Экологические ниши, их современная интерпретация и перспективы моделирования

Автор: Шитиков В.К., Розенберг Г.С., Зинченко Т.Д.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Наземные экосистемы
Статья в выпуске: 4 т.30, 2021 года.
Бесплатный доступ
Обсуждается развитие теоретических представлений об экологических нишах в условиях интенсивного накопления биосферных данных. Уточняются классификация и формальные определения фундаментальных и реализованных ниш. Сделан обзор методов и программных средств моделирования экологических ниш и распределений видов.
Типы экологических ниш, моделирование распределений видов, статистические пакеты r, метод главных компонент
Короткий адрес: https://sciup.org/148323683
IDR: 148323683 | УДК: 598.12(470.56) | DOI: 10.24412/2073-1035-2021-10427
Ecological niches, their modern interpretation and modeling prospects
The development of theoretical ideas about ecological niches in conditions of intensive accumulation of biosphere data is discussed. The classification and formal definitions of fundamental and realized niches are being clarified. A review of methods and software tools for modeling ecological niches and distributions of species is made.
Текст научной статьи Экологические ниши, их современная интерпретация и перспективы моделирования
Интенсивное развитие ГИС-технологий и дистанционных систем мониторинга, а также постоянно возрастающий объем общедоступных баз данных, обобщающих характеристики природных и антропогенных ландшафтов, динамику климатических изменений и широкий набор иных факторы окружающей среды, открывают новые возможности для изучения механизмов функционирования сообществ и их связи с условиями обитания популяций. За последние десятилетия мощным инструментом (макро)-экологических и биогеографических исследований стало моделирование распределения видов SDM ( Species Distribution Models – Franklin, 2009; Guisan et al., 2017) и экологических ниш ENM ( Environmental Niche Models – Peterson et al., 2011; Melo-Merino et al., 2020).
При этом, однако, обнаружилось, что для тысяч видов легко можно рассчитать некие абстрактные объекты, формально называемые "нишами", но имеющие прямое отношение к оценке географических районов распространения. Дискуссии о том, что именно включает "моделирование экологической ниши", оказались запутанными из-за отсутствия ясности в использовании терминов и трактовке определений (So-berón, 2007; Peterson, Soberón, 2012). Для однозначной интерпретации классических теоретиче-
моделирование распределений видов, статистических концепций, разработанных в середине прошлого века, требуется их переосмысление и четкое изложение основных понятий, что и является целью нашей публикации. Мы также делаем обзор основных методов, алгоритмов и программных средств, широко используемых для моделирования ниш и ареалов.
Экологические ниши Гринелла и Элтона
Закономерности распределения популяций по изучаемой территории определяются тремя важнейшими группами факторов (Brown et al. 1996; Gaston, 2003; Guisan, Thuiller 2005): ( а ) пространственный градиент условий окружающей среды в диапазоне факторов, благоприятных для появления, выживания и размножения видов (в основном, в физиологическом смысле); ( б ) факторы биотической среды, определяющие идентичность и обилие "посредников" (опылителей или переносчиков семян), хищников, паразитов и конкурентов, а также доступность и динамика потребления ресурсов; и ( в ) способность видов к воспроизводству и расселению за счет собственных перемещений или с помощью внешних агентов.
Интерес к описанию, пониманию и прогнозированию географического и экологического распределения видов имеет давнюю историю (Wallace, 1860; Grinnell, 1917, 1924; Elton, 1927). Дж. Хатчинсон (Hutchinson, 1957, 1978), проанализировав эти работы, выделил два класса экологических ниш.



Гринелл Джозеф (Joseph Grinnell; 1877-1939) – американский зоолог, орнитолог
Элтон Чарльз (Charles Sutherland Elton; 1900-1991) – британский зоолог, эколог
Хатчинсон Джордж
(George Evelyn Hutchinson; 1903-1991) – британский, американский эколог, гидробиолог, лимнолог
Ниши Гринелла формируются в пространстве принципиально неинтерактивных переменных, отражающих условия окружающей среды, для которых конкуренция не имеет значения, но необходимых для понимания крупномасштабных экологических и географических закономерностей (Grinnell, 1917; Whittaker et al., 1973). Такие переменные были названы "сценопоэтическими" ( scenoproetic ), от греческих корней "постановка сцены". Другой класс – ниши Элтона сосредоточены на учете всех типов биотических взаимодействий, которые оцениваются главным образом в локальных масштабах (Elton, 1927; MacArthur, 1969). Сюда включаются также интерактивные переменные среды, названные Хатчинсоном биономическими ( bionomic ), которые связаны с потребляемыми ресурсами, а население изучаемых сообществ может влиять на них и конкурировать за них. Различие между сценопоэтическими и биономическими переменными достаточно искусственно, поскольку некоторые из них могут в зависимости от масштаба играть важную роль при описании ниш обоих классов (например, солнечное излучение является и функцией географической широты, и фактором освещенности лесной подстилки).
