Экологическое разнообразие лесного населения жужелиц (Coleoptera, Carabidae) и его изменение в условиях вырубки и дальнейшего лесовозобновления
 и его изменение в условиях вырубки и дальнейшего лесовозобновления")
Автор: Анциферов Анатолий Леонидович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 1, 2017 года.
Бесплатный доступ
В работе рассматривается динамика экологической структуры населения жужелиц при переходе от одного местообитания к другому в контексте резких экологических нарушений при лесозаготовке и дальнейшего лесовозобновления по градиенту давности вырубки. Выявлены закономерности, моделирующие процесс формирования сообщества жужелиц в условиях ранней стадии вторичной сукцессии на обнаженном участке леса и дальнейших тенденций с использованием наиболее простых показателей разнообразия, распределения обилий видов и меры доминирования.
Лесной биотоп, вырубка, лесовозобновление, жужелицы, видовое богатство, обилие, экологический состав, переформирование
Короткий адрес: https://sciup.org/146116711
IDR: 146116711 | УДК: 574.34
Ecological diversity of forest populations of ground beetles (Coleoptera, Carabidae) and its change under logging and reforestation
We discuss the dynamics of the ecological structure of the beetle population during the transition from one habitat to another in the context of sharp ecological disturbances during logging and further reforestation. By using the simplest indicators of diversity, the distribution of species and the measure of dominance we revealed the certain trends modelling the formation of the beetle community in the conditions of the early stage of secondary succession on the exposed forest area and following sussessive communities.
Текст научной статьи Экологическое разнообразие лесного населения жужелиц (Coleoptera, Carabidae) и его изменение в условиях вырубки и дальнейшего лесовозобновления
Введение. Одним из наилучших примеров, иллюстрирующих степень изменения компонентов видового разнообразия и обилия, являются антропогенные нарушения, в частности такого рода, как сплошное сведение древостоя при лесозаготовках. Проблема массовой вырубки леса и ее влияния на структуру экосистем особенно остро стоит для регионов с промышленным запасом лесных ресурсов и, как следствие, развивающимся приоритетом лесозаготовительной отрасли. По этой причине изучение структуры экологического разнообразия и закономерностей его трансформации при лесных вырубках и последующих процессах вторичной сукцессии видится, несомненно, актуальным и значимым.
Оценка разнообразия наиболее информативна и легка для интерпретации, когда применяется к четко ограниченным и строго определенным таксономическим категориям. В данном случае в качестве модельной группы выбрано семейство жуков-жужелиц (Coleoptera, Carabidae). Жужелицы являются наиболее удобным объектом для изучения процессов формирования сообществ и реструктуризации в экосистемах под влиянием антропогенных и естественных причин, так как они многочисленны, повсеместно распространены, легко отлавливаются и учитываются и, что особо важно, чувствительны к разным по характеру экологическим нарушениям.
Цель данной работы состоит в оценке динамики экологического разнообразия, включающего компоненты видового богатства и численности населения жужелиц, а так же их зоогеографической, экологической и фенологической структуры при переходе от одного местообитания к другому в контексте резких экологических нарушений при вырубке древостоя и дальнейшего лесовозобновления по градиенту давности вырубки.
Материал и методика. Сбор фактологического материала производился с 30 апреля по 30 сентября 2014 года в окрестностях д. Лазарево Судиславского района Костромской области. В качестве объекта исследований был выбран лесной биотоп с двумя прилегающими сплошными вырубками возрастов до 1 года и 4-х лет. Такое расположение разновозрастных вырубок по отношению друг к другу и исходному участку дает возможность вполне достоверно изучить смену разных стадий вторичной сукцессии одного и того же биотопа в один полевой сезон. Выбранный лесной массив характерен для южно-таежной зоны, к которой относится большая часть территории Костромской области.
На основании лесорастительного описания данный биотоп охарактеризован как ельник сложный (далее исходный лес). Для него характерно присутствие таких древесных пород, как ель европейская (Picea abies L.), сосна обыкновенная (Pinus silvestris L.), береза бородавчатая (Betula verrucosa Ehrh.), осина (Populus tremula L.). Подлесок представлен черемухой кистевидной (Padus racemosa С.К. Schneid) и рябиной обыкновенной (Sorbus aucuparia L.). Ель и береза в данном типе леса составляют также незначительную долю подроста. Среди кустарников наибольшей плотностью выделяются смородина черная (Ribes nigrum L.) и жимолость обыкновенная (Lonicera xylosteum L.). Травяной покров не богат видами, основная доля приходится на щитовник мужской (Dryopteris filixmas Schott.) и кислицу (Oxalis acetosella L.). Данный участок леса имеет рыхлый суглинистый гумусовый слой почвы, с достаточно мощной (6-7 см) подстилкой.
