Экологические факторы, регулирующие рост макроводорослей в сообществах арктических морей E. V. Shoshina, V. I. Kapkov, O. A. Belenikina

Автор: Шошина Елена Васильевна, Капков Валентин Иванович, Беленикина Ольга Алексеевна
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Океанология
Статья в выпуске: 1-2 т.19, 2016 года.
Бесплатный доступ
Проанализированы особенности макроводорослей в арктических сообществах в связи с периодическими и непериодическими воздействиями экологических факторов. Показано, что существование фитоценозов бентосных водорослей северных морей контролируется в основном первичными периодическими экологическими факторами, выступающими в качестве триггеров, которые определяют направленность вегетативных и генеративных процессов, а также и способствуют появлению адаптационных приспособлений к экстремальным условиям внешней среды. Поэтому периодические воздействия экологических факторов вызывают лишь структурные перестройки в фитоценозах фукусовых благодаря упругой устойчивости популяций водорослей, приобретенной в результате длительного процесса адаптации к условиям северных морей. Непериодические первичные факторы также нарушают соотношение численности за счет у элиминации и угнетения роста определенных возрастных стадий водорослей. Однако и в данном случае благодаря резистентной устойчивости сообщество водорослей способно со временем восстановить структурно-функциональную организацию.
Макроводоросли, периодические и непериодические экологические факторы, арктические сообщества
Короткий адрес: https://sciup.org/14294898
IDR: 14294898 | УДК: 574.587:582.26/.27(268.4) | DOI: 10.21443/1560-9278-2016-1/2-334-344
Ecological factors regulating growth of seaweeds in arctic communities
Features of seaweeds in the Arctic communities in connection with periodic and unperiodic influences of ecological factors have been analyzed. It has been shown that the existence of benthic algae biocenosis of the northern seas is mainly controlled by the primary periodic environmental factors acting as triggers that determine the direction of vegetative and generative processes, as well as contribute to the emergence of adaptive devicesto extreme environmental conditions. Therefore, periodic exposure to environmental factors cause only structural changes in plant communities due to the elastic stability of fucus algae populations acquired as a result of the long process of adaptation to the northern seas conditions. Unperiodic primary factors also violate the ratio of the number by elimination and inhibit growth of certain algae age stages. However thanks to the stability of resistant the algae community can eventually restore its structural and functional organization.
Текст научной статьи Экологические факторы, регулирующие рост макроводорослей в сообществах арктических морей E. V. Shoshina, V. I. Kapkov, O. A. Belenikina
В прибрежных водах гидробионты подвергаются воздействию разнообразных экологических факторов, действующих в совокупности и одновременно, и оказывающих прямое и косвенное влияние на развитие бентосного сообщества. Совместное действие экологических факторов наиболее отчетливо проявляется в северных морях, где температура среды, солнечная радиация и приливно-отливные процессы взаимосвязаны и определяют распределение и доминирование бурых водорослей.
Первичные периодические абиотические факторы выступают в качестве элементов среды, оказывающих непосредственное влияние на метаболизм водорослей бентосного сообщества. При этом могут создаваться как оптимальные, так и неприемлемые условия для роста популяций водорослей в пределах экологической устойчивости вида. Для гидробионтов прибрежных экосистем чрезвычайно важна адаптация к периодическим факторам, особенно – к резким изменениям температуры во время полусуточных приливно-отливных явлений, а также к сезонным изменениям солнечной радиации во время полярного дня и полярной ночи. Именно эти экологические факторы регулируют не только численность особей в популяциях, но и жизненные циклы и сроки размножения водорослей.
Вторичные периодические факторы (шторма, подвижки льда, атмосферные осадки), а также внутривидовые и межвидовые отношения в биоценозе обычно сопряжены с первичными факторами и могут усиливать, ослаблять или нейтрализовывать их влияние на водоросли [1]. Причем особенно важно взаимное влияние факторов или явление синергизма, которое применительно к биологическим системам означает, что абиотические и биотические факторы, когерентно взаимодействуя между собой, действуют не аддитивно, а с проявлением нелинейности [2; 3].
Основными экологическими факторами, определяющими рост и развитие водорослей, являются солнечная радиация, температура воды и концентрация биогенных элементов. Для арктических районов характерны доступность световой энергии в течение короткого летнего периода, длительный темновой период, низкая температура воды на протяжении всего года, низкое содержание биогенов во время летней вегетации.
На встречаемость макроводорослей влияют наличие подходящего твердого субстрата и распреснение воды во время таяния снега и льда.
В данной статье анализируются метаболические особенности макроводорослей в сообществах северных морей в связи с воздействием периодических и непериодических экологических факторов. Работа является продолжением анализа материалов исследований сообществ макроводорослей Баренцева моря в связи с экстремальными условиями обитания [4; 5].
Регион исследования. Материал и методы
Исследования адаптационных приспособлений к экологическим факторам макроводорослей проводили в различных районах Баренцева, Белого и Карского морей. Авторами использованы материалы собственных исследований, основанные на результатах экспедиций в указанные моря. Объектами исследования служили макроводоросли бентосных фитоценозов прибрежной зоны морей в различных гидрологических условиях. При визуальном осмотре учитывали характер грунта, проективное покрытие и изобатическое распределение донных сообществ. Применялась методика гидроботанических разрезов от уреза воды до границы произрастания водорослей. На каждом разрезе обрабатывалось 5–7 станций с необходимым числом мерных площадок. В пробах определяли видовой состав, возраст, численность, биомассу каждого вида и всего сообщества. При анализе результатов исследования опирались на данные, полученные по макроводорослям Баренцева моря: Кольского залива [5; 6], Восточного Мурмана (северная часть высокобореальной подзоны) – губа Дальнезеленецкая (68 ° с. ш.) [7], побережья Земли Франца-Иосифа (арктическая зона) - о. Гукер (80 ° с. ш.) [8; 9]. Использованы также материалы по водорослям Карского (арктическая зона) – мыс Желания, Новая Земля [10] и Белого морей (высокобореальная подзона) – губа Чупа, Порья губа Кандалакшского залива [11; 12].