Основополагающие идеи фундаментальных и реализованных ниш Хатчинсона отражают общие концепции n -мерного гиперпространства переменных и могут быть применены к нишам обоих классов, однако детали значительно меняются, поскольку проблемы измерения сценопоэтических и биономических осей совершенно разные. Отметим, что когда Хатчинсон (1957) определил нишу как совокупность всех "...состояний окружающей среды, которые позволили бы видам... существовать бесконечно", он не делал отличий между биотическими и сценопоэтическими переменными, хотя позже осознал их важность (Hutchinson, 1978).
Концепция фундаментальных и реализованных ниш
Предположим, что область исследований G разделена сеткой географических координат, ячейки которых имеют разрешение, соизмеримое с масштабом гринеллевских переменных (например, > 10 3 км 2 ). Прежде, чем оценивать функции распределения анализируемого вида, необходимо принять соглашения об условиях абстрагирования каждой ячейки как "присутствие вида" с учетом масштаба и темпов демографических процессов. Этими условиями могут быть, например: "плотность населения превышает некоторый заданный порог" или "уровень рождаемости особей превышает уровень смертности". Часто оперируют неотрицательным параметром внутреннего темпа роста r ( intrinsic growth rate ) популяции, оцениваемом в предположении, что биотические факторы среды не налагают никаких ограничений.
Если для каждого вектора e j , являющегося j -й произвольной комбинацией переменных окружающей среды, известно значение внутреннего темпа роста r , то фундаментальную нишу Гри-нелла можно строго определить как N F = { e j | r ( e j ) > 0} , т.е. « набор всех возможных векторов e n-мерного пространства переменных среды, для которых внутренний темп роста численности вида положителен и/или популяционная плотность превышает заданный порог » (So-berón, 2007, р. 1118). Отметим, что нишу как среду обитания следует отличать от ниши как функции (Whittaker et al., 1973). Если рассматривать конкретное географическое пространство G , то можно найти все ячейки в G со значениями векторов переменных, принадлежащих N F :
J F = {j ∈ G | r ( e j ) > 0}.
Этот набор ячеек, привязанный к сетке географических координат, определяется как фундаментальная область распространения вида
( fundamental area of distribution of a species ). На основе N F можно вычислить площадь J F , но теоретически допустима и обратная операция: если известна J F , то из ее ячеек можно извлечь комбинации векторов переменных среды для построения N F . Однако эта операция справедлива только в предположении, что J F является потенциальной областью распространения, и такие факторы, как перемещения особей и взаимодействия между видами, игнорируются.
Дж. Хатчинсон определил фундаментальную нишу, как объем n -мерного гиперпространства всех внутренних и внешних факторов, необходимых для выживания каждого вида в условиях отсутствия других видов, а реализованную нишу – как часть фундаментальной ниши, в которой присутствие вида зафиксировано в условиях лимитированных ресурсов и межвидовых взаимодействий. Однако, даже в случае ниши Гри-нелла, сведение N F к реализованной нише является сложной проблемой, начиная с того факта, что по определению не может существовать конкуренции за переменные среды. Следовательно, уменьшение площади N F может происходить только за счет процессов в элтоновской нише. Дж. Чейз и М. Лейболд (Chase, Leibold, 2003) подробно рассмотрели, как фундаментальная ниша сокращается в ходе конкуренции за ресурсы при переходе к реализованной нише, которая также может быть выражена в пространстве ресурсных переменных. Они уточнили основные определения ниш с учетом того, что вид потенциально может существовать только там, где плотность ресурсов превышает минимальные потребности вида, чтобы поддерживать положительный (или нейтральный) чистый рост населения.
Таким образом, общий подход к моделированию экологических ниш основывается на системе уравнений популяционной динамики, и использует разложение суммарной скорости роста численности вида в заданной точке пространства (или каждой j -й ячейки на сетке) на три положительных или отрицательных компонента (So-berón, 2007):
-
- внутренний темп роста r ( e j ), не зависящий от интенсивности биотических взаимодействий и определяющийся (при наличии необходимых ресурсов) простым вектором крупномасштабных (> 10 3 км 2 ) факторов среды гринеллев-ских ниш, таких как средняя температура, осадки, солнечная радиация, рельеф местности и т. д.;
-
- изменения темпов роста с ( R j ), обусловленные биотической средой и конкуренцией за ресурсы, в которых доминируют процессы ниши Элтона с пространственным разрешением, соиз-
- меримым с перемещениями индивидов (до 10 км2);
-
- компонент, связанный со структурой мета-популяции и динамикой дрейфа, заданный матрицей переходов T , которая определяет вероятности того, что в любой ячейке в течение определенного периода времени произойдет прием и отправка мигрантов.
Эта упрощенная теоретическая схема позволяет определять ниши с точки зрения наборов параметров, а области распространения вида – с точки зрения наборов ячеек с фактическими или потенциальными демографическими свойствами популяций. Однако практическое моделирование ниш в зависимости от перечисленных трех групп факторов является чрезвычайно сложной задачей. Первая проблема связана с различным масштабом ниш. Ниша Элтона имеет мелкоячеистую экологическую структуру и, в результате хищничества или конкуренции за ресурсы, многие местные популяции могут быть сокращены или даже уничтожены. Однако в географических масштабах, достаточно больших, чтобы нивелировать некоторую пространственную неоднородность, конкурентные виды могут сосуществовать в течение длительных периодов времени, и это никак не будет сказываться на размерах ниши Гринелла.
