Экологические особенности промысловых фукусовых водорослей Мурманского побережья Баренцева моря

Автор: Шошина Елена Васильевна, Капков Валентин Иванович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Статья в выпуске: 1 т.17, 2014 года.
Бесплатный доступ
Проведено исследование экологических адаптационных приспособлений популяций фукусовых водорослей в составе литорального фитоценоза, которые обеспечивают целостность прибрежной экосистемы в условиях Баренцева моря.
Фукусовые водоросли, литоральные фитоценозы, мурманское побережье баренцева моря
Короткий адрес: https://sciup.org/14294668
IDR: 14294668
Текст научной статьи Экологические особенности промысловых фукусовых водорослей Мурманского побережья Баренцева моря
Бурые водоросли играют главную роль в формировании и функционировании литоральных биоценозов прибрежных экосистем Баренцева моря, характеризующихся высокими видовым разнообразием и биомассой бентосных организмов. Наибольший вклад в биомассу органического вещества здесь вносят фукусовые водоросли – эдификаторы фитоценозов, имеющие длительный цикл развития и определяющие структуру биотического сообщества. Доля литоральных водорослей в условиях открытого и скалистого Мурманского побережья, где большие осушки встречаются редко, в общей продукции бентических сообществ составляет 6 % ( Кузнецов, Шошина , 2003). Существенное значение в пищевых цепях принадлежит бурым водорослям: ими (особенно проростками), питаются многочисленные беспозвоночные фитофаги, которые при этом, прореживая новые поселения, регулируют видовое разнообразие и вертикальную структуру фитоценозов ( Lubchenko, Cubit , 1980). Известна роль бурых водорослей в процессах очищения вод от тяжелых металлов и нефтяных углеводородов ( Капков, Беленикина , 2007; Капков, Шошина, 2011; Воскобойников, Пуговкин , 2012).
Промысловые фукусовые водоросли, которые доминируют в литоральных сообществах, характеризуются широкой экологической валентностью. Они обитают в условиях сезонных и приливноотливных перепадов температуры и солнечной радиации, в значительном интервале солености, а также при постоянно возрастающей антропогенной нагрузке на прибрежные морские экосистемы. Бурые водоросли входят в состав многоярусных структурированных биоценозов с высокой биологической информацией, в которые интегрированы многочисленные популяции гидробионтов. Устойчивость популяций к экстремальным экологическим факторам, включая конкуренцию с партнерами по сообществу и пресс фитофагов, обеспечивается посредством мобилизации ферментативной и гуморальной регуляции, в том числе снижением обмена веществ в периоды замедленного роста. Существование сложного многолетнего фитоценоза благоприятствует развитию у популяций, входящих в растительное сообщество, приспособлений к совместному обитанию и увеличивает шансы на выживание каждого вида в условиях литорали северных морей.
Развитие промысловых фукусовых водорослей в экосистеме литорали контролируют в основном первичные экологические факторы, среди которых ведущим является наличие твердого субстрата, необходимого для прикрепления слоевища. Следующим важным фактором, определяющим распространение водорослей на литорали, оказываются прибрежные и приливно-отливные течения, которые перемещают водные массы в горизонтальном и вертикальном направлениях. Бентосные водоросли нуждаются в постоянном движении воды, которая обеспечивает водоросли необходимыми ресурсами, удаляет продукты метаболизма и создает условия для осуществления вегетативных и генеративных функций. В водных экосистемах передача информационных сигналов посредством метаболитов происходит медленно, поскольку здесь важна не столько скорость, сколько продолжительность ответных реакций, обеспечивающих развитие биотического сообщества. Функция гуморального регулирования метаболизма у литоральных водорослей направлена на сохранение состояния внутренней среды, поддерживающей оптимальный рост популяции в биотическом сообществе ( Хайлов , 1971; Iken , 1996).
