Экологический мониторинг морской биоты бухты Алексеева (залив Петра Великого)
")
Автор: Федорец Ю.В., Шарова О.А., Косьяненко А.А., Аксентов К.И., Раков В.А., Васильева Л.Е.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Природопользование и мониторинг
Статья в выпуске: 1-6 т.13, 2011 года.
Бесплатный доступ
Изучен видовой состав ихтиопланктона и макробентоса б. Алексеева. Установлено, что гибель икры рыб в море на ранних этапах развития связана в основном с факторами ветрового волнения, температуры и солености воды. Построена карта распределения ртути в поверхностном слое донных осадков.
Макробентос, ихтиопланктон, личинки асцидии, ртуть, бухта алексеева
Короткий адрес: https://sciup.org/148199871
IDR: 148199871 | УДК: 574
Marine biota ecological monitoring in Alekseev Bay (Peter the Great gulf)
The specific structure of ichthyoplankton and macrobenthos of Alekseev bay are studied. It is established that the destruction of fishes caviar in the sea at early stages of development is connected basically with factors of wind excitement, temperature and salinity of water. The map of mercury distribution in the surface layer of ground deposits was constructed.
Текст научной статьи Экологический мониторинг морской биоты бухты Алексеева (залив Петра Великого)
Цель настоящей работы: мониторинг морской биоты б. Алексеева. Для этого предстояло изучить видовой состав и количественное распределение макробентоса и ихтио-планктона, проследить изменения икры на разных стадиях развития в течение нерестового сезона; установить распределение ртути в поверхностном слое донных осадков.
Бухта Алексеева находится в югозападной части Амурского залива, вдается в северо-западный берег о. Попова (рис. 1 А). Вершина бухты мелководная, в средней части глубина увеличивается до 11 м и по направлению к выходу равномерно возрастает до 20 м. В августе 2006 и 2007 гг. температура воды в бухте практически не различалась и составляла в среднем 22-23ºС, с максимумом 29ºС. Осенью 2006 г. температура воды была 8-11ºС, что ниже чем в 2007 г. (10-13ºС). Соленость поверхностных вод летом колеблется от 24 до 32‰, зимой близка к 34‰ [3]. В 2010 г. температура воды в летние месяцы составила в среднем 15-24ºС, соленость поверхностных вод колебалась от 27 до 33‰.
Материал и методы. Макробентос собирали в б. Алексеева в июне-августе 2009 и 2010 гг. на 6 станциях дночерпателем Петерсена с площадью раскрытия 0,025 м2 (см. рис. 1 А). Пробы промывали через гидробиологические сита с ячеей 0,5 мм. Макробентос взвешивали на электронных весах с точностью до 0,01 г. Видовую принадлежность флоры и фауны определяли, а таксономическую принадлежность сверяли по «Таксономическому каталогу…» [7-10]. Для гранулометрического анализа грунт сушили при температуре 60°С в течение 12 часов, затем просеивали через почвенные сита. Структуру осадка определяли по существующей номенклатуре [11]. Ихтио-планктон собирали с мая по октябрь 2006-2010 гг. дважды в месяц в дневное время на 10-15 станциях с глубинами от 1,5 до 20 м (см. рис. 1 А). Сбор проб осуществлялся икорной сетью с диаметром входного отверстия 56,5 см (ИКС-56,5) и сетным мешком из капронового газа № 14 [12]. Горизонтальные (поверхностные) ловы выполняли на циркуляции судна при средней скорости 1,3 м/с в течение 15 мин. Горизонтальное траление выполняли по стандартной методике [12]. Вертикальных лов осуществлялся в слое 20-0 м, а на участках с глубинами менее 20 м – от дна до поверхности воды. Для горизонтальных обловов количество икры (шт.) и личинок (экз.) выражали на лов, а для вертикальных – на 1 м2 акватории. Более 2 тыс. проб ихтиопланктона фиксировали 4% раствором формалина. Для определения видовой принадлежности икры и личинок использовали определители [13, 14]. Одновременно измеряли температуру воды у поверхности. Пробы грунта для определения в них ртути собирали дночерпателем «Океан» с площадью захвата 0,025 см3 и геологической трубкой ГОИН-1,5. Анализ выполнялся на анализаторе ртути РА 915+ с пиролитической приставкой ПИРО.