Сплошная вырубка возрастом менее 1 года (далее, однолетняя вырубка) примыкает к исходному участку леса. Лесозаготовка на данном участке завершена в зимний период того же года в котором производились настоящие учеты жужелиц. В травостое преобладает щитовник мужской (Dryopteris filix-mas Schott.), чертополох курчавый (Carduus crispus L.), пикульник (Caleopsis speciosa Mill.) и некоторые злаковые: пырей ползучий (Triticum repens L.), вейник наземный (Calamagrostis epigeios Rotch.,) мятлик (Роа trivialis L.) и др. Поверхность территории вырубки неровная, подстилка сохранилась частично, почва приобрела более сухой и плотный характер.
Вырубка 4-летнего возраста (далее 4-летняя вырубка) примыкает к однолетней вырубке и исходному участку леса. Данная площадка отличается густым возобновлением осины, березы и ольхи серой (Alnus incana L.). В среднем ярусе господствует малина (Rubus idaeus L.). В травостое отмечена высокая плотность кипрея узколистного (Epilobium angustifolium L.) и злаков. Давность рубки определялась по возрасту поросли от пня березы (Мелехов и др., 1965). '
Для определения видового состава и динамической плотности популяций жужелиц был использован метод почвенных ловушек. Благодаря этому методу можно получить наиболее полное представление о видовом составе и численности почвенных жуков исследуемых биотопов. В качестве фиксирующей жидкости применялся раствор поваренной соли близкий к насыщенному. Ловушки устанавливались в каждом биотопе леса и вырубок равномерно в линейные трансекты по 15 шт. Расстояние между ловушками в пределах одной трансекты составляло около 10 м. За единицу выборки принималась совокупность материала из ловушек каждой трансекты участка леса и каждой вырубки, взятая единовременно. Собираемый материал каждой трансекты суммировался. Выборки производились одновременно со всех трансект в среднем через каждые 10 суток. Всего за сезон исследований было сделано по 12 выборок в каждом типе площадок.
В анализе видового богатства использован наиболее простой -индекс разнообразия Маргалефа (DMg):
DMg = (S-l)/LnN, где S - число выявленных видов, N - общее число особей всех S видов. Данный показатель учитывает как число видов, так их относительное обилие, что делает ее наиболее реальной основой для изучения видового разнообразия (Мэгарран, 1992).
В качестве меры доминирования использованы показатели, основанные на относительном обилии видов, учитывающие одновременно и выровненность и видовое богатство - индекс Симпсона (D) (Simpson, 1949):
D = Z pi2, где pi - доля особей i-ro вида, а также индекс Бергера-Паркера (d) (Berger, Parker, 1970):
d = Nmax/N, где Nmax - число особей самого обильного вида.
Классификация жужелиц по обилию проводилась по системе О. Ренконена (Renkonen, 1938). Виды, доля которых превышает 5% от общего сбора в отдельном биотопе, составляют группу доминантов; находится в пределах 2-5% - группу субдоминантов; составляет менее 2% - группу редких. Данная шкала наиболее объективна в оценке соотношения видов в биотопе и отмечает ее широкое применение отечественными исследователями (Грюнталь, 2008).
Определение биотопического преферендума видов производилось с использованием данных С.Ю. Грюнталя (2008), разделение видов на группы жизненных форм имаго проведено согласно системе, разработанной И.Х. Шаровой (1981). Определение жужелиц осуществлялось по таблицам О.Л. Крыжановского (1986) и А.Ю. Исаева (2002).
В общей сложности за период исследований на указанных участках отловлено и учтено 5739 особей жужелиц 48 видов 20 родов. В исходном лесу отмечено 2849 особей 28 видов, на однолетней вырубке - 1679 особей 41 вида, на 4-летней вырубке -1211 особей 36 видов.
Структура фауны и населения жужелиц исходного участка леса. Характеристика видового состава исходного участка леса (видовое богатство, обилие и доля за весь сезон) приведена в таблице 1. Общий список видового состава жужелиц исследованного лесного массива представлен 28 видами 16 родов. Среди них, по анализу фауны, на первом месте находятся транспалеарктические виды (42,9%), что отличается от данных при анализе населения (преобладают европейско-сибирские -51,2%).
Видовое богатство жужелиц исходного участка леса по индексу Маргалефа (DMg) составляет 3,4.