В северных морях фукусовые водоросли обитают на осушках и дважды в сутки значительную часть времени находятся в воздушной среде. Во время отлива слоевища водорослей обезвоживаются и подвергаются световому и температурному шоку. Поэтому клеточная стенка бурых водорослей прочная и многослойная: внутренний слой оболочки состоит из целлюлозы, а внешний (матриксный) образован сульфатированными полисахаридами, в состав которых входят альгиновая кислота, альгинаты и фукоиданы. На долю альгинатов матрикса приходится до 40 % сухого вещества; они способны к 200300-кратному связыванию воды. Благодаря этому водоросли образуют много слизи, что и помогает клеткам удерживать воду во время отлива. Потере структурно связанной воды слоевищами препятствует также высокий осмотический потенциал клеток бурых водорослей. Способность фукусовых водорослей выдерживать значительные суточные и сезонные перепады температуры среды связана с наличием криопротекторов, к числу которых относятся ламинараны и другие низкомолекулярные полисахариды. Содержание ламинаранов у фукусовых водорослей невелико (1-4 %), но их роль в преодолении колебаний температуры окружающей среды во время отлива в зимний период весьма существенна ( Облучинская, Шошина , 2008).
Во время отлива литорали, особенно в период полярного дня, фукусовые водоросли подвергаются мощному потоку солнечной радиации. Бесцветные флортаннины, содержащиеся в везикулах клеток, на воздухе окисляются, и водоросли приобретают темную окраску, что препятствует воздействию солнечных лучей во время осушки. Полагают также, что функция флортаннинов связана и с защитой талломов от поедания водорослей фитофагами ( Hoek et al ., 1995).
В течение последнего столетия промысловые фукусовые водоросли Баренцева моря привлекали пристальное внимание многих исследователей, которые изучали видовой состав, распространение, морфологию, рост, фенологию размножения и продукционные характеристики. Однако остаются практически не исследованными многие особенности экологии фукусовых водорослей, обитающих в специфических условиях литоральной зоны арктических морей. В этой связи целью настоящей работы является анализ экологических адаптационных приспособлений популяций фукусовых водорослей в составе литорального фитоценоза, которые обеспечивают целостность прибрежной экосистемы в условиях Баренцева моря.
2. Регион исследования. Материал и методы
Данная работа является продолжением публикации результатов исследований биологических и экологических особенностей фукусовых водорослей в литоральных фитоценозах в условиях арктических морей ( Шошина , 1998; Завалко, Шошина , 2008; Шошина и др ., 2012; Гончарова, Шошина , 2013). Изучение экологических особенностей бурых промысловых водорослей проводили в акватории Мурманского побережья Баренцева моря (рис. 1). В качестве объектов исследования выбраны популяции фукусовых водорослей Fucus vesiculosus L., F. distichus L., F. serratus L. и Ascophyllum nodosum (L.) Le Jolis. Материалом служили многолетние наблюдения и сборы проб водорослей в губах и бухтах моря с различными гидрологическими и гидрохимическими условиями. Пробы отбирали во время отлива в характерных для каждого вида водорослей местах обитания методом вертикальных разрезов и 3-5 пробных площадок с использованием рамок 50X50 см2. Фукусовые водоросли каждой пробы разделяли по видовому, возрастному и половому признакам. В лабораторных условиях определяли численность водорослей в каждой возрастной группе ( Ni , экз./м²), биомассу популяции каждого вида ( Wi , г/м²) и площадь поверхности слоевища ( Si , см²); площадь поверхности талломов измеряли стандартным весовым способом по бумажным контурам, а также помощью компьютерной системы анализа изображения ( Завалко, Шошина , 2008). Возраст растений у фукусов оценивали по числу дихотомических ветвлений, считая, что в год образуется два ветвления; на слоевище аскофиллума на Мурманском побережье ежегодно формируется один порядок воздушных пузырей и одно дихотомическое ветвление ( Кузнецов , 1960). Полученные результаты обрабатывали с помощью пакета программ "Microsoft Office Excel".
3. Результаты и их обсуждение
В районах проведения исследований на Мурманском побережье, где видовое разнообразие бурых водорослей наиболее широко представлено, фукусовые доминируют на литорали. При этом Ascophyllum nodosum и Fucus vesiculosus занимают верхний горизонт, F. distichus – средний, а F. serratus предпочитает нижний горизонт литорали. В исследованных районах на литорали в ассоциациях фукусовых обитают также бурые водоросли Dictyosiphon foeniculaceus , Stictysiphon tortilis , Sphaecelaria arctica ; красные – Dumontia incrassata , Polyides caprinus ; зеленые – Cladophora sp. , Enteromorpha prolifera , Monostroma grevillii , Rhizoclonium riparium и Ulothrix pseudoflaccа и многие другие виды, однако биомасса каждого из них на 1-3 порядка ниже, чем у популяции фукоидов. Характерно, что видовое разнообразие водорослей существенно возрастает от верхнего к нижнему горизонтам литорали и особенно в переходной зоне – на границе литорали и сублиторали.