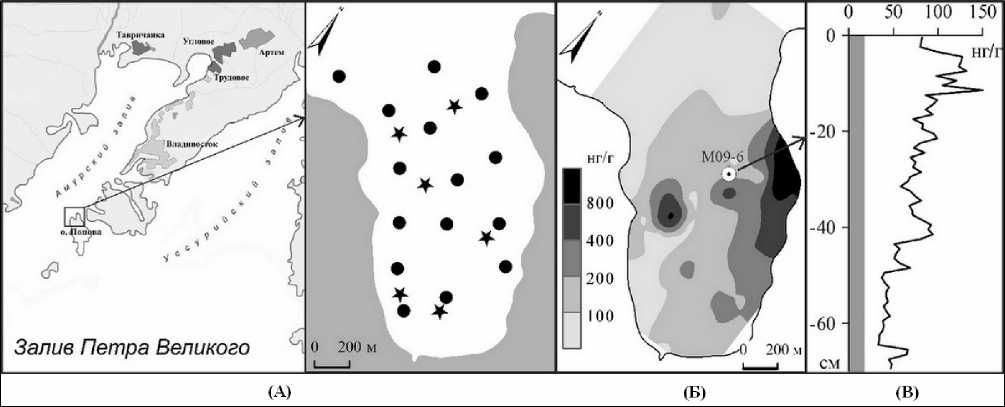
Рис. 1. А – места сбора проб ихтиопланктона (точки) и макробентоса (звездочки); Б – содержание ртути в поверхностном слое (0-5 см) донных осадков; В – содержание ртути в колонке донных осадков М09-6 (серым цветом показан фон для залива Петра Великого)
Результаты и обсуждение. В макробентосе в б. Алексеева обнаружено 128 видов животных и 24 вида макрофитов. Морские травы в бухте представлены Zostera marina и Phyllospadix iwatensis. Заросли зостеры простираются вдоль южного и юго-западного побережья и широкой полосой в кутовой части. Зеленые водоросли Ulva fenestrata и Enteromorpha linza часто встречаются на глубине 0,5-4 м. Среди многочисленных видов бурых водорослей (табл. 1) преобладают Chorda filum и Sar-gassum miyabei. Красные водоросли в основном представлены Corallina pilulifera на участках со скалистыми и каменистыми грунтами в восточной части бухты, на глубине 5-7 м. Виды Neorhodomela larix и Crateloupia divaricata встречаются вдоль северо-восточного и югозападного берегов бухты на глубине от 0,5 до 5-6 м. В юго-западной части бухты на глубине 0,5-1 м на песчано-галечном грунте выделен биоценоз Grateloupia divaricata+Ulva fenestrata+Patiria pectenifera [1]. На этом участке нами отмечен каменистый грунт с наи- лком (частицы от 7 до 10 мм составляют 36 %). Камни покрыты Corallina pilulifera (12-18 %). Наибольшей биомассы достигают офиуры (228,8 г/м2). Высокую численность имеют Polychaeta и Gastropoda (176 экз./м2 и 322 экз./м2 соответственно). Биомасса макробентоса здесь не превышает 930 г/м2, из которой 46,8 г/м2 приходится на Ulva fenestrata.
В южной части бухты между зарослей зостеры на глубине 3-5 м на песчано-илистом грунте (частиц 0,25-0,5 мм – 36 %) располагается полоса Ulva fenestrata. Здесь встречены представители Loricata, Polychaeta, Amphipoda, Izopoda, Nemertini, Ophiuroidea (табл. 2). Офиура Amphiodia fissa образует скопления плотностью до 184 экз./м2. Зеленые водоросли достигают биомассы 1453 г/м2 (Ulva fenestrata) и бурые – 397,5 г/м2 (Dictyota dichotoma). В июле отмечено оседание личинок брюхоногих моллюсков, молодь которых (менее 3 мм) покрывала практически всю поверхность макрофитов. Биомасса и численность Gastropoda здесь составила 477 г/м2 и 6123 экз./м2 соот- ветственно. Наблюдалось также оседание личинок Bivalvia, но их плотность и биомасса были небольшими. В северо-западной части бухты на каменистых грунтах и глубине от 1 до 5-6 м располагается биоценоз Crenomytilus grayanus [1]. Здесь общая биомасса животных составляет 2386,32 г/м2 (из них мидия Грея -1764,97 г/м2), общая плотность 223 экз./м2. Обнаружены Carolina (покрытие 40-50 %), и редкие домики Spirorbidae. Преобладали Loricata
(47 экз./м2 и 44 г/м2), Bivalvia (112 экз./м2 и 16 г/м2) и Polychaeta (48 экз./м2 и 15 г/м2) (табл. 2). Встречены пурпурная и бугорчатая асцидии. Эти животные предпочитают селиться на камнях и скалах в открытой части бухты на глубине от 10 до 17 м. На глубине от 7 до 10 м найдены скопления асцидий (15 экз./м2), а с увеличением глубины они встречаются реже (до 3 экз./м2).