Вторая проблема заключается в том, что на практике эффекты взаимодействий и другие биономические параметры модели элтоновской ниши трудно предсказуемы по данным полевых наблюдений или экспериментов. Матрица R j не может быть определена просто как набор чисел, поскольку для оценки ее компонентов требуются гипотезы о конкретных формах и моделях взаимодействий (Chase, Leibold, 2003).
Модели ниш и распределения видов: различия и практическая реализация
Моделирование ниш ENM и распределений видов SDM стало основной методологией количественной оценки связи структуры экологических сообществ с факторами окружающей среды. По состоянию на 18.02.2008 г. поиск в ISI Web of Science с использованием термов ‘‘species distribution models’’ или ‘‘niche models’’ или ‘‘habitat models’’ обнаружил 21 973 статей, 15% из которых посвящены биологии морских и внутренних вод (Thuiller et al., 2009). К настоящему времени этот объем, вероятнее всего, удвоился, однако в подавляющем большинстве построение моделей основывалось на легкодоступных географических и климатических переменных или факторах абиотической среды, которые имеют свою собственную внутреннюю динамику и не зависят от встречаемости или численности рассматривае- мых видов. Наборы переменных, которые динамически связаны с численностью популяций, такие как лимитирующие пищевые ресурсы, влияние конкурентов или хищников и межвидовые взаимодействия в практике построения моделей почти не встречаются. Сместились и терминологические акценты: целью моделирования является, в основном, «географическое воплощение потенциальной ниши (“potential niche”) как пересечение фундаментальной экологической ниши, физиологически определяющей диапазоны переменных среды, в которых вид может выжить, с набором факторов среды, которые фактически существуют на рассматриваемом реальном ландшафте» (Barve et al., 2011).
Постоянно разрабатываются новые методы и алгоритмы, позволяющие сделать моделирование как экологических ниш, так и распределений видов более удобным, полным, интерпретируемым и адекватным. Ранние модели SDM были ориентированы исключительно на географические измерения, а, например, в случае пространственной эпидемиологии, вообще могли не иметь привязки к условиям окружающей среды. Однако многие современные методы (Guisan et al., 2017; Ovaskainen, Abrego, 2020; Шитиков и др., 2021а) в качестве основной цели прогнозирования ставят оценку степени различий между фактическим ареалом вида и его потенциальным распределением на основе статистического анализа комплекса внешних переменных.
Некоторые исследователи видят серьезные нарушения концептуальной строгости, когда во многих статьях описывается процесс моделирования распределения видов SDM, тогда как на самом деле рассматриваются модели ENM, связанные с экологическими нишами, и наоборот. Они считают, что термин ENM следует использовать, если основное внимание уделяется оценке любого потенциального распределения в гипотетических условиях без привлечения "существующих" проявлений. Действительно, фундаментальная ниша Гринелла N F , которая представляет собой "конструкцию физиологических реакций" (т.е. любой произвольный набор условий e j ), может выходить далеко за рамки географии изучаемой области, и даже находиться вне реального мира. С другой стороны, использование термина SDM должно включать шаги по преобразованию ареалов, оцененных как потенциальные, в фактическую реконструкцию распределения видов (Peterson, Soberón, 2012, р. 4).
Однако современный процесс разработки SDM включает в себя, по крайней мере, два перехода между пространствами: (а) начальные стадии моделирования выполняются в географическом пространстве; (б) сама модель формиру- ется в пространстве факторов окружающей среды; (в) результаты моделирования снова, как правило, визуализируются в географическом пространстве. Таким образом, подлинное "моделирование распределения видов" включает в себя как этап выделения экологической ниши, так и этапы оценки способности вида к распространению или его потенциал колонизации.
Для оценки географических ареалов видов получил широкое распространение метод максимальной энтропии MaxEnt (Phillips et al., 2006; Лисовский и др., 2020; Шитиков и др., 2021b), которая с байесовской точки зрения из всех возможных распределений вероятностей наилучшим образом представляет моделируемые данные. Итогом работы MaxEnt является обобщенная экспоненциальная функция, аргументами которой являются частные функции отдельных предикторов (линейные, квадратичные, множественные и др.) с настроенными коэффициентами λ, оценивающими вклад соответствующего экологического фактора, т.е. по сути, модель экологической ниши (Petrosyan et al., 2020). С другой стороны, связь между различными факторами в географическом и экологическом пространствах исследуют с помощью моделей ENM, построенных для совокупности видов, называемых искусственными или "виртуальными", с одинаковым и известным откликом по отношению к внешним воздействиям (Hirzel et al., 2001). Таким образом, различия моделей ENM и SDM в значительной мере нивелируются.