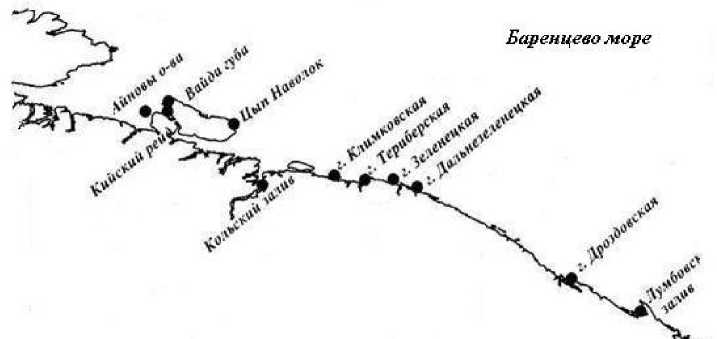
Рис. 1. Карта района исследования и отбора проб
В прибрежной зоне Баренцева моря наблюдаются выход коренных пород, скопления грубообломочного материала и валунно-галечно-гравийные отложения, которые и формируют фации донного ландшафта. В местах выхода скальных пород и на крупных валунах формируются сообщества фукусовых водорослей с максимальной биомассой, но с относительно невысоким видовым разнообразием. На валунно-галечных грунтах численность водорослей и видовое разнообразие заметно увеличиваются на фоне снижения биомассы сообщества. На гравийно-галечных грунтах произрастают в основном однолетние виды и лишь немногочисленные проростки фукусовых водорослей. Из-за штормов и перекатывания гальки взрослые формы многолетних водорослей на галечной литорали практически не встречаются.
Картина распределения фукусовых водорослей на литорали заметно меняется в зависимости от степени прибоя. В губах Восточного Мурмана F. vesiculosus обитает как на скалистых субстратах, так и на валунно-галечных отложениях, занимая защищенные от прибоя участки литорали, но практически отсутствует на открытых сильному прибою скалах. С некоторым уменьшением прибойности на скалистых участках литорали F. vesiculosus часто образует узкий ярус с F. distichus , а на валунногалечных отложениях становится доминирующим видом, вытесняя партнеров по сообществу. Популяции A. nodosum в бухтах с сильным прибоем и проливах с мощным течением становятся господствующими на верхнем горизонте скалистой литорали, на валунно-галечных отложениях произрастают совместно с F. vesiculosus , если прибой умеренный или слабый, а на гравийных россыпях при слабом волнении уступают место F. distichus .
На скалистых субстратах литорали A. nodosum образует сплошные густые заросли, под пологом которых молодые проростки F. vesiculosus практически лишены солнечного света. После шторма часто происходит обрыв талломов и прореживание зарослей аскофиллума. Видимо, в это время зиготы фукуса пузырчатого, осевшие на субстрат, прорастают и затем вытесняют А. nodosum и другие водоросли с твердого субстрата, при этом некоторые из них становятся эпифитами фукуса.
Ослаблению конкуренции между A. nodosum и F. vesiculosus за биотоп в верхнем и среднем горизонтах литорали способствуют сопутствующие виды, включая эпифитов и симбионтов, которые повышают резистентную устойчивость водорослевого сообщества. На талломах A. nodosum обитает облигатный эпифит - красная водоросль Polysiphonia lanosa . Обычно багрянка поселяется на поврежденных участках таллома и на месте сброшенных рецептакул, используя метаболиты аскофиллума в качестве питательных веществ. Сообщалось также, что красные водоросли выделяют соединения, подавляющие рост ювенильных стадий бурых водорослей - конкурентов аскофиллума ( Lining, Garbary , 1992; Denboh et al. , 1997).
Очевидно, в экстремальных условиях литорали северных морей жесткая конкуренция уступает место сосуществованию партнеров, при котором каждая популяция достигает сбалансированной плотности по отношению к другим членам сообщества.
В литоральных фитоценозах популяции фукусовых водорослей характеризуются длинными возрастными рядами, в которых преобладают молодые особи и сравнительно большое количество фертильных растений. В верхнем горизонте литорали A. nodosum и F. vesiculosus растут кустами, в которых представлены особи разного возраста до 750 и 1700 экз./м² соответственно. При этом наблюдается заметное уменьшение особей старших возрастных групп. В то время как средний и нижний горизонты занимают F. distichus и F. serratus , водоросли растут группами всего по 3-10 одновозрастных растений численностью 120 и 170 экз./м2; в фитоценозе, как правило, доминируют 4-6-летние особи (рис. 2).
