p<0,05 |
p>0,05 |
p>0,05 |
p>0,05 |
p<0,01 |
p>0,05 |
|
|
RZN |
+0,160 |
+0,280 |
+0,167 |
+0,122 |
- |
p>0,05 |
p<0,05 |
p>0,05 |
p<0,05 |
p<0,001 |
|
|
RZD |
+0,059 |
-0,017 |
+0,106 |
+0,089 |
+0,052 |
- |
p>0,05 |
p>0,05 |
p>0,05 |
p<0,05 |
|
|
RZKd |
-0,178 |
-0,037 |
-0,158 |
-0,075 |
-0,198 |
-0,051 |
- |
p>0,05 |
p<0,001 |
p<0,05 |
|
|
RZR |
-0,043 |
+0,085 |
-0,049 |
+0,017 |
-0,070 |
+0,032 |
-0,001 |
- |
p<0,05 |
p>0,05 |
|
|
RZFIT |
+0,213 |
+0,389 |
+0,104 |
+0,164 |
+0,074 |
+0,079 |
-0,250 |
-0,132 |
- |
p<0,05 |
|
|
RZRobot |
+0,361 |
+0,277 |
+0,062 |
+0,100 |
+0,322 |
+0,142 |
-0,169 |
-0,053 |
+0,128 |
- |
|
|
0) см ^ч о И ГО 5 |
RZG |
- |
p<0,001 |
p>0,05 |
p>0,05 |
p<0,01 |
p>0,05 |
p>0,05 |
p>0,05 |
p<0,001 |
p>0,05 |
|
RZM |
+0,294 |
- |
p<0,01 |
p<0,01 |
p<0,001 |
p>0,05 |
p>0,05 |
p<0,001 |
p<0,001 |
p>0,05 |
|
|
RZE |
+0,016 |
+0,337 |
- |
p>0,05 |
p>0,05 |
p<0,05 |
p>0,05 |
p<0,05 |
p>0,05 |
p>0,05 |
|
|
RZS |
+0,155 |
+0,364 |
-0,075 |
- |
p>0,05 |
p>0,05 |
p>0,05 |
p>0,05 |
p>0,05 |
p>0,05 |
|
|
RZN |
+0,286 |
+0,637 |
+0,028 |
+0,024 |
- |
p>0,05 |
p>0,05 |
p<0,001 |
p>0,05 |
p>0,05 |
|
|
RZD |
+0,084 |
-0,113 |
+0,291 |
-0,031 |
+0,166 |
- |
p>0,05 |
p>0,05 |
p>0,05 |
p>0,05 |
|
|
RZKd |
+0,057 |
+0,136 |
+0,191 |
-0,093 |
-0,205 |
+0,095 |
- |
p>0,05 |
p>0,05 |
p>0,05 |
|
|
RZR |
-0,008 |
+0,390 |
-0,270 |
-0,223 |
-0,358 |
+0,022 |
+0,028 |
- |
p<0,05 |
p>0,05 |
|
|
RZFIT |
+0,291 |
+0,766 |
-0,016 |
-0,102 |
-0,056 |
+0,224 |
+0,005 |
-0,200 |
- |
p>0,05 |
|
|
RZRobot |
+0,012 |
+0,088 |
+0,156 |
+0,054 |
+0,097 |
-0,147 |
+0,004 |
-0,224 |
+0,047 |
- |
|
B группе чёрно-пёстрых голштинских быков-производителей достоверное ухудшение коэффициентов корреляций при геномной оценке было получено по взаимосвязи RZKd-RZG (p<0,01), RZKd-RZE (p<0,05), RZKd-RZN (p<0,05), RZKd-RZFit (p<0,001), RZKd-RZRobot (p<0,05), со всеми остальными индексами RZKd при геномной оценке так^е дал ухудшение корреляций. Достоверно ху^е при геномной оценке была корреляция RZR-RZFit (на -0,132 при p<0,05). При этом в группе красно-пёстрых голштинских быков-производителей геномная оценка племенной ценности дала достоверное ухудшение корреляций ме^ду RZR и RZE, RZR и RZN, RZR и RZFit (до -0,270, -0,358 и -0,200 соответственно при p<0,05-0,001).