Результаты и их обсуждение
Наиболее разнообразный видовой состав бентосных сообществ макроводорослей и высокие биомассы характерны для Мурманского побережья. При движении на север и восток Баренцева моря и по направлению к Белому морю с увеличением "ледовитости" района наблюдается снижение видового разнообразия и оскуднение донных фитоценозов как в качественном, так и количественном отношениях. На литорали Мурманского побережья доминируют фукусовые водоросли, в сублиторали густые и многолетние сообщества формируют ламинариевые водоросли. В арктических районах сообщества фукусовых водорослей опускаются в сублитораль и постепенно исчезают в суровых ледовых условиях.
Факторы среды и водоросли высоких широт . Жизнедеятельность растений согласована с сезонной динамикой абиотических и биотических факторов. Ведущим является солнечная радиация, однако существенное влияние оказывают температурный режим, динамика биогенов и интенсивность движения водных масс. В суровых климатических условиях арктических морей эндогенная программа ростовых и репродукционных процессов макроводорослей как неподвижных бентосных фототрофных организмов четко согласована с периодическими сезонными изменениями абиотических факторов среды.
Световые (и ледовые) условия в полярный день и полярную ночь и определяют сжатые сроки вегетации макроводорослей в арктических районах. На побережье Восточного Мурмана (губа Дальнезеленецкая, 68 ° с. ш.) вегетационный период составляет около 9 месяцев, в высоких широтах Земли Франца-Иосифа (о. Гукер, 80 ° с. ш.) — не более 3-4 месяцев. Бентосные макроводоросли отсутствуют в полностью покрытых льдами районах, но произрастают в местах, освобождаемых ото льда на короткое (1–2 месяца) летнее время [13]. Характерной особенностью "светового окна" в высоких широтах являются относительно высокая интенсивность света (при открытой воде), длинный световой день (полярный день), низкое положение солнца над горизонтом, постоянная облачность. Важно и то, что световое окно (июль – сентябрь) приходится на период уменьшения фотопериода (после летнего солнцестояния), что служит сигналом для фототрофных организмов к снижению ростовых процессов и подготовке к зиме.
Вегетативные органы, споры, зиготы, микроскопические стадии водорослей, обитающих в Арктике, способны к переживанию длительного темнового периода. Известно, что одноклеточные гаметофиты ламинарии выживают в течение 6 месяцев темноты и при перенесении в благоприятные световые условия в течение 2 недель (как обычно) образуют спорофиты, тогда как споры теплолюбивых видов погибают [14].
На побережье арктических островов встречаются однолетние и многолетние формы растений. По числу видов преобладают микроскопические водоросли, но основную биомассу и облик сообществ создают многолетние бурые водоросли. Однолетние водоросли зимуют на микроскопической стадии развития – в виде спор или микрослоевищ. Многолетние водоросли сохраняют слоевище (или его часть) на протяжении всего года. В полярную ночь, при отсутствии света и при низкой температуре воды, процессы деструкции у водорослей замедляются. Под мощным снежным и ледовым покровом водоросли находятся в достаточно стабильных условиях, не подвергаясь механическому повреждению во время штормов. В результате многие многолетние виды водорослей имеют относительно крупные размеры слоевища.
В тундре на морском арктическом побережье среди высших растений многие виды сбрасывают листья перед наступлением неблагоприятного сезона, но многочисленны вечнозеленые растения. Вечнозеленые кустарнички и полукустарнички типичны для побережья Кольского полуострова [15]. Подобные явления характерны и для макроводорослей, которые образуют подводные заросли в высоких широтах. Так, у Laminaria solidungula слоевище состоит из нескольких небольших по размеру пластин разных лет образования, а Desmarestia aculeata сбрасывает ветви, и только старые части слоевища сохраняются на протяжении зимы, на них затем весной формируются новые побеги [16]. На побережье Земли Франца-Иосифа отмечены растения Laminaria saccharina с одной-двумя старыми пластинами, тогда как в высокобореальных районах Мурманского побережья у этого вида старая пластина (предыдущего года образования) в течение весны разрушается, поэтому летом встречаются растения только с пластиной текущего года образования. Наличие сформированных вегетативных тканей позволяет многолетним водорослям Арктики (как и высшим растениям в тундре) начинать фотосинтез и вегетацию сразу при наступлении благоприятных световых условий [9].
В связи с особенностями сезонного поступления световой энергии фототрофоным организмам свойственна определенная периодичность метаболических и ростовых процессов. Фотопериод как наиболее точный показатель времени для живых организмов выступает ведущим синхронизирующим фактором жизненных циклов до периода года. Характеризуя сезонную динамику роста слоевища водорослей в условиях северных морей в целом, можно выделить период быстрого роста, когда ростовые процессы являются доминирующими (весна – лето) и период замедленного роста, когда скорость и интенсивность роста снижаются до нулевых значений, а процессы деструкции преобладают над процессами роста (осень – зима).