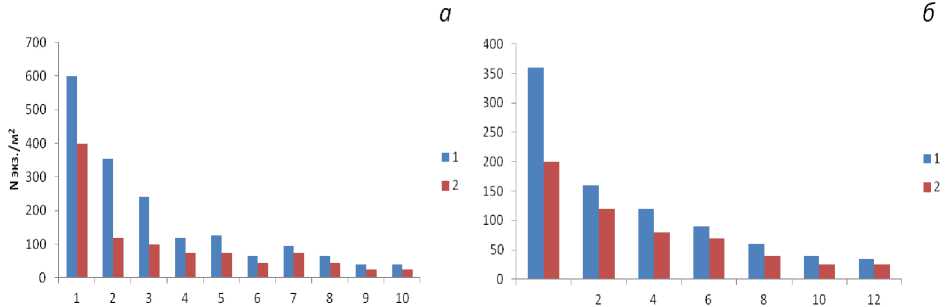
Рис. 2. Численность фукусовых водорослей разных возрастных групп в губе Зеленецкой (1) и бухте Завалишина, губа Териберская (2): а - F. vesiculosus;
б - A. nodosum . По оси абсцисс - возраст (год), по оси ординат - число особей на 1 м2
У морских водорослей скорость роста таллома в значительной степени зависит от величины солнечной радиации и прежде всего от способности клеток поглощать красные и синие лучи. Анатомическое строение слоевищ фукусовых водорослей имеет специфические особенности: под слоем эпидермиса у них располагаются крупные пигментированные клетки, в которых сосредоточена основная масса хлоропластов. С возрастом количество ассимиляционных слоев и хлоропластов в клетках увеличивается как в апикальном участке таллома, так и дихотомических ответвлениях, причем объем пластидома возрастает до 50 % массы клетки. Хлоропласты бурых водорослей, которые локализованы в поверхностном слое таллома, различаются по размеру и форме; они изменяются с возрастом водоросли. Кроме хлорофиллов а и с у бурых водорослей обнаружены каротиноиды: фукоксантин, виолоксантин, зеаксантин, антераксантин, неоксантин, диадиноксантин, диатоксантин и каротины ( Hoek et al ., 1995). Наличие большого набора пигментов и строение слоевища обеспечивают максимальный выход фотосинтеза как в водной, так и в воздушной среде и предохраняют водоросли от солнечной радиации во время отлива ( Воскобойников, Камнев , 1991).
Данные о размерно-весовой структуре популяции баренцевоморских фукусовых водорослей представлены для двух характерных мест обитания на восточном побережье: открытой для волновых явлений губы Зеленецкой и закрытой от прямого воздействия волн бухты Завалишина в губе Териберской (рис. 3). Рост слоевищ фукусовых происходит за счет деления клеток верхушечной меристемы, причем формирование таллома в результате дихотомического ветвления продолжается у разных видов с разной скоростью в течение всего жизненного цикла. У трех видов фукусов ( F. vesiculosus , F. distichus , F. serratus ) талломы растут линейно, и скорость роста существенно выше в открытых для волн бухтах как результат хорошего водообмена и обогащения среды биогенными элементами. Линейный рост талломов у А. nodosum происходит в течение первых шести лет, а затем скорость роста замедляется, и размеры растений старших возрастных групп остаются практически одинаковыми до конца жизни.
Масса талломов фукусов в течение жизненного цикла возрастает экспоненциально как за счет дихотомического ветвления, так и за счет структурных перестроек и дифференцировки тканей, а также в результате накопления органических веществ водорослью. Среди фукусов самые мощные слоевища характерны для F. serratus , который обитает в наиболее благоприятных условиях нижнего горизонта литорали. Масса слоевищ А. nodosum с возрастом, в отличие от других фукусовых, остается практически постоянной за счет снижения скорости роста в результате механических повреждений таллома. При внешнем морфологическом сходстве талломы фукусовых водорослей различаются: у фукусов слоевище представляет собой цельную уплощенную ветвь, рецептакулы образуются на верхушке ветвей, после их сброса ветви прекращают свой рост. У аскофиллума таллом состоит из сдавленных цилиндрических образований, которые при сильном волнении моря часто обрываются, на местах обрыва формируются боковые ветви, рецептакулы в массе образуются по краю всех ветвей, что и сказывается на массе водорослей старших возрастных групп (рис. 4).