По суммарному обилию особей (за весь период исследований) в нем преобладают: Trechus secalis Pz. - 1424 экз., что составляет 50,0% от общей массы жужелиц лесного биотопа, Pterostichus oblongopunctatus F. - 589 экз. (20,7%), Carabus nemoralis Mull. - 297 экз. (10,4%) и Pterostichus niger Schaller. - 249 экз. (8,7%). По классификации О. Ренконена эти виды отнесены к категории доминантов, так как доля каждого из них превышает 5% от общей массы. Суммарная доля доминантных видов жужелиц в лесу составила 89,8% от общей массы жужелиц при достаточно малом количестве видов (4), представляющих эту группу. В число субдоминантов лесного участка вошел единственный вид - Platynus assimilis Pk., доля которого составила 3,7% от общего числа жужелиц. Расчет меры доминирования в сообществе жужелиц исходного участка леса по индексу Симпсона (D) и Бергера-Паркера (d) показывает степень выровненности видового состава в значениях 3,2 и 2 соответственно.
Таблица 1
Видовое богатство, обилие и доля жужелиц в исходном участке леса
|
Исходный участок леса |
Однолетняя вырубка |
4-летняя вырубка |
||||
|
Вид |
Доля, % |
Вид |
Доля, % |
Вид |
Доля, % |
|
|
S Д S S О |
Trechus secalis Pz. |
50,0 |
Trechus secalis Pz. |
26,1 |
Trechus secalis Pz. |
49,1 |
|
Pterostichus oblongopunctatus F. |
20,7 |
Pterostichus oblongopunctatus F. |
18,6 |
Poecilus versicolor Sturm. |
11,6 |
|
|
Carabus nemoralis Mull. |
10,4 |
Pterostichus niger Schall. |
12,4 |
Pterostichus niger Schall. |
6,4 |
|
|
Pterostichus niger Schall. |
8,7 |
Poecilus versicolor Sturm. |
6,4 |
Pterostichus strenuus Pz. |
6,2 |
|
|
Poecilus cupreus L. |
5,7 |
|||||
|
Carabus nemoralis Mull. |
5,4 |
|||||
|
S О |
Platynus assimilis Pk. |
3,7 |
Carabus granulatus L. |
4,7 |
Amara aenea Deg. |
3,9 |
|
Pterostichus strenuus Pz. |
2,4 |
Pterostichus oblongopunctatus F. |
3,5 |
|||
|
P. melanarius Ill. |
2,0 |
P. melanarius Ill. |
2,8 |
|||
|
Carabus granulatus L. |
2,2 |
|||||
|
Редкие |
23 вида |
8,2 |
31 вид |
16,0 |
30 видов |
17,1 |
|
Видовое богатство |
28 |
41 |
36 |
|||
|
Обилие, экз. |
2849 |
1679 |
1211 |
|||
Характер видового состава жужелиц данного биотопа сопоставим с аналогичным типом леса (ельником сложным с преобладанием хвойных пород), обследованным ранее (в 2005 году) в той же географической местности (Анциферов, Смирнов, 2010). Особое различие заключается в высоком значении доли Carabus nemoralis Mull, на участке ельника 2014 года - 20,7%, тогда как в ельнике сложном 2005 года этот вид редок (0,2%). Так как Carabus nemoralis является чутким индикатором, увеличивающим свою численность с ростом фактора рекреации (Грюнталь, Бутовский, 1997; Грюнталь, 2008), это характеризует исследуемый участок леса как территорию с повышенной рекреационной нагрузкой.
По биотопическому преферендуму основу комплекса жужелиц исходного участка леса составляют характерные лесные виды - (15 видов - 96,6% от общего обилия). Из них самые массовые - Trechus secalis Pz. (50,0%), Pterostichus oblongopunctatus F. 20,7%), Carabus nemoralis Mull. (10,4%), Pterostichus niger Schaller. (8,7%) и Platynus assimilis Pk (3,7%). Лесоболотные жужелицы представлены 5 видами (2,3%), из которых самый многочисленный - Pterostichus strenuus Pz. (1,5%). Из других групп в этом биотопе присутствуют лугово-полевые (2 вида), полевые (1 вид), прибрежные (1 вид) и эврибионты (3 вида) с суммарным обилием - 1,1% от общей численности (табл. 2).