Таблица 1. Видовой состав фитобентоса б. Алексеева
|
Вид |
1 |
2 |
3 |
4 |
5 |
6 |
Вид |
1 |
2 |
3 |
4 |
5 |
6 |
|
Enteromorpha linsa (Chl) |
+ |
+ |
+ |
- |
+ |
- |
Chorda filum (Ph) |
+ |
+ |
+ |
+ |
- |
- |
|
Enteromorpha prolifera (Chl) |
+ |
- |
- |
- |
- |
- |
Costaria costata (Ph) |
- |
+ |
- |
- |
- |
- |
|
Ulva fenestrate (Chl) |
+ |
+ |
- |
+ |
+ |
- |
Laminaria cichorioides (Ph) |
- |
- |
- |
- |
- |
+ |
|
Ulvaria splendens (Chl) |
+ |
- |
- |
- |
- |
- |
Dictyota dichotoma (Ph) |
+ |
+ |
- |
- |
- |
- |
|
Codium fragile (Chl) |
- |
- |
+ |
+ |
+ |
- |
Sargassum miyabei (Ph) |
- |
- |
+ |
+ |
+ |
- |
|
Chaetomorpha melagonium (Chl) |
- |
+ |
- |
- |
- |
- |
Sargassum palidum (Ph) |
- |
- |
+ |
- |
- |
- |
|
Cladophora flexuosa (Chl) |
- |
- |
+ |
- |
- |
- |
Corallina pilulifera (Rh) |
+ |
- |
+ |
- |
+ |
- |
|
Chordaria flagelliformis (Ph) |
- |
- |
+ |
- |
- |
- |
Grateloupia divaricata (Rh) |
+ |
- |
- |
- |
- |
- |
|
Dictyosiphon chordaria (Ph) |
+ |
+ |
- |
- |
- |
- |
Neorhodomela aculeate (Rh) |
+ |
- |
- |
- |
+ |
- |
|
Colpomenia peregeina (Ph) |
+ |
- |
- |
- |
- |
- |
Polysiphonia fibrata (Rh) |
- |
- |
+ |
- |
- |
- |
|
Scytosiphon simplicissimus (Ph) |
- |
+ |
- |
- |
- |
- |
Phyllospadix iwatensis (Emb) |
- |
- |
- |
- |
+ |
- |
|
Desmarestia viridis (Ph) |
- |
- |
+ |
- |
+ |
+ |
Zostera marina (Emb) |
+ |
- |
+ |
+ |
+ |
- |
Таблица 2. Видовой состав зообентоса б. Алексеева
|
Вид |
1 |
2 |
3 |
4 |
5 |
6 |
Вид |
1 |
2 |
3 |
4 |
5 |
6 |
|
Eulalia bilineata (Po) |
- |
+ |
- |
- |
- |
+ |
Neoamphitrite groenlandica (Po) |
- |
- |
- |
- |
+ |
- |
|
Nereiphylla castanea (Po) |
- |
- |
- |
+ |
- |
- |
Pista incarrientis (Po) |
- |
- |
- |
- |
+ |
- |
|
Phyllodoce groenlandica (Po) |
- |
- |
- |
+ |
- |
- |
Lysippe labiata (Po) |
- |
- |
- |
- |
- |
+ |
|
Phyllodoce maculata (Po) |
- |
- |
- |
+ |
+ |
+ |
Melinna elisabethae (Po) |
- |
- |
+ |
- |
- |
- |
|
Pterocirrus macroceros (Po) |
- |
- |
- |
+ |
- |
- |
Pectinaria damai (Po) |
- |
- |
+ |
- |
- |
- |
|
Glycera capitata (Po) |
- |
- |
+ |
+ |
- |
- |
Chone olegi (Po) |
- |
- |
- |
- |
- |
+ |
|
G. chirori (Po) |
- |
- |
- |
- |
- |
+ |
Spirorbis semidentatus (Po) |
+ |
+ |
- |
- |
- |
- |
|
G. onomichiensis (Po) |
- |
- |
+ |
- |
- |
- |
S. nipponicus (Po) |
+ |
- |
- |
- |
- |
- |
|
Glycinde armigera (Po) |
- |
- |
- |
- |
- |
+ |
Circeis nipponicus (Po) |
- |
- |
- |
+ |
- |
- |
|
Goniada maculata (Po) |
- |
- |
- |
- |
- |
+ |
Fabricia crenicolis (Po) |
- |
+ |
- |
- |
+ |
- |
|
Harmothoe imbricata (Po) |
+ |
+ |
+ |
+ |
+ |
- |
Oncoscolex pacificus (Po) |
- |
- |
- |
+ |
- |
- |
|
Lepidonotus squamatus (Po) |
+ |
+ |
- |
- |
- |
- |
Aonides oxycephala (Po) |
- |
- |
- |
+ |
+ |
+ |
|
Nereis tigrina (Po) |
- |
+ |
- |
+ |
- |
Myxicola infundibulum (Po) |
- |
- |
- |
+ |
- |
- |
|
|
N. vexillosa (Po) |
- |
+ |
- |
- |
- |
- |
Thysanocardia nigra (Si) |