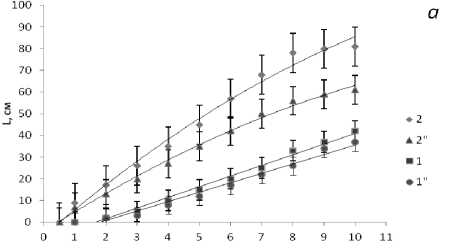
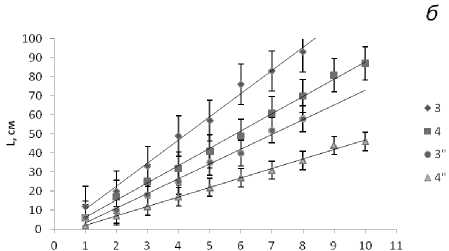
Рис. 3. Размеры слоевищ фукусовых водорослей в Зеленецкой (1-4) и Териберской (1"-4") губах Восточного Мурмана: а - F. vesiculosus (1), A. nodosum (2); б - F. distichus (3), F. serratus (4).
По оси абсцисс - длина слоевища (в см), по оси ординат - возраст (год)
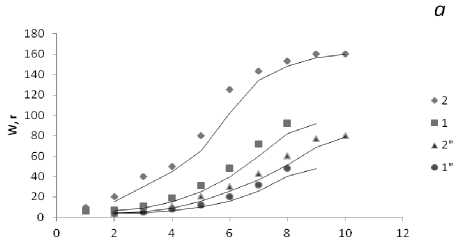
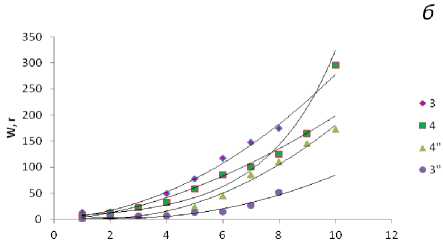
Рис. 4. Масса слоевищ фукусовых водорослей в Зеленецкой (1-4) и Териберской (1"-4") губах Восточного Мурмана. Обозначения те же, что и на рис. 3.
По оси абсцисс - масса слоевища (в г), по оси ординат - возраст (год)
У фукусов ( F. distichus , F. serratus ) наблюдается существенное снижение скорости и интенсивности роста слоевищ от весны к осени. Установлено, что в условиях полярного дня рост талломов происходит с высокой скоростью, при этом блокируется формирование рецептакул. Осенью в условиях короткого дня, когда начинается образование и развитие рецептакул, вегетативный рост талломов снижается. Известно, что фотопериод контролирует чередование вегетативного роста и репродукционного процесса у бурых водорослей посредством синтеза аллохимических соединений, тормозящих собственный рост популяций ( Luning, Dieck , 1989).
Площадь поверхности слоевищ (как и масса) у фукусовых водорослей увеличивается экспоненциально начиная с 4-летнего возраста, причем у F. vesiculosus (обитателя верхнего горизонта литорали) и F. serratus (обитателя нижнего горизонта) этот процесс происходит с более высокой скоростью, о чем свидетельствуют наклоны кривых линий изменения площади таллома (рис. 5). Существенное увеличение площади слоевищ в процессе роста популяции характерно для водоросли F. serratus , которая вступает конкурентные отношения с F. distichus (в среднем горизонте литорали) и с ламинарией (в нижнем горизонте).
Изучение половой структуры фитоценозов фукусовых водорослей позволило установить соотношение женских и мужских особей в популяциях разных видов в зависимости от занимаемого биотопа. В популяциях двудомных видов фукусовых верхнего горизонта литорали A. nodosum и F. vesiculosus, образующих плотные заросли, количество женских особей было в 1,5 раза больше, чем мужских. В сильно разреженных зарослях F. vesiculosus в самом верхнем горизонте литорали женские растения составляли иногда до 90 % от общего числа особей в популяции. В популяциях F. serratus, который обычно занимает нижний горизонт литорали, соотношение женских и мужских особей было близким к 1 : 1. Следует подчеркнуть, что соотношение женских и мужских особей в популяциях исследованных видов фукусовых водорослей сохраняется практически у всех возрастных групп за исключением самых молодых и самых старых растений (рис. 6). Полученные нами результаты свидетельствуют о том, что половая структура популяции фукусовых водорослей в значительной мере определяется конкретными условиями занимаемого водорослью биотопа. Общая тенденция заключается в увеличении числа женских особей в стрессовых условиях в популяциях фукусовых водорослей, обитающих на литорали северных морей.