Поскольку количество доступных алгоритмов и программных средств SDM-ENM велико и неуклонно растет, это затрудняет выбор наиболее подходящей методологии. Расхождения между различными технологиями могут быть очень большими, но, тем не менее, до сих пор в полной мере отсутствует не только единая теория, но и конкретные практические рекомендации. Это обусловлено как объективно существующим многообразием изучаемых экологических сообществ, природно-климатических зон, жизненных форм и техник проведения наблюдений, так и большим арсеналом разработанных методов компьютерной обработки и верификации моделей, выбор которых в значительной мере определяется субъективными вкусами исследователей. В частности, развернутый анализ результатов использования 33 моделей SDM на сообществах птиц, бабочек, деревьев и травянистой растительности (Norberg, 2019) обозначил лишь самые общие ориентиры.
Компьютерные решения по построению моделей SDM-ENM реализуются, в основном, в форме функций пакетов статистической среды R (R Development Core Team): SDMvspecies и virtualspecies для различных схем "виртуальных" видов, dismo, ecospat, modleR для калибровки моделей MaxEnt и их сравнительных исследований, ENMTML, ENMTools, NicheToolBox, BIOMOD2, ENMeval, spThin, kuenm, SDMTools, sdm и многие другие пакеты для подготовки исходных данных, построения, прогнозирования, графической иллюстрации, тестирования и оценки моделей экологических ниш с использованием различных алгоритмов. Альтернативой R может быть кросс-платформенное интерактивное приложение NicheA (Niche Analyst), выпущенное по публичной лицензии GNU и написанное на языке Java (Qiao et al., 2016).
Методы выделения пространства экологических ниш
Выше отмечено, что в подавляющем большинстве случаев под термином "экологическая ниша" понимается простейшая фундаментальная ниша Гринелла. Один из распространенных подходов к моделированию пространства ниш заключается в оптимальном проецировании многомерной коррелированной совокупности исходных абиотических факторов на ортогональные оси пространства малой размерности. Для этого строят ординационные диаграммы в координатах первых двух главных компонент (PCA1 и PCA2) и калибруют их с использованием характера вариации переменных среды в пределах изучаемой области. На сетку ячеек, ограниченную минимальными и максимальными значениями главных компонент и определяющую многомерную экологическую изменчивость территории, нано- сят точки встречаемости анализируемых видов. Модель потенциальной экологической ниши рассматривается как некоторая область в пространстве латентных переменных, в которой плотность вероятности встречаемости вида соответствует заданным ограничениям. Эти условия связывают с индексом пригодности среды обитания (Z е [0,1] - environmental suitability).
Для определения области экологической пригодности условий среды распределение проекций точек встречаемости видов на каждую из осей PCA 1 и PCA 2 аппроксимируют гауссовыми кривыми, после чего плотности вероятности обобщают для получения итоговых значений (Leroy at al., 2016). Экологическая ниша вида для такой простейшей функции – эллипс на ордина-ционной диаграмме с заданным уровнем доверительной вероятности и соответствующими диапазонами значений главных компонент.
Другой способ задать границы ниши, используемый в пакете ecospat , заключается в преобразовании бинарных переменных присут-ствия/отсутствия в относительные плотности двумерного распределения путем сглаживания ядерными ( kernel ) функциями на сетке 100×100 с координатами PCA 1 и PCA 2 (Broennimann et al., 2012; Di Cola et al., 2017). После калибровки поверхности отклика Z в диапазоне [0, 1] потенциальная экологическая ниша в такой интерпретации – некоторая нерегулярная область в пространстве двух главных компонент со значениями пригодности среды обитания z ij > 0 – см. рис. 1.
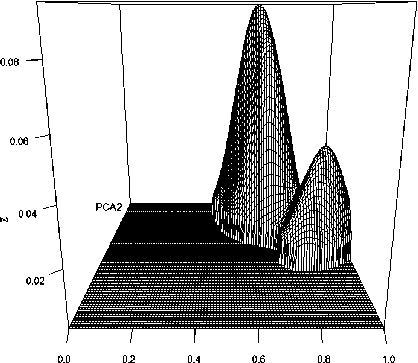
РСА1
Рис. 1. Поверхность экологической ниши Tubifex tubifex , заданная индексом пригодности среды обитания Z в координатах главных компонент PCA 1 и PCA 2 .
Fig. 1. The surface of the ecological niche of Tubifex tubifex , defined by the habitat suitability index Z in the coordinates of the main components PCA 1 and PCA 2 .
Обычно ставится задача оценить степень перекрытия двух ниш, принадлежащих разным таксономическим группам, для чего строят ор-динационные диаграммы их попарного пересечения. Для количественной оценки используют такие индексы, как D Шёнера ( Schoener ) и I Хел-лингера ( Hellinger ), которые отражают долю совмещения ниш в заданном пространстве (Warren et al., 2008) и колеблются между 0 (отсутствие перекрытия) и 1 (полное перекрытие):
D - 1 - o^ Z 1 i - z 2 i l) ;
I ( 0.5 0.5 2 I
I - 1 0.5^Xl ij ( Z 1 ii z 2ij ) J •
На рис. 2 представлен пример сравнения экологических ниш трех видов макрозообентоса: личинок комара-звонца Prodiamesa olivacea , малощетинковых червей Tubifex tubifex и жуков-падальщиков Berosus sp. Диаграммы построены с использованием пакета ecospat статистической среды R и на основе данных многолетнего комплексного гидробиологического мониторинга 92 малых и средних рек Среднего и Нижнего Поволжья (Зинченко, 2011; Шитиков, 2020; Шитиков и др., 2021а, b).