Таблица 2
Состав и обилие группировок жужелиц по биотопическому преферендуму в исходном участке леса, однолетней и 4-летней вырубках
|
Исходный участок |
Однолетняя вырубка |
4-летняя вырубка |
|
|
Лесные |
96,6 |
73,4 |
65,7 |
|
Лесоболотные |
2,3 |
3,3 |
9,3 |
|
Лугово-полевые |
0,08 |
16,4 |
19,1 |
|
Луговые |
1,8 |
0,3 |
|
|
Полевые |
0,04 |
0,4 |
1,8 |
|
Прибрежные |
0,2 |
0,4 |
0,6 |
|
Эврибионты |
0,7 |
4,2 |
3,7 |
По фенологическому признаку структура фауны жужелиц отличается от структуры населения. Для фауны жужелиц исходного лесного участка характерно преобладание видов с весенним размножением -16 видов из 28, тогда как в населении ведущая роль принадлежит летне-осенним видам - 51,3% от общей массы. Основу всего обилия летне-осенних видов составляет Trechus secalis Pz. -50,0%.
Сведения о структуре фауны и населения, а так же биотопической приуроченности жужелиц исследуемого лесного биотопа во многом согласуются с аналогичными данными, приведенными для разных типов ельников Костромской (Еремин, 1986), Московской (Грюнталь, 1983, 2008) областей и Марийской АССР (Арнольди, Матвеев, 1973). Это указывает на типичность выявленного состава жужелиц для данного типа леса, а перечисленные доминантные виды можно назвать его количественными индикаторами.
В комплексе жизненных форм имаго жужелиц исходного участка леса отмечено абсолютное господство зоофагов (99,4%), что является характерной чертой населения лесов подзоны южной тайги таежной зоны (Грюнталь, 2008). Преобладают представители трех групп: подстилочные стратобионты, подстилочно-почвенные стратобионты и эпигеобионты ходящие (табл. 3). Среди первых наиболее обилен Trechus secalis Pz., (50,0%) и он же единственный доминант. В составе подстилочно-почвенных стратобионтов доминируют Pterostichus oblongopunctatus F. (20,7%) и Pterostichus niger Schall. (8,7%), а эпигеобионтов ходящих крупных - только Carabas nemoralis Mull. (10,4%).
Таблица 3
Структура комплекса жизненных форм жужелиц исходного участка леса
|
Жизненные формы |
Доля, % |
||
|
Исходный участок |
1 -летняя вырубка |
4-летняя вырубка |
|
|
Зоофаги в том числе: |
99,4 |
95,9 |
93,2 |
|
подстилочные стратобионты |
53,8 |
31,7 |
60,8 |
|
подстилочно-почвенные стратобионты |
30,0 |
33,6 |
12,9 |
|
поверхностно-подстилочные стратобионты |
4,0 |
17,2 |
14,9 |
|
эпигеобионты ходящие крупные |
11,6 |
12,3 |
3,8 |
|
эпигеобионты летающие |
- |
0,7 |
0,1 |
|
геобионты роющие |
- |
- |
0,7 |
|
Миксофитофаги в том числе: |
0,6 |
4,1 |
6,8 |
|
геохортобионты гарпалоидные |
0,5 |
2,3 |
1,6 |
|
стратохортобионты |
0,04 |
0,4 |
1,1 |
|
стратобионты скважинки |
0,04 |
1,4 |
4,3 |
Таким образом, анализ населения жужелиц характеризует исходный участка леса (ельник сложный), как биотоп с преобладанием широко распространенных лесных зоофагов с летне-осенним размножением, населяющие преимущественно подстилку (подстилочные стратобионты) и верхние горизонты почвы (подстилочно-почвенные стратобионты).
Структура фауны и населения жужелиц однолетней вырубки в сравнении с исходным участком леса. Анализ структуры сообщества жужелиц показывает резкую смену фауны и населения в первом же году существования вырубки. Число видов на однолетней вырубке не падает, а напротив, увеличивается с 28 до 41 вида 18 родов. Суммарное обилие жужелиц при этом снизилось с 2849 до 1679 особей. Среди них большинство видов (34,1%) относятся к европейско-сибирскому типу ареала, тогда как в отношении населения европейско-сибирские виды уступают транспалеарктическим, хотя и незначительно - 39,0 и 40,3% соответственно. Снижение обилия европейско-сибирских видов на вырубке в первый год обусловлено, прежде всего, значительным сокращением численности Trechus secalis Pz. с 1424 до 439 особи (с 50 до 21,1% соответственно). Однако данный вид по-прежнему возглавляет перечень доминирующих видов наряду с Pterostichus oblongopunctatus F. (18,6%), Pterostichus niger Schall. (12,4%), Carabus nemoralis Mull. (5,4%) но с дополнением новых видов-вселенцев: Poecilus versicolor Sturm. (6,4%) и Poecilus cupreus L. (5,7%). Доминирующий в лесном участке Carabus nemoralis Mull, и субдоминант Platynus assimilis Pk. утрачивают свою значимость на вырубке первого года. Общая доля доминантных видов однолетней вырубки снизилась по отношению к исходному участку (89,8%) до 74,6%. Комплекс субдоминантов на данной вырубке формируют Carabus granulatus L. (4,7%), Pterostichus strenuus Pz. (2,4%) и P. melanarius Ill. (2,0%), их общее обилие составляет 9,1% от всей численности (табл. 1).