В полярную ночь в зонах роста ламинарии и других видов (фукусы, пальмария, ульвария) не отмечается каких-либо деградационных изменений фотосинтетического аппарата, но регистрируется уменьшение количества запасных веществ. Показано, что в период полярной ночи интенсивность освещения на Мурманском побережье в течение нескольких дневных часов является достаточной для осуществления у водорослей процесса фотосинтеза [17–19]. Наблюдения в природе и эксперименты при длительном отсутствии света в лаборатории показали, что виды с тонкопластинчатой организацией (ульва, пальмария) сохраняют пластинку при отсутствии освещения в течение 20–30 дней, а виды с плотным и дифференцированным слоевищем (ламинария, фукус) остаются живыми спустя 60–90 дней темноты. У фукуса апикальные части оставались живыми в течение 270 дней темноты. Отмечаются следующие механизмы адаптаций, направленные на сохранение функциональной активности зон роста, обеспечивающих длительное существования при отсутствии освещения: потребление запасных веществ (виды с тонкопластинчатой организацией); использование веществ, образующихся при деструкции таллома (ламинариевые водоросли, наличие дальнего транспорта веществ); потребление растворенных органических веществ (органотрофия фукусовых водорослей) [20; 21].
Водоросли, встречающиеся в Арктике, холодоустойчивы. На Мурманском побережье (Восточный Мурман) среднемесячная температура воды на поверхности изменяется от —1,5 ° С (февраль) до 9 ° С (август). На побережье архипелага Земля Франца-Иосифа крайне низкая температура воды наблюдается на протяжении всего года, составляя —1,8 ° С в зимние месяцы! и всего около -0,7 ° С в июле — августе. Водоросли в морских арктических экосистемах живут при температуре воды! ниже 0 ° С на протяжении всего года, а в течение полярной ночи – около точки замерзания морской воды. Особенно ограничивает рост водорослей низкая температура воды в период летней активной вегетации. Зимой низкая температура воды и образование ледовой кромки встречается не только на арктическом побережье, но и в северных районах бореальной зоны. Так, губа Чупа Кандалакшского залива Белого моря покрыта льдом в декабре – мае, в феврале температура воды! поверхностного слоя составляет около —1 ° С, а летом отмечается хороший прогрев этого слоя. В целом водоросли, обитающие на литорали, более устойчивы к воздействию низких зимних температур (как и высоких летних температур, особенно во время отлива) по сравнению с сублиторальными водорослями. Причем наиболее уязвимы водоросли на ранних стадиях развития и в период активной вегетации слоевища.
Характерной особенностью арктических прибрежных районов является отсутствие литорального пояса макроводорослей, фукусы опускаются в сублитораль, хотя микроскопические виды на литорали присутствуют. Так, на побережье моря Бофорта, где ледовый покров присутствует в течение 7-8 месяцев в году, на литорали широкое распространение имеют водоросли небольшого размера: Calothrix scopulorum , Blidingia minima , Pseudendoclonium spp. , Audouinella purpurum ; эти виды имеют широкое географическое распространение, способны жить в широком диапазоне значений температуры и солености [16]. Отсутствие макроводорослей на литорали связывают с подвижками льда, значительным распреснением, а также с низкой температурой воды и воздуха во время отлива. В арктических районах в холодный период года под толстым слоем льда и снега водоросли защищены от воздействия очень низкой температуры воздуха.
На Мурманском побережье, где наблюдаются правильные полусуточные приливы и отливы (высота прилива в губе Дальнезеленецкой составляет до 4,2 м) литоральные водоросли зимой подвергаются дважды в сутки воздействию отрицательной температуры воздуха, которая может опускаться до —18 °С и держаться на этом уровне в течение нескольких дней. Несмотря на такую низкую температуру, крупные бурые водоросли с плотным слоевищем – фукусовые (Fucus vesiculosus, Ascophyllum nodosum) и красные водоросли с тонкими пластинами – пальмария (Palmaria palmatа) и порфира (Porphyra umbilicalis) не утрачивают своей жизнеспособности. Их слоевища покрываются ледяной коркой, и этот ледяной панцирь защищает водоросли. Слоевище порфиры, которая растет на прибойной скалистой литорали и осушается в сизигийный отлив на длительное время (6–8 часов) может промерзать полностью, однако такое случается нечасто, поскольку зимой наблюдается постоянное волнение моря. Механизм устойчивости баренцевоморских водорослей к замораживанию и размораживанию остается не до конца ясным. Полагают, что жизнеспособность водорослей при низких температурах сохраняется за счет наличия криопротекторных веществ в клетках и отсутствия свободной воды – при природной заморозке водоросли теряют влагу [22].
Насколько биогены могут лимитировать рост водорослей в летний период вегетации в арктических условиях? Сезонная динамика трансформации биогенов в арктических прибрежных районах остается мало исследованной. Предполагается, что общая схема такая же, как и на Мурманском побережье: в осенне-зимний период идет накопление биогенов за счет регенерации и конвективного перемешивания, поступление биогенов также идет при таянии снега и льда, а летом они активно поглощаются автотрофами – фитопланктоном и макроводорослями [7].
Рост водорослей зимой за счет запасных веществ – хорошо известный факт. Ламинария Laminaria saccharina способна накапливать органические вещества в слоевище и использовать их для роста в зимний период, когда солнечной радиации недостаточно для положительного баланса продукционных процессов [7; 19]. Так, в условиях Мурманского побережья рост пластины Laminaria saccharina у многолетних растений начинается в декабре в разгар полярной ночи, когда дыхание преобладает над фотосинтезом. При этом у однолетней ламинарии, не имеющей развитой пластины, а следовательно, и больших запасов питательных веществ, рост начинается позднее, с увеличением уровня солнечной радиации, т. е. в конце января – начале февраля. У Laminaria solidungula , единственного вида из ламинариевых водорослей, имеющего преимущественное распространение в Арктике, формирование пластины начинается в конце полярной ночи, а в течение полярного дня происходит запасание веществ в процессе вегетативного роста [16].