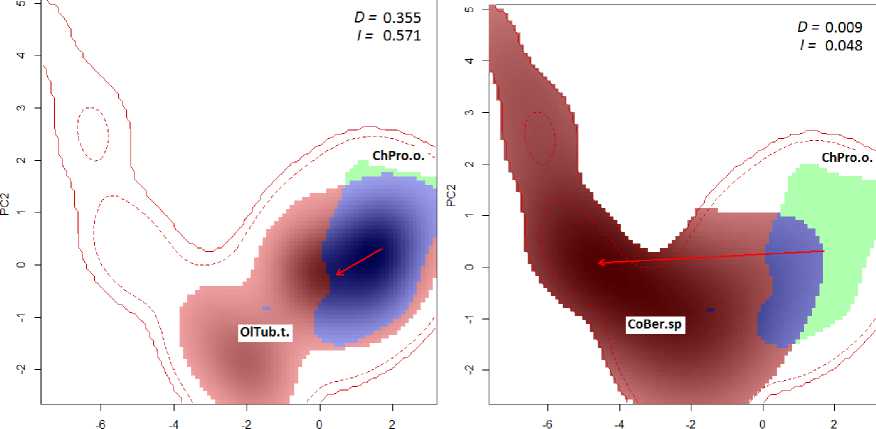
РС1 PCI
Рис. 2. Ординационные РСА-диаграммы попарного перекрытия экологических ниш трех видов: Prodia-mesa olivacea (ChPro.o.), Tubifex tubifex (OlTub.t.) и Berosus sp. (CoBer.sp). Красными контурными линиями обозначены границы изучаемой области (полной и с удалением 5% маргинальных участков). Красным и зеленым цветом закрашены ниши двух сравниваемых видов, а синим – совместно используемая область экологического пространства. Конец и начало стрелки показывают положение центроидов обеих ниш. Приведены значения индексов перекрытия ниш D Шенера и I Хеллингера.
Fig. 2. Coordination RSA diagrams of pairwise overlap of ecological niches of three types: Prodiamesa olivacea (ChPro.o.), Tubifex tubifex (OlTub.t.) and Berosus sp. (CoBer.sp). Red contour lines indicate the boundaries of the studied area (complete and with the removal of 5% of marginal areas). The niches of the two species being compared are painted in red and green, and the shared area of ecological space is painted in blue. The end and beginning of the arrow show the position of the centroids of both niches. The values of the overlap indices of the niches D Schoener and I Hellinger are given.
РСА-анализ проводился с привлечением 7 геоклиматических переменных (температура и уровень осадков, высота и шероховатость рельефа и др.) и 4 биотопических показателей (минерализация, насыщение кислородом O 2 , содержание ионов аммония NH 4 + и категория донного грунта). Дополнительно можно выяснить, какими конкретно переменными среды обусловлено различие экологических ниш. На рис. 3 показаны графики плотности распределения по шкалам важнейших абиотических факторов встречаемости видов Prodiamesa olivacea и Berosus sp., наиболее «контрастных» по условиям своего обитания.
ЗАКЛЮЧЕНИЕ
Моделирование экологических ниш и ареалов популяций является эффективным инструментом проведения биосферных исследований, который позволяет существенно расширить биологические представления об экологии видов и перевести их на строгую количественную основу. Возникает возможность рассматривать наблюдаемые явления в пространственновременном контексте: оценить скорость распространения инвазивных видов (Di Cola et al., 2017) или сокращения биопродуктивности за различные периоды времени; исследовать механизмы сосуществования партеногенетических форм животных (Petrosyan et al., 2020); выявить специфические пространственные закономерности организации мета-сообществ рамках крупномасштабной экосистемы (Шитиков и др., 2021b); осуществить выделение статистически значимых изолированных зон, на границе которых проис- ходит сукцессионная смена видового состава и т.д. К сожалению, современная трактовка термина "экологическая ниша" несколько обесценивается: под ней понимается лишь простейшая фундаментальная ниша Гринелла, а основополагающие теоретические представления Элтона Хатчинсона и МакАртура выносятся, как бы за скоб-ки1. В определенной мере прогресс в этом направлении реализуется в рамках иерархических моделей сообществ видов HMSC (Hierarchical Modelling of Species Communities – Ovas-kainen, Abrego, 2020; Шитиков и др., 2021а), в которых параметризация комплексного пространства ниш идет с использованием абиотических факторов, филогенетического сигнала, функциональных характеристик видов, пространственной автокорреляции и межвидовых взаимодействий в сообществе.
