Экологическая характеристика лотической системы на примере малых рек Волжского бассейна: методологические подходы исследований

Автор: Зинченко Татьяна Дмитриевна, Промахова Екатерина Васильевна, Головатюк Лариса Владимировна, Абросимова Элина Владимировна, Попченко Тимур Викторович, Шитиков Владимир Кириллович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-2 т.20, 2018 года.
Бесплатный доступ
В работе дается обзор результатов многолетних исследований водотоков Волжского бассейна с акцентом на анализ подходов к экологической классификации рек. Обсуждаются методы оценки экологического состояния лотических систем как информационной основы обоснования региональных критериев оценки качества поверхностных вод по гидролого-гидрохимическим и биологическим характеристикам планктонных и донных сообществ.
Малые реки, классификация рек, экологическое состояние, планктонные и донные сообщества, методы исследования
Короткий адрес: https://sciup.org/148314052
IDR: 148314052 | УДК: 574.6+504.4
Environmental characteristics of the lothic system on the example of small rivers of the Volga river basin: methodological approaches
In this work, the authors review the results of long-time studies of the watercourses of the Volga River Basin with an emphasis on analysis of approaches to ecological classification of rivers. We discuss methods for assessing the ecological state of lotic systems as an information basis for justifying regional criteria for assessing the quality of surface waters by the hydrological, hydrochemical and biological characteristics of plankton and benthic communities.
Текст научной статьи Экологическая характеристика лотической системы на примере малых рек Волжского бассейна: методологические подходы исследований
связи с тем, что малые реки являются элементом более крупных каскадных систем, они формируют фоновый уровень гидролого-биологических характеристик средних и крупных рек. Особенностью рек Волжского бассейна является наличие подпорных явлений устьевых участков при прохождении максимальных расходов воды на крупных реках в период половодья или паводков, что приводит к морфологическим изменениям русел малых рек [3] и, как следствие, изменению биотических компонентов экосистемы рек.
Основная задача экологических исследований состоит в накоплении, систематизации и анализе информации, результатом которых являются оценка качества изучаемых экосистем; выявление причин наблюдаемых и вероятных структурно-функциональных изменений биотических компонентов, прогноз устойчивости экосистем и параметров допустимой нагрузки на гидроэкосистемы. До настоящего времени не существует универсального метода оценки экологического состояния лотических систем. Одним из способов решения этой задачи может служить комплексная количественная оценка факторов воздействия, вызывающих неблагополучное состояние экосистемы.
Идея о том, что можно изолировать какой-то участок реки как ограниченную и охраняемую территорию и таким образом использовать его как репрезентативный эталон биологического разнообразия, не имеет ничего общего с оценкой, например, экологического состояния водного объекта. [4].
Классификацию экологического состояния рек осуществляется обычно на основе различных подходов, включая анализ измеряемых физикохимических, гидрологических и биотических показателей рек [5, 6]. Например, в Великобритании, Канаде и др. странах ближнего и дальнего зарубежья, а также и в России используют многомерные классификации сообществ макробеспозвоночных на различных участках рек для разработки системы биологической оценки качества вод (система RIVPACS [7, 8, 9, 10]). Австралийская речная система оценки качества вод (AUSRIVAS), также основанная на принципах и методологии RIVPACS (River Invertebrate Prediction and Classification System) позволяет сравнить наблюдаемое разнообразие и богатство сообществ макробеспозвоночных с богатством, полученным по измерениям на различных участках рек [4, 6, 11].
Для исследованных речных систем мы основывались, преимущественно, на данных по биоразнообразию планктонных и донных и сообществ, полученных в ходе многолетнего мониторинга. Достаточно трудно четко разделить реки на сообщества различного типа, поскольку, используя время прохождения различных стадий своего жизненного цикла, большая часть видов адаптирована к обитанию в определенных биотопах или микробиотопах при выявлении уровня воздействия абиотических и биотических факторов. Анализируя современные пресноводные классификации можно констатировать, что используют обычно порядок рек как один из разделов информации при многомерном картировании [12, 13]. Однако порядок реки можно считать субъективной мерой, которая изменяется в зависимости от выбора конкретного масштаба карты региона [14, 15]. Поскольку исследованные нами высокоминерализованные реки Приэльтонья являются реками первого порядка [16], мы для их типологической оценки использовали водосборную площадь, расход воды, размер частиц субстрата, изменение концентрации осадочных пород, уклон, эрозию русла и берегов. В теории динамического равновесия [2, 17] используется набор основных гидрологических и гидрофизических характеристик рек (то есть, ширину, глубину, наличие осадочных включений, расход воды, электропроводность и др.) для объяснения регулярности изменений в нижнем течении рек в связи с изменением площади водосбора и балансом между процессами эрозии и устойчивостью грунтов, подвергающихся размыванию, влиянию подпорных вод и основного руслового потока реки.