- |
- |
+ |
- |
- |
- |
|
N. zonata (Po) |
- |
+ |
+ |
- |
- |
- |
Phascolosoma agasizii (Si) |
- |
- |
+ |
- |
- |
- |
|
Plathynereis bicanaliculata (Po) |
- |
- |
- |
+ |
- |
- |
Balanus crenatus (Ci) |
+ |
- |
+ |
- |
+ |
- |
|
Typosyllis orientalis (Po) |
- |
- |
- |
- |
+ |
- |
Balanus improvisus (Ci) |
- |
- |
- |
- |
+ |
- |
|
Lumbrinereis fragilis (Po) |
- |
- |
+ |
- |
- |
- |
Neomysis sp. (My) |
- |
- |
+ |
- |
- |
- |
|
L. cervicalis (Po) |
- |
- |
+ |
- |
- |
- |
Amphithoe japonica (Am) |
- |
- |
+ |
+ |
- |
- |
|
Scoloplos armiger (Po) |
- |
+ |
- |
- |
- |
+ |
Amphipoda sp. (Am) |
- |
- |
- |
+ |
- |
- |
|
Laonice cirrata (Po) |
- |
- |
- |
- |
- |
+ |
Anisogammarus pugettensis (Am) |
+ |
- |
- |
+ |
- |
- |
|
Cirratulus cirratus (Po) |
- |
- |
- |
- |
- |
+ |
Hyale bassargini (Am) |
+ |
+ |
- |
- |
- |
- |
|
Cirriformia tentaculata (Po) |
- |
- |
- |
+ |
- |
- |
Parhyale zibellina (Am) |
+ |
- |
+ |
+ |
- |
- |
|
Spiophanes uschakovi (Po) |
- |
- |
- |
- |
- |
+ |
Caprella danilewskii (Am) |
- |
+ |
- |
- |
- |
- |
|
Diplocirrus lorgisetosus (Po) |
- |
- |
+ |
- |
- |
- |
Caprella sp. (Am) |
- |
- |
+ |
- |
- |
- |
|
Pherusa plumosa (Po) |
- |
- |
- |
- |
+ |
- |
Cymodoce japonica (Is) |
- |
- |
- |
+ |
- |
- |
|
Ammotrypane multipapilla (Po) |
- |
- |
- |
+ |
+ |
- |
Holotelson tuberculatus (Is) |
- |
- |
- |
+ |
- |
- |
|
Scalibregma inflatum (Po) |
- |
- |
- |
+ |
- |
+ |
Paranthura japonica (Is) |
- |
- |
+ |
- |
- |
- |
|
Axiotella catenata (Po) |
- |
- |
- |
- |
- |
+ |
Pentias hayi (Is) |
- |
+ |
- |
- |
- |
- |
|
Maldane sarsi (Po) |
- |
- |
- |
- |
+ |
+ |
Pachycheles stevensii (De) |
- |
- |
+ |
- |
+ |
- |
|
Pisoides bidentata (De) |
- |
- |
- |
+ |
- |
- |
Modiolus kurilensis (Bi) |
- |
+ |
- |
- |
+ |
- |
|
Pugettia quadridens (De) |
- |
- |
- |
+ |
- |
- |
Mytilus trossulus (Bi) |
+ |
- |
- |
- |
+ |
- |
|
Telmessus cheiragonus (De) |
- |
+ |
- |
- |
- |
- |
Crenomytilus grayanus (Bi) |
- |
- |
+ |
- |
+ |
- |
|
Cancer amphioetus (De) |
- |
- |
- |
- |
- |
+ |
Septifer keenae (Bi) |
- |
- |
- |
+ |
- |
- |
|
Продолжение таблицы 2 |
|||||||||||||
|
Pagurus middendorfii (De) |
+ |
+ |
- |
+ |
- |
- |
Glycymeris yessoensis (Bi) |
- |
- |
+ |
- |
+ |
- |
|
Pagurus sp. (De) |
- |
- |
- |
- |
+ |
- |
Crassostrea gigas (Bi) |
- |
- |
+ |
- |
- |
- |
|
Leptochiton assimilis (Pol) |
- |
- |
- |
+ |
- |
- |
Mizuhopecten yessoensis (Bi) |
- |
- |
- |
+ |
- |
- |
|
Lepidozona albrechti (Pol) |
- |
- |
- |
- |
+ |
- |
Hiatella arctica (Bi) |
+ |
- |
+ |
+ |
- |
- |
|
Tonicella granulata (Pol) |
- |
- |
- |
+ |
- |
- |
Panope japonica (Bi) |
- |
- |
- |
- |
+ |
+ |
|
Tonicella squamigera (Pol) |
+ |
+ |
- |
- |
- |
- |
Keenocardium californiense (Bi) |
- |
+ |
- |
- |
+ |
- |
|
Placiphorella Stimpsoni (Pol) |
+ |
+ |
- |
+ |
- |
- |
Felaniella usta (Bi) |
- |
- |
- |
+ |
+ |
+ |
|
Notoacmea concinna (Ga) |
+ |