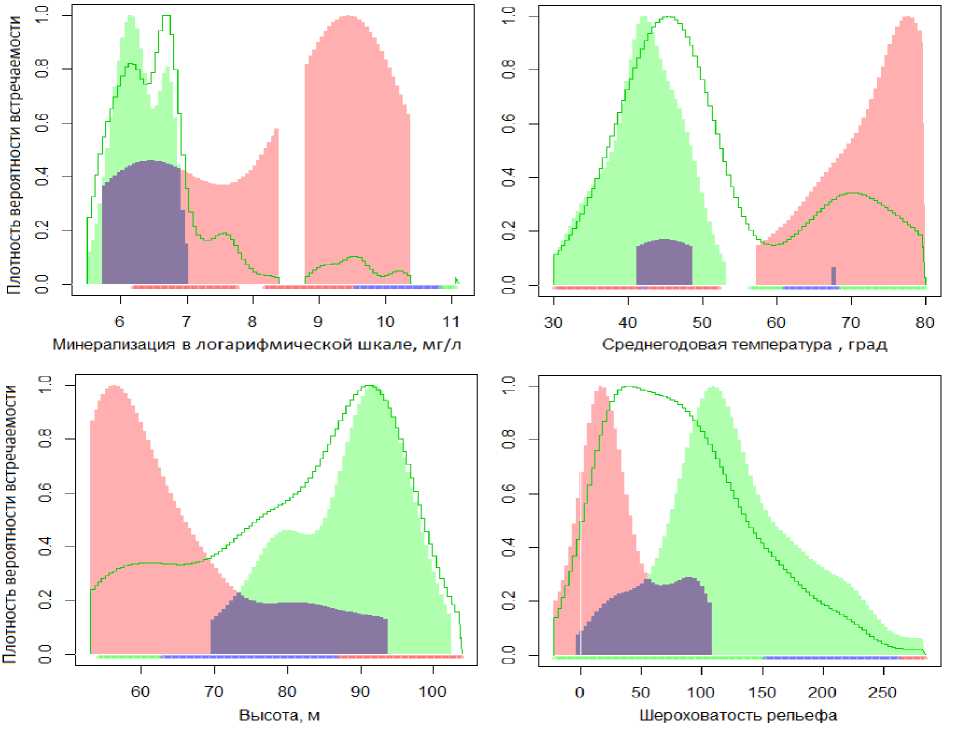
Рис. 3. Графики плотности распределения встречаемости Prodiamesa olivacea (зеленым цветом) и Berosus sp. (оранжевым цветом) по шкалам абиотических факторов. Синим цветом показана область совместной встречаемости. Зеленой контурной линией обозначена кривая плотности распределения для всех точек наблюдений.
Fig. 3. Graphs of the distribution density of the occurrence of Prodiamesa olivacea (in green) and Berosus sp. (in orange) according to the scales of abiotic factors. The blue color shows the area of joint occurrence. The green contour line indicates the distribution density curve for all observation points.
1 Здесь следует несколько слов сказать о «пульсирующей» многомерной экологической нише, представления о которой активно развивает И.Ю. Усманов и его школа (Усманов и др., 2014, 2016; Мавлетова-Чистякова и др., 2017). «Можно предположить, что пульсирующая смена лимитов является характерной, если не обязательной чертой индивидуальных ниш растений. Новым методологическим приемом, иначе оценивающим сложность и стохастичность живых и неживых систем разного уровня, является анализ их фрактальной или мультифрактальной организации. Принципиально важной характеристикой фракталов является возможность формирования самоподобных структур на всем поле, на котором выстраивается анализируемая иерархия. <…> Учитывая стохастический, пульсирующий характер смены лимитов - осей многомерной экологической ниши, растения должны для каждой индивидуальной ниши реализовать соответствующий набор адаптаций. В связи с прикрепленным образом жизни в условиях пульсирующей смены лимитов целесообразно ввести понятие индивидуальной ниши растения. Полученные данные ставят вопрос об организации адаптивного потенциала растений, который обеспечивал бы выживание растений в пульсирующих многомерных экологических нишах» (Усманов и др., 2016, с. 527, 528).
Список литературы Экологические ниши, их современная интерпретация и перспективы моделирования
- Зинченко Т.Д. Эколого-фаунистическая характеристика хирономид (Diptera, Chhironomidae) малых рек бассейна Cредней и Нижней Волги (Атлас). Тольятти: Кассандра, 2011. 258 с.
- Лисовский А.А., Дудов С.В., Оболенская Е.В. Преимущества и ограничения использования методов экологического моделирования ареалов. 1. Общие подходы // Журн. общ. биологии. 2020. Т. 81, № 2. С. 123–134.
- Мавлетова-Чистякова М.В., Щербаков А.В., Иванов В.Б., Юмагулова Э.Р., Усманов И.Ю. Пульсирующая мозаичность параметров почв Южного Зауралья // Вестн. Нижневартов. гос. ун-та. 2017. № 4. С. 124-133.
- Усманов И.Ю., Семенова И.Н., Щербаков А.В., Суюндуков Я.Т. Эндемичные экологические ниши Южного (Башкирского) Зауралья: многомерность и флуктуирующие режимы // Вестн. Башкир. гос. аграр. ун-та. 2014. № 1. С. 16-21.