По индексу разнообразия Маргалефа (DMg) показатель видового богатства жужелиц увеличился с 3,4 в исходном участке до 5,39 на вырубке в первый год. Уровень неоднородности видового состава по индексам Симпсона (D) и Бергера-Паркера (d), описывающих распределение обилий видов, возрос до значений 7,7 и 3,84 соответственно (рис. 1).
Данные показатели отражают характер изменений в структуре лесного населения жужелиц. Очевидно, что следствием сплошной вырубки древостоя в первый год является увеличение разнообразия сообщества, выражающееся в росте видового богатства в сочетании с фактором выровненности, т.е. равномерности распределения обилий видов этого семейства. Высокая выровненность, когда обилие видов равное или почти равное, условно считается эквивалентной высокому разнообразию (Мэгарран, 1992).
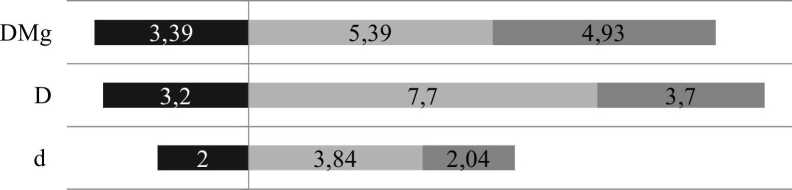
■ Исходный участок Однолетняя вьтпубка
Рис. 1 . Изменение значений индексов Маргалефа (DMg), Симпсона (D) и Бергера-Паркера (d) от исходного участка леса к вырубкам по градиенту их давности
Резкое изменение среды обитания для тенелюбивых лесных видов жужелиц на вырубке приводит к заметному снижению их значимости в отношении остальных групп преферендума. Процентная доля характерных лесных видов на однолетней вырубке по-прежнему высока, но снизилась по отношению к исходному участку (95,6%) и составила 73,4% от остального населения жужелиц (табл. 3). Число видов, составляющих эту группу, изменяется не значительно, но резко уменьшаются в обилии фоновые жужелицы, такие как Trechus secalis Pz., Pterostichus oblongopunctatus F., Carabus nemoralis Mull., Platynus assimilis Рк. Одновременно на вырубке существенно возросло значение лугово-полевых жужелиц, обилие которых составило 16,4% в составе 9 видов. В массе появляются новые виды-вселенцы: Poecilus versicolor Sturm. (6,4%), Р. cupreus L. (5,7%), Bembidion assimile Gyll. (1,8%). Среди мигрантов однолетней вырубки отмечены, в том числе 4 вида луговых жужелиц: Agonum sexpunctatum L., A. gracile Sturm., Amara familiaris Duft. и A. eurynota Pz., в общей сложности, составляющие 1,8%. Нарастает в численности и группа эврибионтов -их доля увеличилась с 0,7% в исходном участке до 4,2% на месте вырубки. Значение лесоболотных видов выросло с 2,3 до 3,3%.
В обилии фенологических групп для фауны жужелиц однолетней вырубки характерно господство весенних видов по отношению к исходному участку леса - 56,3% и 39,1% соответственно. Тогда как летне-осенние виды, преобладавшие в лесном массиве, резко снизили свою значимость на вырубке - с 51,3% до 29,0% (рис.2). Такое изменение обусловлено существенным снижением численности Trechus secalis Pz., обилие которого составляет основу всего комплекса летне-осенних жужелиц. В группе жужелиц с весенним типом размножения доминирует Pterostichus oblongopunctatus F. (18,6%), хотя и с меньшей численностью по отношению к исходному лесу.
|
Весенние |
56,3 |
38 |
||||
|
Летне-осенние |
29 |
52,3 |
||||
|
Мультисезонные L |
14,|б |
|||||
|
■ Исходный участок |
□ Однолетняя вырубка □ 4-летняя вырубка |
|||||
Рис. 2 . Изменение обилия (в %) фенологических групп жужелиц от исходного участка леса к однолетней вырубке
Высоких значений достигают так же и весенние виды жужелиц - вселенцев однолетней вырубки, такие как Poecilus versicolor Sturm. (6,4%) и Р. cupreus L. (5,7%). В связи с этим, можно предположить, что изменение почвенно-растительных и микроклиматических условий в первый год после вырубки влияет на формирование фенологического состава фауны и населения жужелиц, в результате которого участок вырубки в первый год более привлекателен для весенних фенологических групп и в меньшей степени летне-осенних.