От ледовых условий во многом зависит распределение бентосных сообществ в арктических районах Баренцева моря [13]. Припайные льды при движении удаляют подводную растительность на больших пространствах с любого типа субстрата. При образовании льда во время волнения моря шуга сдирает даже мелкие водоросли. Припайный лед оказывает неблагоприятное воздействие и на субстрат, на котором произрастают водоросли: валуны переворачиваются, а камни "замываются" мягкими грунтами из песка, гравия и ила. Поэтому сублиторальные водоросли в спокойных бухтах часто бывают покрытыми достаточно толстым слоем ила. Скопления айсбергов также оказывают большое влияние на сублиторальную растительность. Перемещаясь течениями, они срезают водоросли с верхушек камней и валунов. Таяние айсбергов и льдов приводит к распреснению поверхностного слоя воды. Кроме того, сходящие ледники несут массу частиц грунта с берега, увеличивая мутность и уменьшая освещенность. Этими факторами объясняется скудность и разреженность зарослей водорослей в местах скопления айсбергов и льдов. Так, на литорали островов архипелага Земля Франца-Иосифа растительность практически отсутствует, на глубине до 2–3 м отчетливо выражена деятельность подвижных припайных льдов, а влияние айсбергов, оставляющих вмятины и борозды на грунте, прослеживается до 25–30 м. При отсутствии существенного действия айсбергов и льдов пояс ламинариевых выражен отчетливо, заросли разреженные, с достаточно разнообразным видовым составом [9].
Распреснение поверхностного слоя воды в результате таяния снега и льда также может оказывать значительное влияние на распределение макроводорослей в арктических экосистемах. Известно, что у побережья Гренландии в бухтах при таянии айсбергов соленость воды у нижней поверхности льдин может снижаться до 3–4 ‰, и эта величина является критической для морских видов. По-видимому, распреснение в сочетании с другими негативно влияющими факторами является одной из причин, по которой фукусы опускаются в сублитораль в арктических районах. Водоросли, обитающие на литорали, в целом лучше адаптированы к значительным колебаниям солености на протяжении суток и года. Многие литоральные водоросли являются эвригалинными. Ламинариевые водоросли, напротив, имеют достаточно узкий диапазон толерантности по отношению к солености, поэтому и обитают в сублиторальной зоне. В прибрежных арктических районах распространение получают виды водорослей, которые обладают достаточно широким диапазоном устойчивости по отношению к солености.
Продуктивность макроводорослей в арктических сообществах. Короткое световое окно и длинная полярная ночь, низкая интенсивность света из-за ледяного и снежного покрова на протяжении большей части года, постоянная облачность летом, низкая температура воды (около нуля) на протяжении всего года, и в летний вегетационный период обусловливают низкую продуктивность автотрофов в арктической зоне. У макроводорослей в арктических экосистемах отмечается уменьшение размеров слоевища, снижение интенсивности фотосинтеза, скорости и интенсивности вегетативного роста, снижение репродуктивного потенциала растений. В сообществах макроводорослей в арктических районах наблюдается падение видового разнообразия, биомассы и плотности поселений. Продуктивность макроводорослей и их сообществ в арктических экосистемах существенно ниже по сравнению с бореальными районами.
Многолетние макроводоросли в Арктике существуют при минимальной продукции органического вещества в течение года. Многолетники включают "грубопластинчатые" формы ( Alaria esculenta , Laminaria solidungula ), "грубокустистые" ( Desmarestia aculeata ) и корковые известковые красные водоросли. Некоторые из них могут иметь достаточно крупное слоевище, что является следствием продолжительной жизни растений в связи со снижением интенсивности процессов деструкции из-за низкой температуры. Отмечается уменьшение размеров слоевища, меньшее число ветвей, уменьшение коркового и центрального слоев клеток у арктических форм водорослей ( Phycodrys rubens , Ptilota serrata , Callophyllis cristata и другие) по сравнению с бореальными формами тех же самых видов [16]. Для ламинарий Земли Франца-Иосифа характерна слабая морфологическая расчлененность пластины: у Laminaria saccharina пластина ровная, со слабой гофрировкой, без булей, а у Laminaria digitata она практически не рассечена на лопасти [9]. На примере ламинариевых установлено, что по размерам и особенно по массе слоевища водоросли архипелага Земли Франца-Иосифа уступают растениям с Мурманского побережья.
У макроводорослей на побережье Земли Франца-Иосифа в силу географических и климатических условий фотосинтез и дыхание соответственно в среднем на 20 и 50 % ниже, чем у аналогичных видов южной части Баренцева моря [7; 23]. Результатом снижения метаболической активности водорослей в Арктике являются меньшие размеры и масса большинства бентосных водорослей. При этом уровень интенсивности фотосинтеза и дыхания ламинариевых в районе Земли Франца-Иосифа по своим абсолютным значениям во время короткого летнего периода может приближаться к величинам, отмечаемым для растений Восточного Мурмана. Однако, как показали измерения, уже в августе в водах Земли Франца-Иосифа, фотосинтез макроводорослей превышает дыхание (табл. 1). Содержание хлорофилла имеет достаточно большой разброс, что обусловлено различиями в морфологической структуре слоевища. Величина продукции и дыхания водорослей, обитающих на побережье архипелага, ниже, чем в южной части Баренцева моря: у бурых водорослей продукция ниже на 26 %, дыхание – на 58 %, у зеленых – на 42 и 83 % соответственно. В первую очередь различия обусловлены температурой воды, разница между районами составляет 7—8 ° С.