+ |
- |
- |
- |
- |
Diplodonta semiosperoides (Bi) |
- |
- |
- |
+ |
- |
- |
|
Collisella heroldi (Ga) |
+ |
- |
- |
- |
- |
- |
Macoma incongrua (Bi) |
- |
- |
- |
+ |
- |
- |
|
Acmaea pallida (Ga) |
- |
+ |
- |
- |
- |
- |
Ruditapes philippinarum (Bi) |
+ |
+ |
- |
+ |
- |
- |
|
Umbonium costatum (Ga) |
- |
- |
- |
+ |
+ |
- |
Protothaca jedoensis (Bi) |
- |
- |
- |
+ |
- |
+ |
|
Homalopoma amussitata (Ga) |
+ |
- |
- |
+ |
+ |
- |
Callithaca adamsi (Bi) |
- |
- |
- |
- |
- |
+ |
|
Homalopoma sanqarense (Ga) |
+ |
- |
+ |
+ |
- |
+ |
Anisocorbula venusta (Bi) |
+ |
+ |
+ |
+ |
+ |
+ |
|
Epheria turrita (Ga) |
+ |
- |
- |
- |
+ |
- |
Mya priapus (Bi) |
- |
- |
- |
- |
+ |
- |
|
Littorina breviculata (Ga) |
- |
+ |
- |
- |
- |
- |
Mya arenaria (Bi) |
- |
- |
- |
- |
- |
+ |
|
Littorina mandshurica (Ga) |
+ |
- |
- |
- |
- |
- |
Apostychopus japonicus (Hol) |
- |
+ |
- |
- |
+ |
- |
|
Littorina squalida (Ga) |
+ |
- |
- |
+ |
- |
- |
Cucumaria fraudatrix (Hol) |
- |
- |
+ |
+ |
+ |
- |
|
Thapsiella plicosa (Ga) |
- |
- |
- |
- |
+ |
- |
Strongylocentrotus nudus (Ech) |
- |
+ |
- |
+ |
+ |
- |
|
Fartulum bucerius (Ga) |
- |
+ |
- |
- |
- |
- |
S. intermedius (Ech) |
+ |
+ |
+ |
+ |
- |
- |
|
Crypthonatica janthostoma (Ga) |
- |
- |
+ |
- |
- |
- |
Patiria pectinifera (Ast) |
+ |
- |
- |
+ |
+ |
- |
|
Alaba vladivostokensis (Ga) |
+ |
- |
- |
- |
- |
- |
Aphelasterias japonica (Ast) |
+ |
- |
- |
- |
+ |
- |
|
Mitrella burchardi (Ga) |
+ |
- |
- |
- |
- |
- |
Lethasterias fusca (Ast) |
- |
- |
- |
- |
+ |
- |
|
Boreotrophon candelabrum (Ga) |
- |
+ |
- |
+ |
- |
- |
Asterias amurensis (Ast) |
- |
- |
- |
+ |
- |
- |
|
Nucella heyseana (Ga) |
+ |
- |
- |
- |
- |
- |
Amphiodia fissa (Oph) |
- |
+ |
- |
- |
- |
- |
|
Bela erosa (Ga) |
- |
- |
- |
- |
- |
+ |
Amphipholis kochii (Oph) |
+ |
- |
- |
+ |
+ |
- |
|
Retusidae sp. (Ga) |
- |
- |
- |
- |
- |
+ |
Amphipholis sp. (Oph) |
- |
- |
- |
- |
+ |
- |
|
Acila insignis (Bi) |
- |
- |
- |
+ |
- |
- |
Ophiura sarsi (Oph) |
- |
- |
- |
- |
+ |
- |
|
Leonucula ovatotruncata (Bi) |
- |
- |
+ |
- |
- |
- |
Halocynthia roretzi (Asc) |
- |
- |
- |
- |
+ |
- |
|
Leonucula tenus tenus (Bi) |
- |
- |
- |
- |
- |
+ |
Halocynthia aurantium (Asc) |
- |
- |
+ |
- |
+ |
- |
|
Vilasina pillula (Bi) |
- |
- |
- |
- |
+ |
- |
Styela clava (Asc) |
+ |
+ |
+ |
+ |
- |
- |
Примечание к таблицам : Chl – Chlorophyta; Ph – Phaeophyta; Rh – Rhodophyta; Emb – Embriobyonti; Po – Polychaeta; Si – Sipunculidae; Ci – Cirripedia; Am – Amphipoda; My – Mysidae; Is – Isopoda; De – Decapoda; Pol – Polyplacophora; Ga – Gastropoda; Bi – Bivalvia; Hol – Holothuroidea; Ech – Echinoidea; Ast – Asteroidea; Oph -Ophiuroidea; Asc – Ascidiacea. 1 – станция 1 (глубина 0,5-1 м); 2 – станция 2 (глубина 4 м); 3 – станция 3 (свал, глубина 5 м); 4 – станция 4 (глубина 6 м); 5 – станция 5 (глубина 8 м); 6 – станция 6 (глубина 15 м).