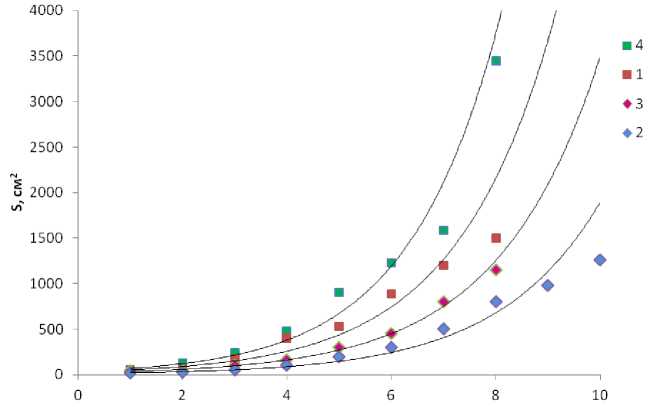
Рис. 5. Площадь поверхности слоевищ фукусовых водорослей в бухте Белокаменка (Кольский залив). Обозначения водорослей те же, что на рис. 3.
По оси абсцисс – площадь слоевищ (в см²), по оси ординат – возраст (год)
Баренцевоморские фукусовые водоросли приступают к размножению в возрасте 2-4 лет, т.е. значительно позднее, чем другие многолетние виды водорослей. Количество фертильных особей в популяциях разных видов фукусовых водорослей может отличаться в зависимости от экологических факторов: от 20-30 % у F. vesiculosus и F. serratus до 50 % у F. distichus и A. nodosum . При этом основная репродуктивная нагрузка приходится на 4-5-летние особи F. vesiculosus и на 5-7-летние растения A. nodosum ( Шошина , 1998).
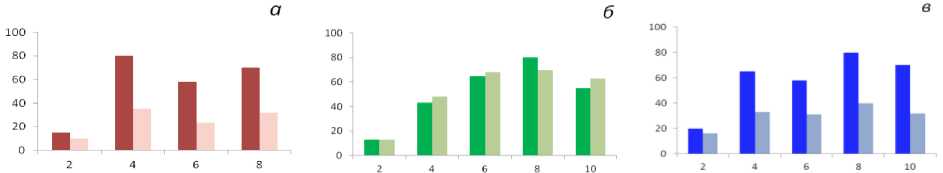
Рис. 6. Соотношение женских (темный цвет) и мужских (светлый цвет) фертильных особей двудомных фукусовых водорослей, губа Териберская: а – F. vesiculosus ; б – F. serratus ; в – A. nodosum
Развитие органов размножения (рецептакул) и образование гамет у разных видов фукусовых водорослей происходит в течение длительного времени. У F. vesiculosus рецептакулы закладываются в октябре, у F. distichus – в августе-октябре; гаметангии формируются и медленно созревают в течение зимы и весны, и только в июне происходит массовый выход антерозоидов и яйцеклеток в воду; т.е. развитие рецептакул занимает 9-12 месяцев. Выход гамет может продолжаться до августа, после чего рецептакулы у фукусов разрушаются. Фукусовая водоросль А. nodosum начинает формировать новые органы размножения в июне, их развитие продолжается в течение 11-12 месяцев, массовый выход гамет происходит в июне следующего года, рецептакулы со зрелыми гаметами у отдельных особей встречаются до августа. Более короткие сроки развития (и в другое время) органов размножения наблюдаются у обитателя нижнего горизонта литорали F. serratus. У этого вида рецептакулы закладываются в мае-июне и созревают к концу августа – началу сентября, причем водоросль способна производить гаметы вплоть до лета следующего года. В условиях Мурманского побережья сроки выхода гамет у фукусовых водорослей сближены, но тем не менее они несколько различаются у разных видов; это требует дополнительного исследования. В целом, в северной части высокобореальной подзоны все время года в сообществах водорослей занято и рационально распределено между размножающимися видами ( Кузнецов, Шошина , 2003).