Интенсивность фотосинтеза, потребление кислорода и содержание хлорофилла а у макроводорослей на побережье Земли Франца-Иосифа [7]
Таблица 1
|
Макроводоросли |
Фотосинтез валовый, мгС/г сырого вещества в сутки |
Дыхание, мл О 2 /г сырого вещества в сутки |
Содержание хлорофилла а , мкг/г сырого вещества |
|
Бурые |
|||
|
Laminaria saccharina |
0,91 |
0,36 |
185,3 |
|
Pilayella litoralis |
1,17 |
0,44 |
147,8 |
|
Красные |
|||
|
Devaleraea ramentacea |
1,08 |
0,41 |
40,4 |
|
Зеленые |
|||
|
Acrosiphonia sonderi |
0,92 |
0,36 |
188,9 |
Исследования фотосинтеза и дыхания макроводорослей в районе Дальних Зеленцов показали, что в полярную ночь жизнедеятельность исследованных водорослей (бурые – Laminaria saccharina , красные – Palmaria palmata , зеленые – Ulvaria obscura ) характеризуется только дыхательными процессами. Фотосинтез начинает преобладать над потреблением кислорода в середине февраля, когда уровень падающей солнечной радиации составляет 30–50 кал/см 2 (рис.). Высокий уровень фотосинтеза отмечается у водорослей в полярный день: в конце апреля ( Ulvaria obscura , Palmaria palmata ) и начале июля ( Laminaria saccharina ). Снижение фотосинтеза до уровня потребления кислорода у всех исследованных видов наблюдается в конце октября, когда уровень падающей солнечной радиации снижается до 50 кал/см 2 . Динамика продукционных характеристик макроводорослей Баренцева моря имеет сезонную изменчивость, которая связана прежде всего с особенностями поступления солнечной радиации.
Исследования в прибрежных водах архипелага Земля Франца-Иосифа указывают на снижение репродуктивного потенциала макроводорослей в арктических районах по сравнению с Мурманским побережьем [13]. На примере ламинарий, обитающих в водах у побережья архипелага, показано, что растения позднее вступают в стадию размножения, поскольку дольше набирают достаточную массу слоевища, необходимую для начала формирования органов размножения. Ниже процент фертильных растений в соответствующих возрастных группах и в поселении в целом, меньше площадь поверхности пластины, занятая спорангиями. Как следствие, на слоевище ламинарий в арктических районах формируется значительно меньше спор, чем в южных районах Баренцева моря.
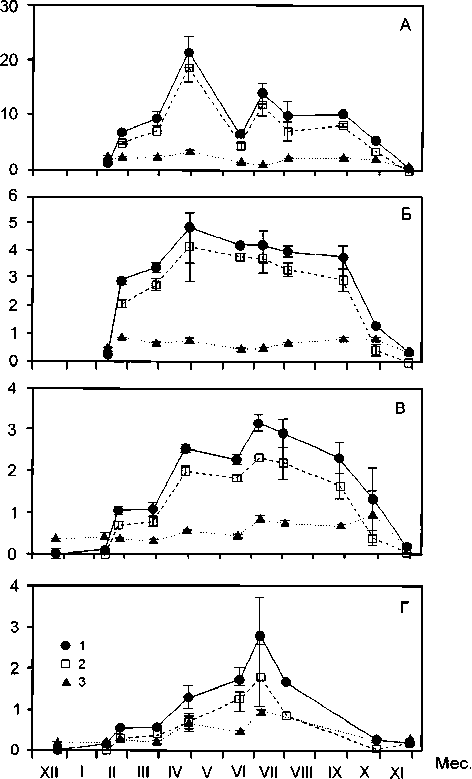
Рис. Годовая динамика валового (1), чистого (2) фотосинтеза и дыхания (3) (млО 2 /г сырого вещества в сутки) водорослей: А – Ulvaria obscura , Б – Palmaria palmata , В–Г – Laminaria saccharina , проростки первого года (В) и взрослые растения (Г) [7]
Считается, что на широте 80 ° с. ш. на поверхность воды поступает 40 % годовой световой энергии от таковой на экваторе и на 25 % меньше, чем в бореальной зоне (60 ° с. ш.) [14; 16]. В Арктике бентосные водоросли встречаются только в местах, освобождаемых ото льда хотя бы на 1–1,5 месяца в летнее время, и отсутствуют в районах с постоянным ледовым покровом. Согласно разным оценкам продуктивность водорослей в Арктике в 2–3 раза ниже, чем в бореальных районах в связи с коротким и холодным вегетационным периодом [24].
Ниже изложенные данные по годовой продукции бентосных талломных водорослей арктических районов свидетельствуют о резком падении ее уровняв в циркумполярной Арктике.