Продольное зонирование лотических систем и связанная с ним оценка состояния речных систем, основанная на гидролого-биологических принципах, достаточно часто используется, что- бы оценивать типы рек по их экологическому состоянию [5, 18].
ТЕОРЕТИЧЕСКИЕ И МЕТОДОЛОГИЧЕСКИЕ ПОДХОДЫ К ОЦЕНКЕ ЭКОЛОГИЧЕСКОГО СОСТОЯНИЯ РЕК
С истема классификации рек [19] применялась на постулате, что гидрологическая сеть формируется согласно законам физики речных процессов. Участки долины реки иерархически классифицировались в сходные морфологические группы в зависимости от значений продольного уклона, поперечного профиля, различия в генезисе вод, формирующих сток малых рек. При этом учитываются такие естественные гидрологические объекты, как запруды, перекаты и стремнины. Неявное предположение этой системы состоит в том, что если восстанавливается физическая структура непосредственно самого потока, то одновременно и восстанавливаются важнейшие (биотические) компоненты экосистемы. Хотя такие методы классификации широко используются в Соединенных Штатах Америки для проектов восстановления рек [5], система критикуется за то, что в ней не рассматриваются связи экосистемы с окружающей средой, а также за то, что не учитывается динамичность видов, отрицание стохастических факторов при видообразовании и способности видов создавать множественные устойчивые ассоциации [20]. Таким образом, существует несколько классификаций геоморфологии потока. Нельзя не согласиться с мнением о том, что физические и соответствующие биотические процессы в реках находятся в динамическом «равновесии» или сбалансированном континуальном состоянии. Использование математического равновесия в экологии может быть связано с метафорой “экологического баланса”, которая основана на идее, что природные компоненты стремятся скомпенсировать влияние различных противодействующих сил, что выражается в инерционности сообществ и регулярных флуктуациях популяций и видового состава [21]. Эта метафора “экологический баланса” играет фундаментальную роль во многих областях, и, по всей вероятности, окрашивает такой нейтральный термин как “динамическое равновесие” дополнительными положительными и отрицательными оттенками [21, 22]. В это же время стало очевидным, что продольное и боковое движение воды вдоль потоков создает питательную спираль - Nutrient Spiralling Concept, NSC [23], которая транспортирует питательные вещества в нижнее течение рек [5]. Известна и концепция “последовательного дисконтинуума” [24], которая предполагает наличия разрывов в континууме, например, запруды, бобровые плотины, пруды, водохранилища, которые нарушают естественную продольную закономерность.
Есть предположение, что рост биомассы и разнообразия в воде рек зависит от связи боковой приточности между поймой и руслом реки, что может происходить во время периодических наводнений [25]. Ward [26] описал четыре размерных структуры для проточных вод, которые включали продольные, поперечные, вертикальные и временные координаты. Можно привести концептуальную модель, связывающую уровень флуктуаций потока с процессами изменения видовой структуры речных систем [27]. Рабочая гипотеза состояла в том, что высокая изменчивость или непредсказуемые режимы потока приводят к такой физической среде, в которой абиотические процессы - главные факторы, определяют структуру сообществ лотической экосистемы.
Тезис о том, что широкий диапазон гидрологической изменчивости необходим для поддержания экологической целостности лотических экосистем, является актуальным и вполне подтвержденным в практике экологических исследований малых рек Волжского бассейна [2]. Очевидно, чтобы лучше всего сохранить и восстановить экосистему потока, необходимо восстановить режим потока, соответствующий не-зарегулированным условиям [28]. В тоже время, при экологических исследованиях лотических систем возникло направление, включающее исследования мозаичности [29, 30] и неоднородности водных экосистем [31], стохастических и периодических возмущений [31, 32], матрицы естественной среды [33] и иерархии масштабов [34]. Возникли очевидные связи с ландшафтной экологией и ГИС [35]. Оказалось, что более информативно рассматривать проточные системы как речную мозаику, т.е. последовательность «пятен», наполненных ресурсами, проводя тем самым аналогию с работами ландшафтных экологов. Идея, что река состоит из мозаики ресурсных пятен подразумевает, что надо исследовать распределение и структуру этих пятен, т.е. установить баланс энергетического и материального обмена как в продольном (от верховьев до устья), так и поперечном (берега - русло) направлении [36].
Представляется, что основное значение необходимо придавать трем ключевым экологическим понятиям: значимости пространственновременных наблюдений, размеру исследуемой области и продолжительности мониторинговых наблюдений, что необходимо для понимания базовых происходящих процессов в реке. Poole [37] предложил теорию иерархической динамики пятен (Hierarchical Patch Dynamics - HPD), как возможность описывать локально-кусочную прерывистую природу речной сети, усложненную слияниями потоков. То есть, проточные экосистемы рассматриваются как дисконтинуум иерархически вложенных и взаимно со-гласуемых элементов, функциональные связи которых должны быть изучены через масштабы изучаемых ландшафтных компонентов. Предполагается, что способность экосистемы к рассеиванию биоты, а не биоразнообразие является доминирующим фактором формирования структуры речных сообществ, что вовсе не очевидно и, по крайней мере, требует достаточно затратных комплексных исследований специалистов разных профилей.