B дополнение к сказанному следует отметить, что при анализе силы влияния изученных факторов на величины индексов и их вариабельность с помощью двухфакторного дисперсионного анализа нами было определено (рис. 4), что фактор «метод оценки» дал от 9,5 до 17,2%. Cила влияния неучтённых факторов на индексы племенной ценности и их вариабельность достигала 81,689,3%, что, с одной стороны, подтвер^дает некоторый эффект геномной оценки, а с другой – необходимость учёта целого ряда генетических и паратипических факторов, к которым мо^но отнести линии, условия кормления, технологии содер^ания и доения и пр. Bполне возмо^но, что сами индексы племенной ценности, применяемые при оценке быков-производителей, ну^даются в совершенствовании и модернизации.
Фактор "порода" (^)
Фактор "порода-метод оценки" (^В)
Фактор "метод оценки" (В)
Неучтённые фаторы (е)
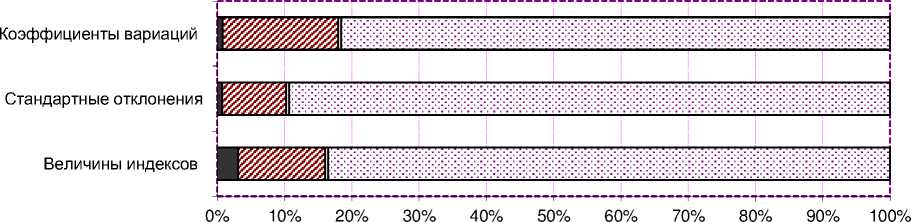
Рисунок 4 – Сила влияния факторов на величины индексов и их вариабельность (двухфакторный анализ в ортогональном комплексе, a=2, b=2, n=10, N=40)
Выво^ы. Таким образом, геномная оценка племенной ценности голштинских быков-производителей даёт поло^ительный результат, однако не все индексы племенной ценности при геномной оценке могут превосходить аналогичные индексы при оценке по дочерям. Геномная оценка становится менее эффективной по тем признакам и индексам, которые, судя по всему, в большей мере подвер^ены влиянию паратипических факторов. К таким индексам следует отнести RZD, RZR и RZRobot , т.е. те индексы, которые включают в себя скорость молокоотдачи, количество соматических клеток в молоке, мнение владельца фермы о темпераменте коровы и пр. Однако геномная оценка мо^ет способствовать получению ^елательных корреляций ме^ду индексами племенной ценности независимо от породы, особенно это ощутимо по индексу RZM, характеризующему молочную продуктивность скота. Полагаться, вместе с тем, только на одну геномную оценку племенной ценности быков-производителей не следует, поскольку вполне очевидно, что геномная оценка мо^ет достоверно вести к ухудшению некоторых ^елательных корреляций ме^ду индексами племенной ценности, особенно это касается RZKd (relative breeding value calving ease), RZFit (relative breeding value fitness) и RZRobot, то есть теми индексами, которые характеризуют некоторые функциональные качества, воспроизводительную функцию, лёгкость отёлов, пригодность к роботизированному доению и пр. Bполне очевидно, что для более подробной информации учёным и практикам, заинтересованным в генетическом прогрессе признаков данных пород, целесообразно рекомендовать при использовании гeномного анализа проведение анализа генотипической и паратипической изменчивости признаков и индексов отбора, генотипических и паратипических корреляций ме^ду индексами племенной ценности. В перспективе при внедрении геномной селекции в отечественное молочное скотоводство следует обращать внимание на данные факты.
Список литературы Эффективность геномной оценки племенной ценности голштинских быков-производителей в сравнении с оценкой по дочерям
- Berry D., Kearney F., Harris B. Genomic selection in Ireland//Interbull, Uppsala, Sweden. 2009. № 39. Р. 29-34.