Аляска, море Бофорта (70 ° с. ш., 147 ° з. д.) (табл. 2). Открытая вода наблюдается в июле — сентябре (9 месяцев в году держится ледовый покров), температура воды в это время изменяется от —1 до 4 ° С, соленость 18–25 ‰. Годовой бюджет света (ФАР – фотосинтетически активная радиация) составляет 45–50 моль фотонов на м 2 в год, что около годового минимума, необходимого для произрастания таких крупных водорослей, как ламинария. Количество ФАР изменяется от 5 цмоль ■ м -2 ■ сек -1 (максимум) при ледовом покрове до 200 цмоль ■ м -2 ■ сек -1 при открытой воде. Продукция зарослей Laminaria solidungula оценивается равной 7 гС ■ м -2 в год. В августе на глубине 6—7 м скорость светового потока составляет в августе 100—200 цмоль ■ м -2 ■ сек -1 в дневное время и 20—3 0 цмоль ■ м -2 ■ сек -1 в ночное время; фиксация углерода ламинарией (радиоуглеродный метод) составила 0,09 мкгС/мг в час [25; 26].
Гренландия, Янг Саунд (74 ° с. ш., 20 ° з. д.). Лед держится 9—10 месяцев, свободная ото льда вода — с середины июля по сентябрь. На глубине 10 м водоросли получают потенциально (при отсутствии облачности) 463 моль фотонов ■ м -2 в год. Годовая продукция бурых водорослей составляет 0,1—1,6 гС ■ м -2 в год [27].
Шпицберген, Конгс-фьорд (78 ° с. ш., 20 ° в. д.). Ледовый покров образуется в сентябре и держится до апреля – середины июня. На глубине 10 м водоросли получают потенциально (при отсутствии облачности) 1 092 моль фотонов на м –2 в год [28]. Прибрежные заросли ламинариевых водорослей хорошо развиты.
Канадская Арктика, Иглулик Айленд (69 ° с. ш., 81 ° з. д.). Ледовый покров толщиной около 1,5 м держится на протяжении большей части года; свободная ото льда вода бывает на протяжении 10 недель с августа до середины октября. Водоросли на глубине 8 м получают 590 Е/м 2 света в год; у нижней границы произрастания (20 м) — всего 49 Е/м 2 в год. Годовая продукция бурых водорослей составляет около 20 гС ■ м -2 в год [29].
Архипелаг Земля Франца-Иосифа, о. Хейса, о. Гукера (80 ° с. ш., 52 ° в. д.) (табл. 2). Большую часть года на мелководье в районе о. Хейса держится сплошной припайный лед 1–1,5 м толщиной, море бывает открытым лишь в течение 1–1,5 месяцев в августе – сентябре. Достаточные для фотосинтеза и развития микроскопических и макроскопических водорослей световые условия наблюдаются на протяжении не более 4 месяцев в году (май – август). Продукция водорослей-макрофитов составляет 31–266 мгС/м 2 в сутки; годовая - 133 гС ■ м -2 в год; она соизмерима с продукцией микрофитобентоса и фитопланктона в арктической зоне [23].
Продукционные характеристики сообществ макроводорослей в Арктике и в северной части бореальной зоны
Таблица 2
|
Район, широта, долгота |
Бюджет света |
Продукция макроводорослей |
Источник данных |
|
Земля Франца-Иосифа, о. Гукера, 80 ° с. ш., 52 ° в. д. |
31,2-265,6 мгС ■ м -2 ■ день -1 , 133 гС ■ м -2 ■ год -1 |
[7] |
|
|
Шпицбереген, Конгс-фьорд, 78 ° с. ш., 20 ° в. д. |
1 092 моль ■ м -2 ■ год -1 |
[28] |
|
|
Гренландия, Янг Саунд, 74 ° с. ш., 20 ° з. д. |
463 моль ■ м -2 ■ год -1 (гл. 10 м) |
0,1-1,6 гС ■ м -2 ■ год -1 |
[24; 27] |
|
Мурманское побережье, Дальние Зеленцы, Баренцево море, 69 ° с. ш., 36 ° в. д. |
50-70 кал ■ см -2 ■ день -1 в феврале и 550 ккал ■ см -2 ■ день -1 в июле |
Литораль: 700 гС ■ м -2 ■ год -1 , сублитораль: 2 000 гС ■ м -2 ■ год -1 |
[7] |
|
*Норвегия, район г.Берген, 60 ° с. ш., 0,5 ° в. д. |
240-600 гС ■ м -2 ■ год -1 |
[30; 31] |
|
|
*Германия, о. Гельголанд, 54 ° с. ш. |
400–1 000 моль фотонов ■ м -2 ■ год -1 (4-2 м) |
[14] |
|
|
Аляска, море Бофорта, 70 ° с. ш., 147 ° з. д. |
45-50 моль ■ м -2 ■ год -1 |
7 гС ■ м -2 ■ год -1 |
[25; 26] |
|
Канада, Иглулик Айленд, 69 ° с. ш., 81 ° з. д. |
590 Е ■ м -2 ■ год -1 (8 м), 49 Е ■ м -2 ■ год -1 (20 м) |
20 гС ■ м -2 ■ год -1 |
[29] |
|
*Канада, Новая Шотландия, 43 ° с. ш., 63 ° з. д. |
1 750 гС ■ м -2 ■ год -1 |
[32] |
Примечание. 1 Е = 1 эйнштейн = 1 моль фотонов = 6,02 ■ 10 23 фотонов света.
* Для сравнения приведены данные для бореальных районов.
Таким образом, продукция зарослей бурых водорослей составляет в арктических районах ниже на 1–2 порядка по сравнению с продукцией макроводорослей в северных районах бореальной зоны. Вместе с тем, в различных районах Баренцева моря наблюдаются достаточно высокие значения первичной продукции [7; 33].