В комплексе жизненных форм имаго жужелиц на однолетней вырубке продолжает господствовать класс зоофагов - 95,9% (табл. 3), состав которого пополняется группой эпигеобионтов летающих (0,7%), представленных одним видом - Cicindela campestris L. В общем составе групп жизненных форм жужелиц сплошная лесосека привела к характерным изменениям. Существенно снизилась доля видов, населяющих преимущественно подстилку (подстилочных стратобионтов), что, безусловно, является показателем степени нарушения лесного подстилочного горизонта в процессе лесозаготовки. Виды, населяющие верхние горизонты почвы (подстилочно-почвенные стратобионты) большой чувствительности к вырубке не проявили, и их население составляет здесь большинство -33,6%. Поверхностно-подстилочные стратобионты напротив, отреагировали резким ростом обилия - с 4,0% в исходном участке до 17,2% на вырубке. Среди них наиболее значимы, Poecilus versicolor Sturm. (6,4%) и Р. cupreus L. (5,7%), Bembidion assimile Gyll. Ha численность крупных ходящих эпигеобионтов, к которым относятся Carabus nemoralis Mull, С. granulatus L., C. cancellatus 111. и др., вырубка леса так же, заметного воздействия не оказала.
Более благоприятно в условиях однолетней вырубки чувствуют себя представители класса миксофитофагов, включающие фитофагов со смешанным питанием. Общая доля их группового состава возросла с 0,6 до 4,1% от всего сообщества жужелиц. Из них геохортобионты гарпалоидные (преимущественно Harpalus laevipes Zett.) представляют большинство - 2,3%.
В итоге необходимо отметить, что формирующееся в первый год на территории вырубки сообщество жужелиц преимущественно состоит из широко распространенных лесных зоофагов, обитающих в верхних горизонтах почвы (подстилочно-почвенные стратобионты), размножающиеся главным образом весной.
Резкая смена населения жужелиц в первом году, быстрое и массовое появление лугово-полевых Poecilus versicolor Sturm., Р. cupreus L. и других связано, несомненно, с миграцией, которая совершается частично путем перелета {Bembidion, Amara). Базой для существования для таких видов и многих других более тепло- и светолюбивых жужелиц является связь с культурными землями, а так же рассеянные в самих ельниках разреженные участки - редины, гари, окна в пологе и другие, более давние лесосеки, в данном случае -примыкающая 4-летняя вырубка.
Структура фауны и населения жужелиц 4-летней вырубки в сравнении с участком однолетней. К периоду четырехлетней давности на вырубке происходит смена растительных условий, характерная для процесса заболачивания, сопровождающаяся формированием густого травостоя с преобладанием кипрея и примесью осоки, свето- и влаголюбивых пород кустарников и деревьев. В свою очередь происходит и трансформация сообщества герпетобионтной мезофауны, отражающаяся на признаках, составляющих ее экологическое разнообразие.
В целом, по истечению 4-летнего периода видовое богатство жужелиц на вырубке снизилось с 41 до 36 видов 16 родов. Общее обилие при этом продолжает снижаться, и по отношению к однолетней вырубке уменьшилось с 1679 до 1211 особи (табл. 1). Среди них как по фауне, так и по населению доминируют виды с европейско-сибирским ареалом - 33,3% и 67,8% соответственно. Подобное колебание величин обилия европейско-сибирских видов от исходного участка к однолетней и далее к 4-летней вырубке соотносится с перепадами обилия в основном одного из представителей данной зоогеографической группы - Trechus secalis Pz. При этом независимо от изменения своей численности на разных площадках, данный вид остается преобладающим.