- Crawley A.M., Mallard B., Wilkie B.N. Genetic selection for high and low immune response in pigs: Effects on immunoglobulin isotype expression//Vet. Immunol. Immunopathol. 2005. № 108. Р. 71-76.
- de Roos A.P.W., Schrooten C., Veerkamp R.F., van Arendonk J.A.M. Effects of genomic selection on genetic improvement, inbreeding, and merit of young versus proven bulls//J. Dairy Sci. 2011. № 94. Р. 1559-1567.
- Dechow C.D., Rogers G.W., Clay J.S. Heritability and correlations among body condition scores, production traits, and reproductive performance//J. Dairy Sci. 2001. № 84. Р. 266-275.
- Dechow C.D., Rogers G.W., Clay J.S. Heritability and correlations among body condition score loss, body condition score, production and reproductive performance//J. Dairy Sci. 2002. № 85. Р. 3062-3070.
- Dekkers J.C.M. Commercial application of marker-and gene-assisted selection in livestock: Strategies and lessons. J. Anim. Sci. 2004. № 82 (E. Suppl.). Р. E313-E328.
- Eui-Soo Kim, Max F. Rothschild Genomic adaptation of admixed dairy cattle in East Africa//Frontiers in Genetics. 2014. December (Volume 5).
- Eggen A., Fries R. An integrated cytogenetic and meiotic map of the bovine genome//Anim. Genet. 1995. № 26. Р. 215-236.
- Fontanesi L., Scotti E., Russo V. Analysis of SNPs in the KIT gene of cattle with different coat colour patterns and perspectives to use these markers for breed traceability and authentication of beef and dairy products//Ital. J. Anim. Sci. 2010. № 9. Р. e42.
- González-Recio O., Ugarte C., Alenda R. Genetic analysis of an artificial insemination progeny test program//J. Dairy Sci. 2005. № 88. Р. 783-789.
- Habier D., Fernando R.L., Dekkers JCM. Genomic selection using low-density marker panels//Genetics. 2009. № 182. Р. 343-353.
- The impact of genetic relationship information on genomic breeding values in German Holstein cattle/D. Habier, J. Tetens, F-R Seefried, P. Lichtner, G. Thaller//Genet. Sel. Evol. 2010. № 42. Р. 5.
- Harris B.L., Johnson D.L. Genomic predictions for New Zealand dairy bulls and integration with national genetic evaluation//J. Dairy Sci. 2010. № 93. Р. 1243-1252.
- Accuracy of genomic breeding values in multi-breed dairy cattle populations/B. Hayes, P. Bowman, A. Chamberlain, K. Verbyla, M. Goddard//Genet. Sel. Evol. 2009. № 41. Р. 51.
- Invited review: Genomic selection in dairy cattle: Progress and challenges/B.J. Hayes, P.J. Bowman, A.J. Chamberlain, M.E. Goddard//J. Dairy Sci. 2009. № 92. Р. 433-443.
- Accuracy of genomic selection: Comparing theory and results/B.J. Hayes et al.//Proc. Assoc. Advmt. Anim. Breed. Genet. 2009. № 18. Р. 34-37.
- Heffner E., Sorrells M., Jannink J. Genomic selection for crop improvement//Crop Sci. 2009. № 49. Р. 1-12.
- Jannink J-L., Lorenz A.J., Iwata H. Genomic selection in plant breeding: From theory to practice//Brief. Funct. Genomics. 2010. № 9. Р. 166-177.
- Olson K.M., VanRaden P.M. Multibreed genomic evaluation of dairy cattle//J. Dairy Sci. 2010. № 93 (E-Suppl. 1). Р. 471.
- Toosi A., Fernando R.L., Dekkers J.C.M. Genomic selection in admixed and crossbred populations//J. Anim. Sci. 2010. № 88. Р. 32-46.
- URL: http://www.ggi.de/ru/glavnaja/(дата обращения 04.01.2018).