Заключение
Существование сложных многоярусных фитоценозов бурых водорослей обеспечивается наличием у популяций адаптационных приспособлений к экологическим факторам при совместном произрастании и увеличивает шансы каждого вида в сообществе на выживание в экстремальных условиях северных морей. Обитая в условиях суточных и сезонных перепадов солнечной радиации, температуры среды, солености и приливно-отливных явлений, бурые водоросли обладают широкой экологической валентностью. Способность бурых водорослей сохранять эндогенные ритмы метаболической активности, синхронизируя их с природными периодическими явлениями, обеспечивает им доминирование в бентосных фитоценозах. Реакции биотической компоненты экосистемы на действие экологических и антропогенного факторов проявляются обычно в форме неспецифического отклика, выражающегося в снижении численности отдельных популяций водорослей, что сопровождается структурной перестройкой фитоценоза. В результате структурных изменений в бентосном сообществе начинают доминировать виды водорослей, обладающие резистентной устойчивостью, способные регулировать численность соотносительно с таковой партнеров по фитоценозу [4].
Первичные периодические абиотические факторы выступают в качестве элементов среды и оказывают непосредственное влияние на продукционные возможности водорослей бентосных сообществ. Вторичные периодические факторы (шторма, подвижки льда, атмосферные осадки) обычно сопряжены с первичными и могут усиливать или ослаблять их влияние. Короткое световое окно и длинная полярная ночь, низкая (около нуля) температура воды в течение всего года, особенно – в летний вегетационный период обусловливают низкую потенциальную продукционную емкость среды для автотрофов в арктической зоне. У макроводорослей в арктических экосистемах отмечаются уменьшение размеров слоевища, снижение интенсивности фотосинтеза, скорости и интенсивности вегетативного роста, снижение репродуктивного потенциала растений, в их сообществах – снижение видового разнообразия, биомассы и плотности поселений. Продуктивность макроводорослей и их сообществ в арктических экосистемах существенно ниже по сравнению с бореальными районами.
Реакции водорослей на экологические факторы меняются в зависимости от сезона, что наиболее четко прослеживается в северных морях. Наибольшее значение в сезонных стадиях активности и покоя принадлежит температуре среды. В зимний период у водорослей существенно падает интенсивность метаболизма, а процессы морфогенеза практически прекращаются, поскольку водоросли переходят преимущественно на эндогенное питание. В клетках водорослей снижается содержание воды, и возрастает концентрация метаболитов, понижающих температуру замерзания цитоплазмы. Зимой во время отлива происходит резкое падение температуры среды, которое сопровождается быстрым охлаждением слоевища водорослей. За столь короткое время центры кристаллизации воды в клетках не успевают сформироваться, стекловидная структура цитоплазмы сохраняет свою целостность, и поэтому резкое падение температуры вызывает у водорослей лишь временный и обратимый анабиоз.
Однако главным регулятором сезонных изменений в фитоценозах макроводорослей являются солнечная радиация и продолжительность светового дня, обусловленные периодическими астрономическими явлениями, которые определяют сезонные циклы роста и размножения. Проявлением адаптации бентосных водорослей к экстремальным условиям арктических морей, обеспечивающей их доминирование в донных фитоценозах, является разобщение во времени и пространстве слоевища вегетативного и генеративного процессов.
Таким образом, существование фитоценозов бентосных водорослей северных морей контролируется в основном первичными периодическими экологическими факторами, выступающими в качестве триггеров, которые определяют направленность вегетативных и генеративных процессов, а также способствуют появлению адаптационных приспособлений к экстремальным условиям внешней среды. Поэтому периодические воздействия экологических факторов вызывают структурные перестройки в фитоценозах фукусовых благодаря упругой устойчивости популяций водорослей, приобретенных в результате длительного процесса адаптации к условиям северных морей. Непериодические первичные факторы также нарушают соотношение численности за счет элиминации и угнетения роста определенных возрастных стадий водорослей. Однако и в этом случае благодаря резистентной устойчивости сообщество водорослей способно со временем восстановить структурнофункциональную организацию.
Список литературы Экологические факторы, регулирующие рост макроводорослей в сообществах арктических морей E. V. Shoshina, V. I. Kapkov, O. A. Belenikina
- Kapkov V. I., Belenikina O. A., Fedorov V. D. Effect of heavy metals on marine phytoplankton//Moscow Univ. Biol. Sci. Bulletin. Allerton Press (NY, USA). 2011. V. 66, N 1. P. 32-36.
- Розенберг Г. С., Рянский Ф. Н. Теоретическая и прикладная экология. Нижневартовск: Нижневартов. пед. ин-т, 2005. 292 с.
- Твердислов В. А., Сидорова А. Э., Яковенко Л. В. Биофизическая экология. М.: КРАСАНД, 2012. 544 с.
- Макроводоросли в системе биологического мониторинга морских прибрежных экосистем/Е. В. Шошина, Г. М. Воскобойников, М. В. Макаров, С. Е. Завалко, В. И. Капков//Вестник МГТУ. 2012. Т. 15, № 4. С. 851-857.
- Шошина Е. В., Капков В. И. Экологические особенности промысловых фукусовых водорослей Мурманского побережья Баренцева моря//Вестник МГТУ. 2014. Т. 17, № 1. С. 180-189.
- Канищева О. В., Шошина Е. В. Возрастная структура поселений Fucus vesiculosus на литорали Кольского залива//Вестник Удмурт. ун-та. Сер. Биология. 2015. Вып. 1. С. 40-47.
- Кузнецов Л. Л., Шошина Е. В. Фитоценозы Баренцева моря (физиологические и структурные характеристики). Апатиты: Изд-во КНЦ РАН, 2003. 308 с.