Мы полагаем, что в любой момент времени вода в сети главного водосбора в конечном итоге течет через главный поток и устье реки. На этом постулате и проводятся обычно расчеты взаимосвязей. Мелкомасштабные процессы в потоках главного водосбора направленно воздействуют на все, что расположено в сети ниже их. Реки - направленные сети [2, 38] и должны быть рассмотрены именно с этих позиций: сегменты ниже по сети не рассматриваются нами как эквиваленты фрагментам, расположенным выше. При оценке экологического состояния малых рек такой связи в исследуемых водотоках мы не установили [16, 39]. Однако, при изучении рек, которые учитывают направленную природу лотических систем, знания значений параметров каждой точки в пространственной динамике, возможно могут помочь в исследованиях связи участков нижнего течения с участками, расположенными выше.
Исследованные нами реки аридных территорий Приэльтонья [6] могут быть исключением, потому что они теряют интенсивность потока по мере увеличения зоны водосбора (например, в связи с зарастаемостью и засушливостью). Однако увеличение расхода воды и ширины рек, возможно, нелинейно связано с изменением абиотических факторов, которые служат естественной средой для множества видов [39]. Мы полагаем, что возможно, необходима такая классификация, которая рассматривает потоки согласно размерам водосборного бассейна в соответствии с особенностями гидробионтов. Кроме того, современная инвентаризация позволила бы абиотическим классификациям, основанным на физических показателях, объединиться с моделями прогноза изменения климата, чтобы предсказать потенциальные изменения в распределении различных лотических сообществ гидробионтов. По-видимому, учитывая вероятность изменения климата и динамическую природу биоразнообразия планктонных и донных сообществ рек, следует ожидать изменения в их классификации и методов оценки экологического состояния разнотипных рек. Проведенные исследования донных сообществ высокоминерализованных рек Приэльтонья показали, что нет четкого разделения биоразнообразия донных таксонов по участкам, а скорее есть динамика видового состава, прерываемая зарегулированными, заросшими, биотопически различными участками рек с разной характеристикой абиотических факторов, в том числе и степенью минерализации [16].
МЕТОДЫ ОЦЕНКИ ЭКОЛОГИЧЕСКОГО СОСТОЯНИЯ РЕК
При обобщении данных гидробиологических исследований 93 средних и малых равнинных рек бассейна Средней и Нижней Волги, включая и реки Приэльтонья, на основе отбора 1392 проб бентоса на 674 станциях рек (1990-2016 гг.), было отмечено в 2, 5 раза большее количество таксонов бентоса в донных сообществах рек лесостепной зоны, в сравнении с реками зоны степей и опустыненных степей (аридная зона бассейна Нижней Волги). Ожидаемо снижение видового богатства макрозообентоса в широтном направлении происходит преимущественно за счет насекомых реофиль-ных таксонов: веснянок, ручейников, поденок и двукрылых (в частности, личинок хирономид из подсемейств Diamesinae и Orthocladiinae). Лим-нофильные формы в реках степной зоны составляют 98% таксономического состава.
Сравнительная оценка числа зарегистрированных в разных ландшафтных зонах видов S выполнялась путем построения кривых разрежения с последующей аппроксимацией данных степенной функцией, позволяющих выполнить сравнительный анализ видового богатства при равном количестве отобранных проб. Проведенный статистический анализ подтверждает постепенное снижение видового богатства макрозообентоса в донных сообществах равнинных рек в направлении от лесостепной зоны к исследован- ным ценозам опустыненных степей (рис. 1).
Для проверки гипотезы о близости видового состава макрозообентоса в реках из различных ландашафтных зон использовался непараметрический дисперсионный анализ Андерсона [40], по результатам выполнения которого приводится рис. 2. Средний квадрат расстояний между центроидами групп существенно превосходит средний квадрат внутригупповых расстояний по F-критерию, т.е. фактор ландшафтной зоны статистически значим с высокой вероятностью, что свидетельствует о постепенном изменении таксономического состава донных сообществ в широтном направлении. Площадь фигуры в пределах каждого обобщающего контура пропорциональна бета-разнообразию групп (т.е. насколько отличаются между собой сообщества рек внутри каждой ландшафтной зоны). Из рисунка 2 видно, что бета-разнообразие максимально для лесостепной зоны.