В отношении остального доминантно-субдоминантного комплекса жужелиц участка 4-летней вырубки следует отметить характерную реструктуризацию. В частности, лесной вид Pterostichus oblongopunctatus F. сократив численность с 18,6% до 3,5%, перешел в категорию субдоминантов. Его место занял лугово-полевой Poecilus versicolor Sturm., доля которого возросла с 6,4% до 11,6%. Лесоболотный Pterostichus strenuus Pz. из разряда субдоминантов (2,4%) поднялся до уровня доминантов, увеличив свою значимость до 6,4%. В свою очередь, к числу субдоминантов 4-летней вырубки присоединился лугово-полевой вид Amara aenea Deg. (3,9%).
Четырехлетние изменения экологического разнообразия на лесной вырубке отражено в расчетах показателей Маргалефа (DMg), Симпсона (D) и Бергера-Паркера (d). На рисунке 1 отражено снижение параметров, включающих соотношение видового богатства, обилия и степени выровненности обилий на 4-летней вырубке по отношению к однолетней. При этом видовое богатство (DMg) уменьшилось не столь явно - с 5,39 до 4,93, тогда как уровень выровненности обилий по Симпсону (D) и Бергеру-Паркеру (d) сократился практически в два раза. Причина подобной динамики объясняется вышеизложенным сопоставлением доминантной структуры двух разновозрастных вырубок, где отмечается резкое колебание обилия Trechus secalis Pz., влияющее и на динамику рассматриваемых показателей.
Структура экологических групп по преферендуму на 4-летней вырубке отличается от таковой на однолетней лесосеке. Отмечается продолжение тенденции снижения доли лесных видов (с 73,4 до 65,7%) и увеличения значимости лесоболотных, лугово-полевых и полевых (табл. 2). При этом основной фон во всем сообществе жужелиц по-прежнему сохраняют лесные виды.
Условия 4-летней вырубки оказались менее благоприятны для жужелиц с весенним типом размножения (рис.2). Их доля снизилась с 56,3 до 38% за счет существенного сокращения обилия таких видов как Pterostichus oblongopunctatus F. (с 18,6 до 3,5%), Poecilus cupreus L. (с 5,7 до 0,9%), Carabus nemoralis Mull, (c 5,4 до 1,2%), C. granulatus L. (c 4,7 до 2,2%), C. cancellatus 111. (c 1,9 до 0,3%). Доля летне-осенних жужелиц напротив, возросла — с 29 до 52,3% по причине отмеченного ранее роста численности Trechus secalis Pz.
Спектр жизненных форм жужелиц 4-летней вырубки характеризуется наибольшим разнообразием - 9 групп, что по отношению к исходному участку и однолетней вырубке больше на 2 и 1 соответственно (табл. 3). Отмечено появление еще и группы геобионтов роющих в лице единственного представителя - Clivina fossor L. За счет вышеуказанного роста обилия Trechus secalis Pz. общая доля подстилочных стратобионтов увеличилась 31,7 до 60,8% от остального состава. Одновременно произошло заметное, в той или иной степени сокращение всех остальных групп класса зоофагов. Наиболее значительно это проявилось у группы стратобионтов подстилочно-почвенных, прежде всего за счет таких видов как Poecilus cupreus L., Bembidion assimile Gyll., Agonum sexpunctatum L., сокративших свою численность. Кроме того присутствие Platynus assimilis Pk., на исходном участке и на однолетней вырубке было значительным (3,7 и 1,0% соответственно), но на 4-летней вырубке уже не отмечено вовсе. В общем обилии класса миксофитофагов видна тенденция роста значимости по градиенту давности вырубки (табл. 3). Их доля увеличилась с 4,1% на однолетней вырубке до 6,8% на 4летней, что характеризует их как показателей меняющихся растительных условий на вырубке. Среди наиболее характерных индикаторов стоит выделить Harpalus rufipes Deg. и Amara aenea Deg., отвечающих на данные изменения увеличением доли присутствия.
Таким образом, по структуре населения жужелиц, к четвертому году давности территория вырубки характеризуется как биотоп с преобладанием широко распространенных лесных зоофагов с летнеосенним размножением, населяющих преимущественно подстилку (подстилочные стратобионты).
Заключение. Итогом данного исследования стало выявление особенностей формирования фауны и населения жужелиц в первый год после сведения древостоя, что соответствует фазе обнажения и миграции, т. е. появлению незаселенного пространства и заселению первыми пионерными видами (Clements, 1916) и дальнейшие тенденции в экологической структуре жужелиц на фоне меняющихся условий микроклимата. Видовое разнообразие и выровненность обилий жужелиц на свежей вырубке значительно больше, чем в исходном лесу, что положительно сказывается на экологической устойчивости сообщества. Это может быть объяснено тем, что территория вырубки активно заселяется из соседних биотопов. В пользу этого утверждения говорит тот факт, что многие жужелицы, обитающие в лесу и на вырубке, имеют высокую численность. Наличие на однолетней вырубке видов жужелиц, не встречающихся в исходном участке леса, говорит о том, что прилегающий лес - не единственный биотоп, из которого идет заселение вырубки жуками. В ближайшие годы показатели экологического разнообразия вырубки начинают снижаться, о чем свидетельствуют данные по 4-летней вырубке.