В кутовой части бухты на глубине 5 м отмечен галечный грунт с примесью крупнозернистого песка (частиц 3-1 мм – 41%). Наибольшая биомасса у двустворчатых моллюсков (659 г/м2). Встречены сипункулиды, полихеты, асцидии (табл. 2). Плотность амфипод была низкой (0,2 экз./м2). В поясе Zostera marina на глубине 1-8 м грунт представлен песком (частиц 0,5-1 мм – 42%) с примесью гальки. В этом районе ранее располагался биоценоз Zostera marina+Stichopus japonicus+Mizuhopecten yes-soensis [1]. Здесь, в поясе зостеры, нами отмечена самая высокая биомасса морской травы (1285,61 г/м2). Биомасса бурых водорослей была меньше ( Sargassum miyabei – 6,75 г/м2 и Chorda filum – 1,32 г/м2). На этой станции преобладали Bivalvia (764 экз./м2 и 981,32 г/м2). Биомасса офиур не превышала 0,12 г/м2. У входной части бухты на глубине более 15 м на илах развит биоценоз Maldane sarsi с молодью брюхоногих и двустворчатых моллюсков (плотностью 3778 экз./м2 и биомассой до 100,05 г/м2).
На экосистему б. Алексеева оказывает влияние ртутное загрязнение, выявленное в 1986 г., тогда концентрации ртути в тканях моллюсков значительно превышали фоновые [6]. В 1990 и 1991 гг. максимальные концентрации ртути обнаружены у модиолуса, наименьшие – у приморского гребешка, однако у последнего были разрушены раковины и снижен прирост. Морфологические аномалии моллюсков характерны для районов с высокими концентрациями ртути в донных осадках [6]. В 2003 и 2004 гг. изучено распределение ртути в поверхностном слое донных осадков (см. рис. 1 Б, В). Обнаружены аномалии в северо-западной и восточной частях бухты. Минимальная концентрация ртути составила 50 нг/г, что в два раза выше геохимического фона зал. Петра Великого [15]. Концентрация ртути в аномальных зонах превышала 800 нг/г, достигая 2500 нг/г. Уровень концентрации ртути в донных осадках наиболее загрязненных акваторий зал. Петра Великого (б. Золотой Рог, устье Второй Речки) составляет 1500-3000 нг/г [16, 17]. В 2009 г. в б. Алексеева отобрана колонка донных осадков (М09-6), в верхней части которой обнаружено увеличение концентрации ртути в два раза по сравнению с нижней. Возможно, обогащение ртутью верхней части колонки связано с антропогенным влиянием. В тоже время в нижней части уровень концентрации ртути был 50 нг/г, что выше фона залива в два раза [16].
Ихтиопланктон б. Алексеева представлен 27 видами рыб из 15 семейств (табл. 2), из которых наибольшее видовое разнообразие у семейств Pleuronectidae и Stichaeidae (по 5 видов). Промысловое значение имеют сельдь C. pallasii , навага Eleginus gracilis , камбалы (Pleu-ronectidae), малоротая корюшка H. japonicus , зубастая корюшка Osmerus mordax и анчоус Engraulis japonicus . Встречены также молодь других видов рыб – постоянных обитателей прибрежного мелководья. Икра и личинки всех видов были подразделены на пространственноэкологические группы, характеризующиеся разной стратегией нереста [17]. Критериями объединения видов в группы служила приуроченность их к водам определенной солености в период нереста [18] и использование определенного нерестового субстрата [19].
Группа I включает солоноватые виды, нерест и жизненный цикл которых проходит на прибрежном мелководье, в пределах литоральной и сублиторальной зоны в условиях изменяющейся солености. Эта группа в ихтио-планктоне была представлена 8 видами ( Liop-setta pinnifasciata, Pholidapus dybowskii, Opist-hocentrus ocellatus, O. zonope, Acanthogobius flavimanus, Chaenogobius heptacanthus, Pallas-ina barbata, Pseudopleuronectes obscurus ), или 30% от общего числа видов.
Группа II объединяет виды, обитающие в морских водах, которые для нереста заходят в реки, устья, приустьевые участки рек и прибрежное мелководье. Этой категории встречено 10 видов (37%). В эту группу входят проходные ( O. mordax ), полупроходные ( H. ja-ponicus , Gasterosteus aculeatus ) и морские ( C. pallasii, E. gracilis , Pseudopleuronectes yoko-hamae , Syngnathus acusimilis и 3 вида из семейства Hexagrammidae) виды, выметывающие пелагическую, демерсальную (литофиты и псаммофилы) или прикрепленную к морским травам и водорослям (фитофилы) икру. Некоторые из них проявляют заботу о потомстве (напр. трехиглая колюшка, формирующая нерестовые гнезда, охраняющие кладки терпуги, а также живородящая рыба-игла).