Оценка разнообразия бентоса рек проведена с выделением уровней γ -, α - и β -составляющих видовое разнообразие донных сообществ (табл. 1). В формировании γ -разнообразия установлено 494 вида макрозообентоса. Выявлено, что отличия в показателях общего разнообразия средних и малых рек невелики, что подтверждает рассчитанный индекс видового разнообразия Шеннона, составляющий для этих групп водотоков 3,9-4,0 бит/экз. соответственно. При экологической диагностике гидроэкосистем целесообразно использовать как характеристики абиотической среды, так и биотические компоненты. Изменение видовой структуры сообществ макрозообентоса речных беспозвоночных определяется как влиянием гидрохими-
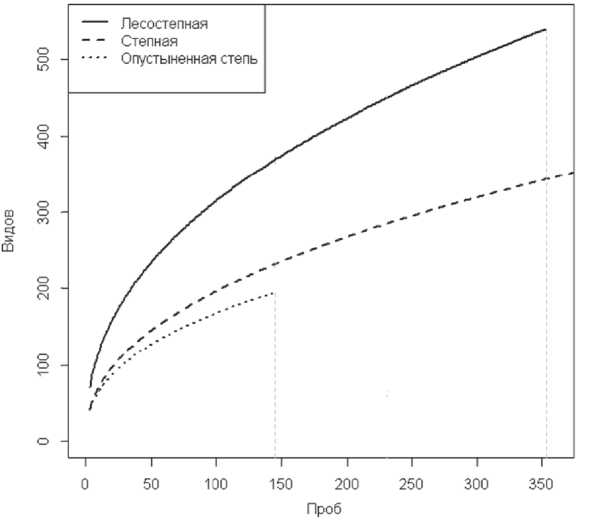
Рис. 1. Фрагмент кривых зависимости видового богатства от числа выполненных проб
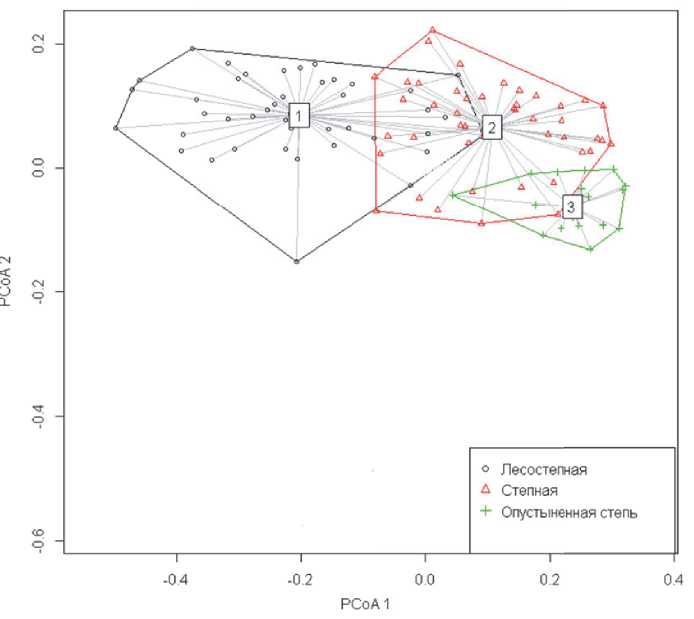
Рис. 2. Положение центроидов таксономического сходства макрозообентоса и оценка бета-разнообразия видового состава рек различных ландшафтных зон
Таблица. 1. Разложение видового разнообразия сообществ макрозообентоса рек лесостепной зоны на α - и β -составляющие; S – число видов; H – индекс Шеннона
В настоящее время используются два основных подхода, позволяющие отделить эффекты химического воздействия от влияния естественных экологических факторов. Первый сводится к применению методик статистического анализа, которые оценивают доли вариации, объясняемые каждым из факторов, в основе которых положе- ны различные многомерные алгоритмы ордина-ции [41]. Другой подход сводится к использованию таких показателей, которые чувствительны, например, к экстремальной изменчивости токсикантов техногенного происхождения.
Следует отметить, что существенным толчком к развитию и совершенствованию методов управления водными ресурсами в странах ЕС стала Европейская Рамочная директива (Water Framework Directive – WFD, Семенченко, 2004) и тесно связанная с ней информационная сеть EuroWaterNet, которая образует статистически стратифицированную систему унифицирован- ных стандартов, приспособленную для решения конкретных задач охраны и управления водными объектами. Согласно директиве введена специальная процедура создания интеркалибровочной сети стран ЕС для оценки биологического состояния водных объектов каждого типа. В связи с этим, до настоящего времени актуальной остается разработка научно-обоснованных комплексных критериев-индикаторов состояния пресноводных экосистем, которая продолжает оставаться сложной задачей [42].