- Усманов И.Ю., Щербаков А.В., Мавлетова М.В., Юмагулова Э.Р., Иванов В.Б., Александрова В.В. Пульсирующая многомерная экологическая ниша растений: расширение объема понятия // Изв. Самар. НЦ РАН. 2016. Т. 18, № 2 (2). С. 525-529.
- Шитиков В.К. Модели пространственного распределения видов. 2020. URL: https://stok1946.blogspot.com/2020/11/sdm.html (датаобращения: 10.10.2020)
- Шитиков В.К., Зинченко Т.Д., Головатюк Л.В. Модели совместного распределения видов на примере донных сообществ малых рек Волжского бассейна // Журнал общей биологии. 2021а. Т. 82, № 2. С. 143-154.
- Шитиков В.К., Зинченко Т.Д., Головатюк Л.В. Модели максимальной энтропии и пространственное распределение донных сообществ Средней и Нижней Волги // Российский журнал прикладной экологии. 2021b. № 2. С. 10-16.
- Zinchenko T.D. Ekologo-faunisticheskaia kharakteristika khironomid (Diptera, Chhironomidae) malykh rek basseina Crednei i Nizhnei Volgi (Atlas).
- [Ecological and faunal review of chironomids (Diptera, Chironomidae) small rivers in Middle and Low Volga basin (Atlas)]. Togliatti: Kassandra, 2011. 258 p. (In Russian).
- Lisovsky A.A., Dudov S.V., Obolenskaya E.V. Preimushchestva i ogranicheniia ispol'zovaniia metodov ekologicheskogo modelirovaniia arealov. 1. Obshchie podkhody [advantages and limitations of application of the species distribution modeling methods. 1. A general approach] // Zhurnal obshchey biologii. 2020. T. 81, № 2. P. 123-134. (In Russian).
- Chase J.M., Leibold M. Ecological Niches: Linking Classical and Contemporary Approaches. University of Chicago Press, Chicago and London, 2003. 212 p.
- Mavletova-Chistiakova M.V., Shcherbakov A.V., Ivanov V.B., Yumagulova E.R., Usmanov I.Yu. Pul'siruiushchaia mozaichnost' parametrov pochv Iuzhnogo Zaural'ia [Pulsing mosaic of soils in the Southern Trans-Ural region] // Vestnik Nizhnevartovartovskogo gosudarstvennogo universitetata. 2017. № 4. P. 124-133. (In Russian).
- Usmanov I.Yu., Semenova I.N., Shcherbakov A.V., Suyundukov Y.T. Endemichnye ekologicheskie nishi Iuzhnogo (Bashkirskogo) Zaural'ia: mnogomernost' i fluktuiruiushchierezhimy [Endemic ecological niches of south (Bashkir) trans-urals: multidimensional and fluctuating modes] // Vestnik Bashkirskogo gosudarstvennogo agrarnogo universiteta. 2014. № 1. P. 16-21. (In Russian).
- Usmanov I.Yu., Shcherbakov A.V., Mavletova M.V., Yumagulova E.R., Ivanov V.B., Aleksandrova V.V. Pul'siruiushchaia mnogomernaia ekologicheskaia nisha rastenii: rasshirenie ob"ema poniatiia [The pulsing multidimensional ecological niche of plants: extension of the concept] // Izvestia Samarskogo nauchnogo tsentra RAN. 2016. Vol. 18, No. 2 (2). P. 525-529. (In Russian).
- Shitikov V.К. Modeli prostranstvennogo raspredelenia vidov. [Models of spatial distribution of species]. 2020. (In Russian). URL: https://stok1946.blogspot.com/2020/11/sdm.html (accessed 10.10.2020).
- Shitikov V.К., Zinchenko T.D., Golovatyuk L.V. Modeli sovmestnogo raspredeleniia vidov na primere donnykh soobshchestv malykh rek volzhskogo basseina[Models of joint distribution of species with benthic communities from the small rivers of the Volga basin as a case study] // Zhurnal obshchei biologii. 2021a. Vol. 82, No. 2. P. 143-154. (In Russian).
- Shitikov V.К., Zinchenko T.D., Golovatyuk L.V. Modeli maksimal'noi entropii i prostranstvennoe raspredelenie donnykh soobshchestv Srednei i Nizhnei Volgi [Maximum entropy models and spatial distribution of bottom community species on the territory of the Middle and Lower Volga region] // Rossiiskii zhurnal prikladnoi ekologii. 2021b. No. 2. P. 10-16. DOI: 10.24852/2411-7374.2021.2.10.16. (In Russian).
- Barve N., Barve V., Jiménez-Valverde A., Lira-Noriega A., Maher S., Peterson A.T., Soberón J., Villalobos F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling // Ecological Modelling. 2011. Vol. 222(11). P. 1810-1819.
- Broennimann O., Fitzpatrick M.C., Pearman P.B., Petitpierre B., Pellissier L., Yoccoz N.G., Thuiller W., Fortin M.J., Randin C., Zimmermann N.E., Graham C.H., Guisan A. Measuring ecological niche overlap from occurrence and spatial environmental data // Global Ecology and Biogeography. 2012. Vol. 21. P. 481-497.