Группа III включает виды, нерест которых проходит в глубоководных районах заливов, в водах с морской соленостью, а на прибрежное мелководье икра и личинки заносятся течениями. В б. Алексеева отмечены икра и личинки 9 видов (33%) этой группы. Они подразделены на две подгруппы. К первой относятся виды рыб, откладывающие демерсальную икру (Neozoarces steindachneri, Stichaeus nozavae), личинки которых в ихтиопланктоне встречались эпизодически. Во вторую подгруппу включены виды, выметывающие пелагическую икру, такие как дальний мигрант анчоус, и рыбы, совершающие сезонные миграции (Limanda aspera, Hippoglossoides herzen-steini, Hypoptychus dybowskii). Основной нерест рыб этой группы проходит на глубине 15-20 м, однако их икра встречается и в прибрежной зоне.
Наибольшее видовое разнообразие ихти-опланктона отмечено в апреле-мае. Летом преобладали икра и личинки пелагофильных видов, представителей морской и полупроходной экологических групп, основной нерест которых протекает в июне-июле. Пелагическая икра рыб, нерестящихся летом в период наибольшего распреснения вод, имеют меньший диаметр, чем у весенненерестящихся видов, их икра снабжена дополнительными жировыми каплями [18], что позволяете ей удерживаться в поверхностном слое воды [20]. К августу практически все виды рыб завершали нерест. Только в конце сентября и начале октября появлялись личинки рыб сем. Hexagrammidae. С начала июня до начала сентября в планктоне встречались личинки асцидий, так как в этот период происходит нерест Halocynthia auran-thium . В июне 2007 г. плотность личинок Halocynthia auranthium составила 0,35 экз./м3. Пик нереста асцидий был в июле, и плотность личинок находилась в пределах 1,0-1,2 экз./м3, а в августе снизилась до 0,45 экз./м3. В начале сентября личинки асцидий встречались единично, что связано с завершением нереста. Личинки Halocynthia auranthium имели длину 2,53,5, в среднем 3,0 мм. В 2008 г., плотность личинок Halocynthia составляла в июне – 1,1 экз./м3, в июле увеличилась до 2,0 экз./м3, в августе снижалась до 1,05 экз./м3. Длина личинок составляла 2,0-3,7 мм, в среднем 2,5 мм. Таким образом, нерест асцидий происходил в летние месяцы и его пик был в июле, что подтверждает данные из литературных источников [21, 22].
В ихтиопланктоне б. Алексеева очень часто присутствуют мертвые или с аномалиями икринки рыб. Так, количество и доля икры анчоуса с нормально развивающимися эмбрионами, возрастали по направлению от прибрежного мелководья к более открытым районам бухты, что также связано с распространением распресненных вод. Высокий процент нормально развивающейся икры отмечался в поверхностных водах с соленостью более
32‰. В районах, где соленость была менее 20‰, доминировали икринки с деформированными эмбрионами. Нерест анчоуса протекал при температуре воды от 9 до 24°С. В нерестовый сезон выживаемость икры возрастала по мере повышения температуры воды. Наибольшая смертность икры отмечена при переходе с I на II этап развития. Это объясняется тем, что при развитии эмбрион наиболее чувствителен к воздействию внешних факторов на начальных этапах развития. Гибель икры рыб связана в основном с факторами ветрового волнения, изменениями температуры и солености воды, а также с содержанием ртути в поверхностном слое донных осадков. Ртуть попадает в воды б. Алексеева из природных источников. Через бухту проходит разломная зона, явным признаком которой является абразионный уступ на м. Низкий, сложенный глиной. Вдоль этого уступа концентрация ртути в воздухе превышает фоновый уровень для о. Попова в 30 раз [23].
Выводы: в макробентосе в б. Алексеева обнаружено 128 видов животных и 24 вида макрофитов. На скалистых и каменистых грунтах большая биомасса складывается за счет мидии Грея. На мягких илистых грунтах преобладают многощетинковые черви. На песчано-галечном грунте в поясе зостеры наблюдалось наибольшее видовое разнообразие макробентоса. В ихтиопланктоне отмечены личинки 27 видов рыб из 15 семейств. Наибольшее видовое разнообразие ихтиопланктона отмечено в апреле и мае. Численность ихтиопланктона зависит от многих антропогенных и абиотических факторов. Высокие концентрации ртути в донных осадках бухты (50 нг/г) обусловлены как антропогенными, так и природными источниками.
Работа выполнена при финансовой поддержке грантов ДВО РАН 11-III-В-07-143, 11-III-В-07-133, 11-III-В-07-007, 11-III-B-07-047, 11-III-B-07-006.
Список литературы Экологический мониторинг морской биоты бухты Алексеева (залив Петра Великого)
- Волова, Г.Н. Бентос бухты Алексеева (залив Петра Великого)/Г.Н. Волова, Т.И. Жакина, Л.В. Микулич//Прибрежный планктон и бентос северной части Японского моря. -Владивосток: ДВНЦ АН СССР. 1980. С. 32-56.
- Гомелюк, В.Е. Ихтиофауна бухты Алексеева острова Попова (залив Петра Великого, Японское море) и влияние на нее культивирования приморского гребешка/В.Е. Гомелюк, С.Л. Кондрашев, А.В. Левин//Биология шельфовых и проходных рыб. -Владивосток: ДВО АН СССР. 1990. С. 5-8.