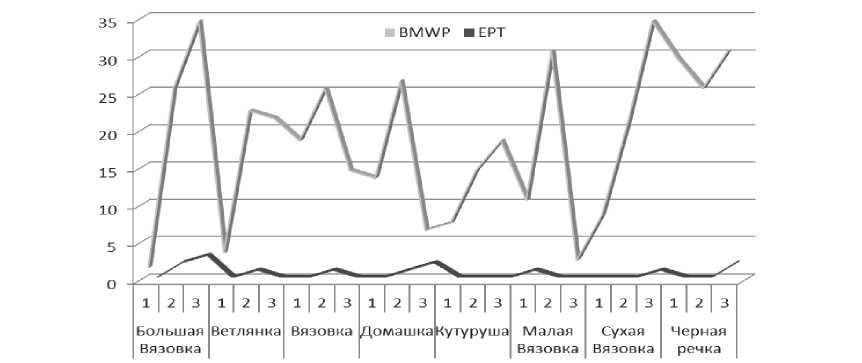
Для оценки экологического состояния лоти-ческих систем применяются различные биотические индексы, основанные на таксономических свойствах сообществ беспозвоночных [43, 44]. Например, величины часто используемого индекса EPT Index , рассчитанные для створов некоторых малых рек бассейна Саратовского водохранилища, в сравнении с его значением для эталонных створов рек, равным 13-15 [45] и другим индексом ВММР ( Biological Monitoring Working Party Index ), показывающим динамичность качества воды, оказались низкими, не превышали 2 балла (рис. 3). Отмечается и вариабельность значений индекса Шеннона на различных участках малых рек, которые изменялись в широких пределах – 0,52-2,42 бит/экз., характеризуя воду рек как «загрязненную» и «грязную».
Ранее, нами, для оценки экологического состояния рек Волжского бассейна были использованы различные интегральные методы и индексы [46, 47]. Несмотря на разработку различных комбинированных систем классификации рек и “интегральных” индексов оценки их экологического состояния, общепринятой методики не существует, в связи со специфическими характеристиками функционирования различных водных объектов в естественных условиях и при воздействии антропогенных факторов. Считаем необходимым привести основные характеристики некоторых методов, представленных в табл. 2. Значения каждого из обобщенных индексов отражают свойства статистических совокупностей экологических объектов [48]. Данные о галотолерантности видов, рассчитанные с использованием индекса SPEARsalinity (см. табл. 2) в донных сообществах [49] могут быть использованы для анализа речных экосистем в условиях происходящих климатических изменений, например, для оценки эффекта увеличения минерализации в лотических системах бассейна нижней Волги Успешное использование нами индекса для оценки изменения видовой структуры донных сообществ в реках с градиентом минерализации на примере рек Приэльтонья, позволил выявить пул индикаторных, в отношении к солености, видов [44].
В практическом использовании применения комплексных критериев необходимо выбрать оптимальный показатель, который обеспечивает минимум ошибки для типологически сходных экологических объектов и, наоборот, иметь значимые отличия для водоемов, находящихся в качественно различном состоянии.
Детальный анализ этапов и алгоритмов получения комплексных показателей, в том числе, методов преобразования исходных данных в сопоставимые шкалы представлен нами ранее [47, 50]. В этой связи, предложенная нами процедура многокритериального экспертного оценивания МЭО,
|
Реки |
Большая Вязовка |
Ветлянка |
Вязовка |
Домашка |
Кутуруша |
Малая Вязовка |
Сухая Вязовка |
Черная речка |
|
H |
1.47-2.42 |
0.63-1.62 |
1.7-2.2 |
1.39-2.17 |
0.99-2.04 |
1.14-2.13 |
0.52-1.73 |
1.75-2.26 |
Рис. 3. Значение индексов качества воды и видового разнообразия некоторых малых рек степной зоны бассейна Саратовского водохранилища
Таблица 2. Основные комплексные показатели оценки качества поверхностных вод, предложенные различными авторами
В качестве примера статистического анализа планктонных и донных сообществ была использована методика экологической диагностики высокоминерализованных рек бассейна нижней Волги для оценки воздействия абиотических факторов на биотические сообщества. [44, 52].
В связи с тем, что планктонные и донные таксоценозы соленых рек остаются мало изученными, то проблема совместного функционирования сообществ и их структурного изменения на сходных биотопах при экстремальных факторах воздействия делает необходимым использование методов многомерного статистического анализ. Например, в соленых реках и озерах нет четкого разграничения планктонных и донных сообществ. Их массовые виды встречаются как на дне, так и в толще воды. Мелководные местообитания соленых рек можно рассматривать как динамические совокупности взаимодействующих между собой макро- и микробиотопов [56]. Они характеризуются разнообразием структурных вариантов сообществ планктона, мейо- и макробентоса, что позволяет сделать анализ взаимодействия локальных сообществ, формирующихся под влиянием солевого градиента и вариабельности среды обитания в пространственных масштабах сопряженных областей (рис. 4). Изучение таких объектов позволяет рассмотреть механизмы перестройки таксономической структуры ценозов, оценить пределы толерантности отдельных видов и выявить потенциал биологических процессов, направленных на адаптацию к лимитирующим факторам, что вызывает интерес в силу специфических особенностей функционирования выделенных комплексов [57].