Следует подчеркнуть особенности стратегии вегетативного роста и генеративного развития фукусовых водорослей, которые в том числе и позволяют этим водорослям не только произрастать, но и доминировать в такой сложной для жизни зоне, как литоральная. У фукусовых водорослей ( Ascophyllum nodosum , виды рода Fucus ) процессы роста и образования гамет разделены в пространстве своего слоевища: весной и начале лета (период интенсивного вегетативного роста на Мурманском побережье) одни ветви растут вегетативно, на других идет рост и созревание рецептакул (их закладка произошла в предыдущий вегетационный период). На молодых ветвях (текущего года развития) у фукусов рецептакулы начинают формироваться в конце лета или осенью. Таким образом, имеет место удлинение времени развития: образование побегов и формирование на них органов размножения захватывает два вегетационных сезона. В результате и закладка, и выход репродуктивных клеток приходится на летнее (более благоприятное) время года в условиях Арктики, что является характерной чертой фукусовых как долгоживущих видов.
У фукусовых водорослей высвобождение гамет из рецептакул синхронизировано с фотопериодом и приливно-отливными явлениями в море. Выход в воду антерозоидов и яйцеклеток происходит исключительно в дневное время суток, как правило, при сизигийных приливах. Дело в том, что во время отлива слоевище водоросли обсыхает, в результате чего происходит сдавливание рецептакул, которые защищены на время отлива от высыхания слизью. Во время последующего прилива стенки рецептакул разрушаются и гаметы выходят в воду. Мужские гаметы у фукусовых водорослей подвижны; они передвигаются в воде с помощью двух неравных жгутиков, один из которых (перистый) направлен вперед, а другой (короткий и гладкий) – в сторону и назад. При этом феромоны водорослей, выделяемые в окружающую среду с метаболитами, координируют движение гамет, а также препятствуют оплодотворению яйцеклеток другого вида ( Hoek et al ., 1995). Во время размножения фукусовых водорослей число антерозоидов и яйцеклеток может достигать десятков и сотен тысяч на 1 м² в год, однако количество производимых гамет оказывается на порядок больше, чем выбрасывается в воду ( Chapman , 1995). Так, у F. distichus из рецептакул в окружающую среду одновременно выходит только 30 % зрелых яйцеклеток, из которых только десятки от общего числа производимых гамет оплодотворятся антерозоидами и только несколько процентов зигот осядут на твердый субстрат и прорастут.
Необходимо отметить, что однодомная водоросль ( F. distichus ), у которой женские и мужские гаметы образуются на одном растении, имеет преимущество перед другими фукусовыми как при оплодотворении, так и при колонизации субстрата. В данном случае небольшая разница во времени прорастания зигот (всего в несколько часов), которая вначале едва различима, повышает конкурентную способность данного вида в сообществе. Процесс оплодотворения у фукусовых водорослей занимает не более двух часов. После слияния гамет зигота развивается без стадии покоя, что предотвращает ее смыв в открытое море во время отлива. Зиготы сразу же оседают на субстрат, приклеиваясь с помощью слизи, образуют первые ризоиды спустя несколько часов после закрепления на твердом субстрате и прорастают ( Chapman , 1995; Hoek et al ., 1995; Максимова , 2010). Последующее выживание ювенильных стадий фукусовых водорослей в значительной степени определяется влиянием биотических и абиотических экологических факторов. Количество выживших однодневных проростков А. nodosum и F. vesiculosus было в пять раз меньше среди зарослей родительских растений, чем на свободном твердом субстрате. А в открытых волновым воздействиям бухтах прикрепленными к натуральному субстрату оставалось не более 0,5 % от числа молодых проростков, осевших на искусственный субстрат, в то время как в закрытых бухтах на естественном субстрате сохранялось почти 80 % проростков ( Vadas et al ., 1990).
Среди бурых водорослей северных морей встречаются виды, обладающие резистентной устойчивостью к стрессовым факторам и сохраняющие свой тип стратегии в течение всего жизненного цикла, другие виды могут переносить стресс на определенных этапах развития популяции, третьи способны менять тип жизненной стратегии в зависимости от обстоятельств и вновь возвращаться к ней при благоприятных условиях. В принципе, в литоральном биотическом сообществе положительные взаимодействия между популяциями (комменсализм, мутуализм, протокооперация) являются более важными, чем жесткая конкуренция между видами, особенно на стадии формирования структурнофункциональной организации биотического сообщества и становления целостности экосистемы ( Алимов , 2000; Бурковский , 2006).