- Завертанова, Ю.В. Ихтиопланктон бухты Алексеева (зал. Петра Великого, Японское море) в 2006-2007 гг.//Вопр. рыб. 2010. Т. 11, № 1 (41). С. 38-47.
- Завертанова, Ю.В. Особенности формирования видового состава ихтиопланктона и его взаимосвязь с гидрологическим режимом бухты Алексеева 2006-2007 гг.//Исследования Мирового океана: мат-лы междунар. науч. конф. 26-30 мая 2008 г. -Владивосток: Дальрыбвтуз, 2008. С. 38-41.
- Федорец, Ю.В. Влияние абиотических факторов на выживание икры из северо-восточной части Амурского залива//Проблемы экологии морского шельфа: мат. всерос. науч. молод. конф.-шк. Владивосток 16-22 сент. 2010 г. -Владивосток: Изд-во ДФУ, 2010. С. 175-178.
- Лучшева, Л.Н. Содержание ртути в компонентах экосистемы бухты Алексеева (залив Петра Великого Японского моря)//Биол. моря. 1995. Т. 21. № 6. С. 412-415.
- Ушаков, П.В. Многощетинковые черви дальневосточных морей СССР. -Л.: Зоологический институт, 1955. 445 с.
- Перестенко, Л.П. Красные водоросли дальневосточных морей России. -СПб.: Ольга, 1994. 331 с.
- Виноградова, К.Л. Определитель водорослей дальневосточных морей СССР. Зеленые водоросли. -Л.: Наука, 1979. 147 с.
- Адрианов, А.В. Таксономический каталог биоты залива Петра Великого Японского моря/А.В. Адрианов, О.Г. Кусакин. -Владивосток: Дальнаука, 1998. 350 с.
- Парсонс, Т.Р. Биологическая океанография/Т.Р. Парсонс, М. Такахаши, Б. Харгрейв. -М.: Легкая и пищевая промышленность, 1982. 432 с.
- Расс, Т.С. Методическое руководство по сбору икринок, личинок и мальков рыб/Т.С. Расс, И.И. Казанова. -М.: Пищевая промышленность, 1966. 35 с.
- Перцева-Остроумова, Т.А. Определительные таблицы пелагической икры рыб залива Петра Великого//Изв. ТИНРО. 1955. Т. 43. С. 43-68.
- Okiyama, M. (ed.). Atlas of the early stage fishes in Japan. Tokyo: Tokai Univ. Press. 1988. 1154 p.
- Аксентов, К.И. Антропогенное загрязнение ртутью донных осадков зал. Петра Великого/К.И. Аксентов, А.С. Астахов//Вест. ДВО РАН. 2009. № 4. С. 115-121.
- Поляков, Д.М. Содержание ртути в компонентах экосистемы залива Петра Великого/Д.М. Поляков, М. Вирцавс, С.И. Козлова и др.//Водные ресурсы. 1991. № 5. С. 101-108.
- Давыдова, С.В. Видовой состав ихтиопланктона бухт залива Петра Великого и его сезонная динамика//Изв. ТИНРО. 1998. Т. 123. С. 105-121.
- Шубников, Д.А. Прибрежно-эстуарное сообщество рыб северной части Индийского океана и экологические связи его компонентов//Вопросы ихтиологии. 1977. Вып. 5. С. 824-830.
- Крыжановский, С.Г. Эколого-морфологические закономерности развития карповых, вьюновых и сомовых рыб//Тр. Ин-та морфол. животных АН СССР. 1949. Вып. 1. С. 237-329.
- Подорванова, А.Ф. Основные черты гидрохимии залива Петра Великого (Японское море)/А.Ф. Подорванова, Т.С. Ивашинникова, В.С. Петренко, Л.С. Хомичук. -Владивосток: ДВО АН СССР, 1989. 201 с.
- Саватеева, Л.Ю. Дальневосточные голотурии и асцидии как ценное пищевое сырье/Л.Ю. Саватеева, М.Г. Маслова, В.Л. Володарская. -Владивосток: Изд-во Дальневост. ун-та, 1983. 184 с.
- Косьяненко, А.А. Экология асцидий рода Halocynthia в б. Алексеева//Современное состояние водных биоресурсов: Мат-лы науч. конф., посв. 70-летию С.М. Коновалова. -Владивосток: ТИНРО-Центр, 2008. С. 566-567.
- Колесник, О.Н. Геоэкологический мониторинг содержания ртути в компонентах морских и прибрежный экосистем Южного Приморья (на примере о. Попова)/О.Н. Колесник, А.Н. Колесник, К.И. Аксентов//Геология морей и океанов: Мат-лы XVII междун. науч. конф. (школы) по морской геологии 12-16 нояб. 2007 г. Москва. -М.: ГЕОС, 2007. С. 237-239.