Для многомерного статистического анализа нами использовались многолетние данные исследований макрозообентоса, мейобентоса и зоопланктона на 15 станциях 5 солёных рек разного уровня минерализации и соответствующий
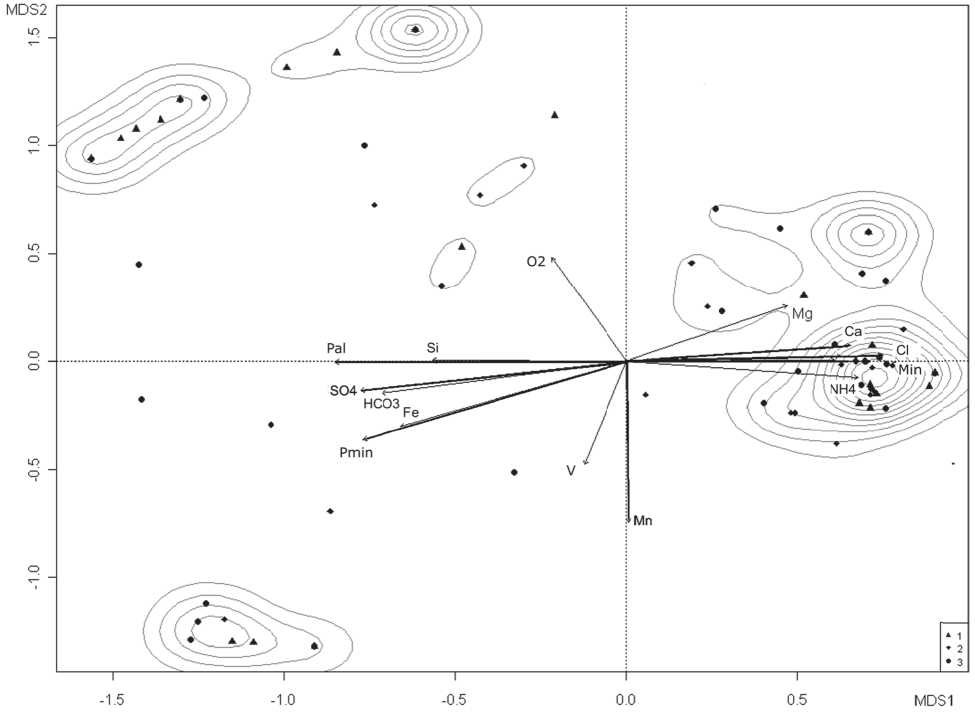
Рис. 4. Ординационная диаграмма распределения видов сообществ макрозообентоса (1), зоопланктона (2) и мейобентоса (3) по осям MDS неметрического шкалирования; стрелками указаны дополнительные оси ведущих факторов среды: содержание кальция (Са), марганца (Mn), магния (Mg), железа (Fe), кремния (Si) аммонийного азота (NH4), минерального (Pmin)и общего фосфора (Pal), растворенного кислорода (O2), сульфатов (SO4), хлоридов (Cl), бикарбонатов (HCO3), взвешенных веществ (V) и общей минерализации (Min).( По: [52])
блок из 19 абиотических показателей (рис. 4). Было установлено, что структуры планктонных и донных сообществ достаточно хорошо коррелируют между собой, следовательно, тесно связаны. С использованием матричного корреляционного анализа Мантеля была подтверждена гипотеза о существовании прямых или косвенных связей между сообществами гидробионтов, обусловленная как их взаимно согласованной реакцией на изменение абиотических факторов, так и межвидовыми взаимодействиями. Особенно сложно выявить реальные отношения в соленых реках и в других лотических системах, где природные условия (уровень минерализации воды, динамичность внешних воздействий.) носят экстремальный характер. Отмечено, что в водоёмах с нестабильными абиотическими условиями решающее значение для популяций приобретают высокая плодовитость организмов, их способность к расселению, неспециализированные трофические связи, способность к анабиозу, поливальтинность амфибиотических насекомых, высокая подвижность личинок, увеличивающие шансы конкурирующих видов на выживание [58].
Для донных сообществ рек Волжского бассейна и высокоминерализованных рек При-эльтонья нами целесообразно использовать информативный метод многомерного анализа (ССА). ССА размещает вместе сходные между собой участки (или реки) вдоль гидрологических градиентов с усреднением обратной величины данных по численности и (или) встречаемости видов (присутствие/отсутствие). Группировки видов вдоль осей ССА связываются с гидрологическими и некоторыми химическими переменными на каждом участке (или реке). Эти параметры абиотических факторов использовались нами, чтобы предсказать конкретную группу видов, выделяемую этим методом (рис. 5).