- Brown J.H., Stevens G.C., Kaufman D.M. Te geographic range: Size, shape, boundaries, and internal structure // Annual Review of Ecology and Systematics. 1996. Vol. 27. P. 597-623.
- Di Cola V., Broennimann O., Petitpierre B., Breiner F.T., D’Amen M., Randin C., Engler R., Pottier J., Pio D., Dubuis A., Pellissier R.G., Mateo R.G., Hordijk W., Salamin N., Guisan A. Ecospat: an R package to support spatial analyses and modeling of species niches and distributions // Ecography. 2017. Vol. 40. P. 1-14.
- Elton C. Animal Ecology. London: Sedgwick and Jackson, 1927. Franklin J. Mapping Species Distributions: Spatial Inference and Prediction. Cambridge: Cambridge University Press, 2009. 320 p.
- Gaston K.J. The structure and dynamics of geographic ranges. Oxford: Oxford University Press. 2003. 266 p.
- Grinnell J. Field tests of theories concerning distributional control. // American Naturalist. 1917. Vol. 51. P. 115-128.
- Grinnell J. Geography and evolution. // Ecology. 1924. Vol. 5. P. 225-229.
- Guisan A., Thuiller W. Predicting species distribution: offering more than simple habitat models. // Ecology Letters. 2005. Vol. 8. P. 993-1009.
- Guisan A., Thuiller W., Zimmermann N.E. Habitat Suitability and Distribution Models:With Applications in R. Cambridge: Cambridge University Press, 2017. 478 p.
- Hirzel A.H., Helfer V., Metral F. Assessing habitatsuitability models with a virtual species // Ecol. Model. 2001. Vol. 145. P. 111-121.
- Hutchinson G.E. Concluding remarks. // Cold Spring Harb. Symp. Quant Biol, 1957. Vol. 22. P. 415-427.
- Hutchinson G.E. An Introduction to Population Ecology. New Haven: Yale University Press, 1978.
- Leroy B., Meynard C.N., Bellard C., Courchamp F. virtualspecies: an R package to generate virtual species distributions // Ecography. 2016. Vol. 39. P. 599-607.
- MacArthur R. The theory of the niche. / In: Population Biology and Evolution (eds. K.C. Lewontin). Syracuse: Syracuse University Press, 1968. P. 159-176.
- Melo-Merino S.M., Reyes-Bonilla H., Lira-Noriega A. Ecological niche models and species distribution models in marine environments: A literature review and spatial analysis of evidence // Ecol. Model. 2020. Vol. 415. No. 108837. P. 1-35.
- Norberg A., Abrego N., Blanchet F.G., Adler F.R., Anderson B.J. et al. A comprehensive evaluation of predictive performance of 33 species distribution models at spe ies and community levels // Ecol. Monogr. 2019. Vol. 89, No 3. P. e01370.
- Ovaskainen O., Abrego N. Species Distribution Modelling: With Applications in R. Cambridge: Cambridge Univ. Press, 2020. 370 p.
- Peterson A.T., Soberón J. Species distribution modeling and ecological niche modeling: Getting the conceptsright. // Nat. Conserv. 2012. Vol. 10. P. 102-107.
- Peterson A.T., Soberón J., Pearson R.G., Anderson R.P., Martínez-Meyer E., Nakamura M., Araújo M.B. Ecological Niches and Geographic Distributions (MPB-49). Princeton: Princeton Univ. Press, 2011. 328 p.
- Petrosyan V., Osipov F., Bobrov V., Dergunova N., Omelchenko A., Varshavskiy A., Danielyan F., Arakelyan M. Species Distribution Models and Niche Partitioning among Unisexual Darevskia dahli and Its Parental Bisexual (D. portschinskii, D. mixta) Rock Lizards in the Caucasus // Mathematics. 2020. Vol. 8. P. 1329-1350. Doi:10.3390/math8081329
- Phillips S.J., Anderson R.P., Schapire R.E. Maximum entropy modeling of species geographic distributions // Ecol. Model. 2006. Vol. 190. № 3-4. P. 231-259.
- Qiao H., Peterson A.T., Campbell L.P., Soberón J., Escobar L.E. NicheA: creating virtual species and ecologicalniches in multivariate environmental scenarios // Ecography. 2016. Vol. 39. P. 805-813.
- Soberón J. Grinnellian and Eltonian niches and geographic distributions of species // Ecol. Lett. 2007. Vol. 10. P. 1115-1123.
- Thuiller W., Lafourcade B., Engler R., Araújo M.B. BIOMOD – a platform for ensemble forecasting of species distributions. // Ecography. 2009. Vol. 32. P. 369-373
- Wallace A.R. On the zoological geography of the Malay Archipelago // Proceedings of the Linnean Society of London. 1860. Vol. 4. P. 172-184.
- Warren D.L., Glor R.E., Turelli M. Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution // Evolution. 2008. Vol. 62. P. 2868-2883.
- Whittaker R.H., Levin S.A., Root R.B. Niche, habitat, ecotope. // Am. Nat. 1973. Vol. 955. P. 321-338.