Однако этим влияние условий вырубки на переформирование сообщества жужелиц не ограничивается. Микроклимат вырубки первого года сказывается на соотношении фенологических групп. Ее территория, при этом, становится более благоприятной для видов с весенним сроком размножения, в отличие от исходного лесного биотопа, где преобладают летне-осенние жуки. Характерно, что к 4летнему возрасту соотношение этих типов размножения изменяется в сторону, более близкую к исходному состоянию. Топическое влияние леса на однолетнюю вырубку остается высоким на протяжение всего четырехлетнего периода лесовозобновления. Характерные лесные виды жужелиц сохраняют господство как на однолетней, так и на 4летней вырубках. Однако имеет место тенденция нарастания значимости лесоболотных, лугово-полевых и полевых видов жужелиц.
Фоновым классом жизненных форм на трех сопоставляемых биотопах являются хищники, но по градиенту давности вырубки увеличивается доля миксофитофагов. В ходе данных исследований выявлен комплекс видов жужелиц, специфически реагирующих на определенные факторы изменяющихся экологических условий в ходе лесовозобновления, что подтверждает их высокое биоиндикационное значение.
Об авторах:
Анциферов А.Л. Экологическое разнообразие лесного населения жужелиц (Coleoptera, Carabidae) и его изменение в условиях вырубки и дальнейшего лесовозобновления / А.Л. Анциферов // Вести. ТвГУ. Сер.: Биология и экология. 2017. № 1. С. 86-99. '
Список литературы Экологическое разнообразие лесного населения жужелиц (Coleoptera, Carabidae) и его изменение в условиях вырубки и дальнейшего лесовозобновления
- Арнольди К.В., Матвеев В.А. 1973. Население жужелиц (Coleoptera, Carabidae) еловых лесов у южного предела тайги (Марийская АССР)//Экология почвенных беспозвоночных. М.: Наука. С. 131-143.
- Анциферов А.Л., Смирнов И.В. 2010. Видовой состав жужелиц (Coleoptera, Carabidae) лесного массива Костромской области: структура фауны леса и участка вырубки//Вестн. ТвГУ. Сер.: Биология и экология. Вып. 27, № 23. С. 34-45.
- Грюнталь С.Ю. 1983. Комплексы жужелиц (Coleoptera, Carabidae) в лесах подзоны широколиственно-еловых лесов//Фауна и экология почвенных беспозвоночных Московской области. М.: Наука. С. 85-99.
- Грюнталь С.Ю. 2008. Организация сообществ жужелиц (Coleoptera, Carabidae) лесов Восточно-Европейской равнины. М.: Галлея-Принт. 463 с.
- Грюнталь С.Ю., Бутовский Р.О. 1997. Жужелицы (Coleoptera, Carabidae) как индикаторы рекреационного воздействия на лесные экосистемы//Энтомологическое обозрение. Т. 76. № 3. С. 547-554.
- Еремин П.К. 1986. Население жужелиц (Coleoptera, Carabidae) девственных еловых лесов южной тайги//Кологривский лес. М.: Наука. С. 110-115.
- Мелехов И.С., Корконосова Л.И., Чертовской В.Г. 1965. Руководство по изучению типов концентрированных вырубок. М.: Наука. 172 с.
- Мэгарран Э. 1992. Экологическое разнообразие и его измерение. М.: Мир. 161 с.
- Шарова И.Х. Жизненные формы жужелиц. 1981. М.: Наука. 293 с.
- Berger W.H., Parker F.L. 1970. Diversity of planktonic Foraminifera in deep sea sediments. Science, 168. 1345-1347.
- Clements F.E. 1916. Plant Succession: An Analysis of Vegetation. Washington D.C.: Carnegie Institution of Washington. 512 p.
- Renkonen O. 1938. Statisch-ökologische Untersuchungen über die terrestrische Käferwelt der finnischen Bruchmoore//Ann. Zool. Soc. Zool. Bot. Fenn. Vanamo. Bd. 6. 231 s.
- Simpson E.N. 1949. Measurement of diversity//Nature. V. 163. 688 p.