Подобные подходы (кластерный анализ и классификацию) используют обычно, чтобы
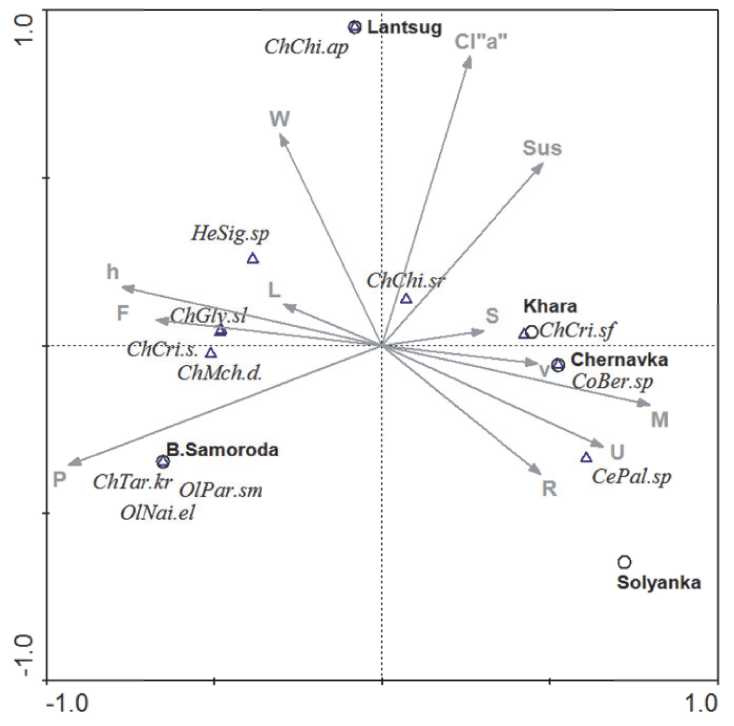
Рис. 5. Ординационная диаграмма ССА взаимосвязи абиотических факторов среды и видового состава таксоценозов бентоса на станциях устьевого участка рек Хара, Солянка, Чернавка, Ланцуг, Б. Саморода (векторы: P – содержание фосфора, F – падение реки, h – глубина рек, L – длина рек, W – ширина рек, Cl“a” – содержание хлорофилла “a”, Sus – взвешенные вещества, S –площадь водосбора, v – скорость течения, M – минерализация, U – уклон реки, R – степень зарастаемости);
Коды видов: OlNai.el - Nais elinguis , OlPar.sm - Paranais simplex , ChTar.kr – Tanytarsus kharaensis , ChCri.s – Cricotopus gr. sylvestris , ChMch.d. – Microchironomus deribae , ChGly.sl – Glyptotendipes salinus , ChChi.ap – Chironomus aprilinus , ChChi.sr – Chironomus salinarius , ChCri.sf – Cricotopus salinophilus , CePal.sp – Palpomyia sp., CoBer.sp – Berosus sp., HeSig.sp – Sigara sp.
найти связанные группы видов вдоль различных экологических градиентов [47, 59, 60]. Так, в представленной ниже ординационной диаграмме (рис. 5) для характеристики бентоцено-зов соленых рек векторы абиотических (гидрологических) и химических факторов образуют три группы: справа- гидрологические факторы среды (степень зарастаемости, скорость течения, взвешенные вещества, уклон реки, минерализация, концентрация Cl“a”, характеризующая продуктивность рек, где высокую продукционную способность демонстрируют популяции массовых хирономид, цератопогонид и жуков, характерных для мезо- и полигалинных рек. Оценить статистическую взаимосвязь каждого из перечисленных факторов можно, спроецировав точку тяжести каждого вида на вектор параметра среды.
Очевидно, что ценоз таксонов бентоса, с доминированием олигохет и эвригалинных хиро-номид (левый квадрант) Microchironomus deribae , Tanytarsus kharaensis и др ., сформированный в устьевом участке мезогалинной реки Большая Саморода развивается в условиях определяющей роли гидрологических факторов и содержания биогенных веществ. Виды Chironomus aprilinus и Ch. salinarius (в верхнем квадранте) образует специфический галофильный ценоз в условиях высокой продуктивности устьевого участка солоноватоводной реки Ланцуг . То есть, установлена значимая связь таксоценозов с продуктивностью гипертрофных водотоков при опосредованном влиянии гидрологических факторов. Индивидуальные гидробиологические показатели ценозов, основанные на видовом богатстве, обилии и разнообразии сообществ, достаточно адекватно оценивают экологические состояние рек в зависимости от комплекса абиотических факторов.