- Виноградова К. Л., Шошина Е. В. Водоросли//Окружающая среда и экосистемы Земли Франца Иосифа (архипелаг и шельф). Апатиты, 1994. С. 109-116.
- Шошина Е. В., Макаров В. Н., Макаров М. В. Биологические особенности ламинариевых Земли Франца-Иосифа//Биология моря. 1997. № 5. С. 286-292.
- Шошина Е. В., Анисимова Н. А. Макроводоросли из района бухты Ледяная Гавань (Новая Земля, о. Северный, Карское море)//Вестник МГТУ. 2013. Т. 16, № 3. С. 530-535.
- Шошина Е. В. Макрофиты//Исследование фауны морей. 2012. Т. 69 (77). С. 132-149.
- Шкляревич Г. А., Шошина Е. В. Макроводоросли эстуарной зоны (на примере Порьей губы Белого моря)//Труды Карел. науч. центра РАН. 2015. № 6. С. 1-7.
- Голиков А. Н., Аверинцев В. Г. Биоценозы верхних отделов шельфа архипелага Земля Франца-Иосифа//Исследования фауны морей. 1977. Вып. 14 (22). С. 5-54.
- Lüning K. Seaweeds. Their environment, biogeography and ecophysiology. New York: John Wiley, 1990. 527 р.
- Василевская Н. В. Поливариантность онтогенетических процессов растений в условиях высоких широт. Мурманск: Изд-во МГПУ, 2007. 231 с.
- Wilce R. T., Dunton K. H. The Boulder Patch (North Alaska, Beaufort Sea) and its benthic algal flora//Arctic. 2014. N 67 (1). Р. 43-56.
- Быков О. Д. Фотосинтез и темновая фиксация СО2 литоральных макроводорослей Баренцева моря в условиях полярной ночи//Ботан. журн. 2003. № 12. С. 68-73.
- Тропин И. В., Макаров М. В. Фотосинтетический аппарат представителей Fucales (Phaeophyta) Баренцева моря после полярной ночи//Альгология. 2004. Т. 4, № 4. С. 393-404.
- Макаров М. В., Рыжик И. В., Воскобойников Г. М. Механизмы существования бурых водорослей в период полярной ночи: функциональная дифференциация и гетеротрофия//Современные проблемы альгологии. Ростов н/Д, 2008. С. 225-227.
- Воскобойников Г. М., Макаров М. В., Рыжик И. В. Изменения в составе фотосинтетических пигментов и структуре клеток Fucus vesiculosus L. и F. serratus L. Баренцева моря при длительном нахождении в темноте//Биология моря. 2006. Т. 32, № 1. С. 26-33.
- Дифференциация пластины Laminaria saccharina (L) Lamour. как приспособление к длительному отсутствию освещения/М. В. Макаров, И. В. Рыжик, Г. М. Воскобойников, Г. Г. Матишов//Доклады РАН. Общая биология. 2006. Т. 409, № 2. С. 1-2.
- Макаров М. В., Воскобойников Г. М. Влияние освещения и температуры на макроводоросли Баренцева моря//Труды КНЦ РАН. 2013. № 1 (14). С. 95-111.
- Кузнецов Л. Л., Макаревич П. Р., Макаров М. В. Структурно-продукционные показатели морских фитоценозов//Среда обитания и экосистемы Земли Франца-Иосифа (архипелаг и шельф). Апатиты, 1994. С. 89-94.
- Biomass, photosynthesis and growth of Laminaria saccharina in a high-Arctic fjord, NE Greenland/J. Borum, M. F. Pedersen, D. Krause-Jensen, P. B. Christensen, K. Nielsen//Mar. Biol. 2002. V. 141. P. 11-19.
- Dunton K. H., Riemnitz E., Schonberg S. An Arctic kelp community in the Alaskan Beaufort Sea//Arctic. 1982. N 35. P. 465-484.
- Dunton K. H. Growth and production in Laminaria solidungula: relation to continuous underwater light levels in Alaskan High Arctic//Mar. Biol. 1990. V. 106. P. 297-304.
- Benthic primary production in Young Sound, Northeast Greenland/D. Krause-Jensen, M. Kühl, P. B. Christensen, J. Borum//Bioscience. 2007. N 58. P. 160-173.
- Light regime in an Arctic fjord: A study related to stratospheric ozone depletion as a basis for determination of UV effects on algal growth/D. Hanelt, H. Tüg, K. Bischof, C. Gross, H. Lippert, T. Sawall, U. Karsten, C. Wiencke//Mar. Biol. 2001. V. 138. P. 649-658.
- Chapman A. R. O., Lindley J. E. Seasonal growth of Laminaria solidungula in the Canadian high Arctic in relation to irradiance and dissolved nutrient concentrations//Mar. Biol. 1980. V. 57. P. 1-5.
- Sjotun K., Fredriksen S., Lein T.E., Rueness J., Sivertsen K. Population studies of Laminaria hyperborea from its northern range of distribution in Norway//Hydrobiologia. 1993. V. 260/261. P. 215-221.
- Ecological studies of the kelp Laminaria hyperborea (Gunnerus) Foslie in Norway/K. Sjøtun, S. Fredriksen, J. Rueness, T. E. Lein//Ecology of Fjords and Coastal Waters. Amsterdam, 1995. P. 525-536.
- Mann K. H. Ecological energetics of seaweed zone in a marine bay on the Atlantic coast of Canada. II. Productivity of the seaweeds//Mar. Biol. 1972. V. 14. P. 199-209.
- Макаревич П. Р. Первичная продукция Баренцева моря//Вестник МГТУ. 2012. Т. 15, № 4. С. 786-793.