Развитие фитоценозов бурых водорослей на литорали северных морей контролируется первичными экологическими факторами: наличием твердого субстрата, величиной солнечной радиации, температурой и соленостью воды, наличием биогенных веществ и волновыми явлениями, которые выступают в качестве триггеров, запускающих вегетативные и генеративные процессы в популяциях водорослей. В то же время периодически воздействующие на литораль приливно-отливные явления не приводят к существенным изменениям в фитоценозах фукусовых водорослей. Они вызывают лишь допустимые отклонения в росте отдельных популяций водорослей: фитоценоз путем структурных перестроек устраняет негативные последствия подобных воздействий за счет упругой устойчивости, выработанной за длительный период существования на литорали. В случаях сильных штормовых воздействий возможны нарушения структурно-функциональной организации литорального фитоценоза, однако и этом случае произойдут лишь определенные изменения в соотношении популяций отдельных видов, поскольку за счет резистентной устойчивости сообщество продолжит свое существование. Устойчивость фитоценоза к экстремальным экологическим факторам обеспечивается мобилизацией ферментативных и гормональных регуляторов, которые замедляют обмен веществ водорослей в неблагоприятный для активного роста период.
Таким образом, баренцевоморские бурые водоросли являются видами-эдификаторами в сложных многоярусных фитоценозах литорали, обеспечивая функционирование бентосных сообществ. Фукусовым водорослям принадлежит главная роль в организации информационной составляющей литоральной экосистемы – в формировании трофических, топических и гуморальных связей биотического сообщества. Информационные процессы в экосистеме литорали обеспечивают структурнофункциональную организацию сообщества, направляя потоки вещества и энергии на сохранение целостности экосистемы.
4. Заключение
Анализ полученных результатов свидетельствует о том, что на развитие литоральных фитоценозов бурых водорослей оказывают влияние как абиотические, так и биотические факторы, соотношение которых регулирует численность отдельных популяций в биотическом сообществе. Развитие фитоценозов фукусовых водорослей контролируют солнечная радиация, температура, соленость, биогенные элементы, а также наличие твердого субстрата и приливно-отливные явления. Регуляция плотности популяций по принципу обратной связи обеспечивается биотическими взаимоотношениями между видами в сообществе в зависимости от биотического потенциала и биоценотической емкости среды.
Очевидно, что в экстремальных условиях обитания на литорали в фитоценозе жесткая конкуренция за ресурсы уступает место существованию партнеров по сообществу, при котором каждая популяция достигает сбалансированной плотности по отношению к другим членам сообщества.
Жизненный цикл фукусовых водорослей основан на принципе регулирования вегетативных и генеративных стадий, который в основных чертах сохраняется при воздействии экологических факторов. Такая система регулирования жизненных функций представляет собой характерную для литоральных водорослей особенность, обусловленную генетической структурой популяции. В то же время в отличие от детерминированного типа регулирования, свойственного определенному виду водорослей, у отдельных особей популяции процессы регулирования жизненных функций варьируют чрезвычайно широко. Показано, что жизненные стратегии баренцевоморских фукусовых водорослей включают в себя совокупность фенотипических адаптаций популяции к периодически меняющимся экологическим факторам среды.
Так, в экстремальных условиях обитания у фукусовых водорослей наблюдаются морфологические изменения талломов и в популяциях преобладают водоросли с относительно высоким отношением площади поверхности таллома к его массе, что позволяет отдельным видам занимать определенную экологическую нишу и получать преимущество в конкурентных отношениях с партнерами по сообществу. Экологические адаптации водорослей отражают как влияние определенного вида на биотическое сообщество, так и реакцию сообщества на присутствие конкретного вида в нем. Поэтому в экосистеме литорали обитают только виды, положительно влияющие на биотическое сообщество.
Поскольку фукусовые водоросли размножаются преимущественно половым способом, то большое количество производимых ими гамет, растянутые сроки их выброса в воду, короткий период процесса оплодотворения, развитие зигот без стадии покоя, быстрое прикрепление зигот к субстрату и последующее прорастание обеспечивают существование сложного фитоценоза, который определяет структурно-функциональную организацию биотического сообщества литорали.