С использованием методов многомерной ординации было выполнено проецирование центров распределения популяционной плотности массовых видов макрозообентоса в пространстве двух главных координат и показана их упорядоченность относительно внешних факторов среды, что позволило оценить, какое место занимает каждый вид на градиенте внешних условий [44]. На основе проведения градиентного анализа распределения различных показателей популяционной плотности (численность, встречаемость, доминирование) для массовых видов были проведены расчеты с использованием пяти возможных форм регрессионных моделей, рекомендуемых при проведении экологических исследований: модели логистической (сигмоидальной) регрессии, гауссовой модели отклика, обобщенных линейной (GLM) и аддитивной (GLM) моделей и модели Хаусмана-Олфа-Фре-ско (HOF). Осуществлена классификация видов бентоса по форме кривой отклика, местополо- жению экологического оптимума минерализации и ширине интервала толерантности [44].
ЗАКЛЮЧЕНИЕ
Несмотря на общее мнение, что донные макробеспозвоночные реагируют на изменения комплекса гидрологических параметров, объединенных в пределах речных экосистем, нет достаточных данных для подтверждений этому тезису данными изменений структуры сообществ и процессов, происходящих в экосистемах крупных масштабов в условиях климатических изменений. Эти сведения необходимы в связи с тем, что антропогенные воздействия продолжают изменять естественную гидрогеоморфологию лотических систем, в большинстве случаев с последствиями для водных сообществ, которые остаются неизвестными.
Гидрологические изменения модулируют изменения как планктоннных, так и донных сообществ через множественные изменения естественной среды (субстрат, структура растительности, гидрохимия), пищевые ресурсы (транспорт и накопление органических, биогенных вещества и их растворимость). В условиях временных изменений климата и увеличивающейся антропогенной нагрузки можно ожидать изменение разных ассоциаций лотических экосистем, предсказание которых возможно только на основе мониторинговых исследований. Проведенный сравнительный анализ различных интегральных индексов, выполненный в результате многолетних исследований лотических систем разного типа и характера антропогенной нагрузки, позволил установить наиболее надежные методы для оценки качества вод малых рек Волжского бассейна. Определенную реакцию на антропогенное воздействие обнаруживают практически все исследованные таксоны сообществ макрозообентоса. Балльная оценка дает возможность показать индикаторную ценность различных характеристик экосистемы реки и выделить наиболее работоспособные индикаторы [44]. Изменения биоразнообразия в экосистеме рек, а также функциональный отклик планктонных и донных сообществ на внешние воздействия находятся под структурирующим влиянием абиотических факторов. Таксономическое разнообразие и количественная динамика планктонных и донных сообществ изменяются на фоне естественной смены климатических условий и градиента отдельных абиотических факторов в условиях, например, эвтрофиро-вания рек. К числу важных факторов надо отнести также значения pH и скорость течения. Эти параметры важны при развитии и смене одного вида другими, особенно в условиях градиента абиотических факторов при исследовании рек аридных территорий. Адаптации биоты связаны прежде всего с гидро-геоморфологическими различиями сравниваемых рек и неоднородностью естественной среды обитания - физической, химической и биотической [61, 62]. Реакция донных беспозвоночных в условиях гидрологических изменений может меняться вдоль речного континуума.
В заключении следует акцентировать внимание на том, что при исследовании экосистемы рек бассейна Нижней Волги мы придерживались гипотезы о существовании пространственно распределенных множеств организмов, адаптированных к обитанию в экстремальных условиях жизни и их взаимодействия с другими ценозами и факторами среды, использовав некоторые методы и подходы для выявления причинно-следственных связей распределения биотических сообществ в условиях воздействия комплекса абиотических факторов среды. Показана роль некоторых методов в экологии сообществ для обработки наблюдаемой информации с целью выявления значимых статистических связей таксономической структуры планктонных и донных сообществ и факторами окружающей среды. Представляется, что возможность формализации наших представлений оценки экологического состояния лотических систем позволит в дальнейшем выявить совокупность нерешенных задач и сформировать представление о путях их решения.
Список литературы Экологическая характеристика лотической системы на примере малых рек Волжского бассейна: методологические подходы исследований
- Доманицкий А.П., Дубровина Р.С., Исаева А.И. Реки и озера Советского союза. Л.: Гидрометеоиздат. 1971. 106 с.
- Малые реки волжского бассейна. Под ред. Н.И.Алексеевского. М.: 1998. 234 с.
- Никитина Н.А., Чалов Р.С. Узлы слияния рек и их морфологические типы // Геоморфология. 1988. №4. С. 64-70.
- Turak E., Linke S. Freshwater conservation planning: an introduction // Freshwater Biology. 2011. V.56. P. 1-5.
- Gordon N.D., Mcmahon T.A., Finlayson B.L., Gippel C.J., Nathan R.J. Stream Hydrology: An Introduction for Ecologists. 2004. P. 233-286.
