Экология близкородственных видов мухоловок рода Ficedula в местах их симбиотопии

Автор: Иванов А.Е.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 251 т.13, 2004 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149828
IDR: 140149828
Ecology of two closely related species of the genus Ficedula at symbiotopy
Текст статьи Экология близкородственных видов мухоловок рода Ficedula в местах их симбиотопии
Сравнительно-экологические исследования близких видов птиц в условиях их симбиотопии занимают важное место в экологии, систематике и филогении. Особую актуальность эти вопросы приобретают в связи с синан-тропизацией и урбанизацией авифауны. Удобными модельными видами для подобных популяционных исследований служат два вида пёстрых мухоловок — белошейка Ficedula albicollis и пеструшка Ficedula hypoleuca в местах их совместного обитания. Биология этих видов изучена не одинаково. Если мухоловке-пеструшке посвящено очень много работ (Благосклонов 1948; Мальчевский 1959; Лихачёв 1966; Зимин 1972; Анорова 1976, 1977, 1984; Иноземцев 1976, 1978, 1987; Артемьев 1981, 1983; Бардин и др. 1987; Прокофьева 1994; Нумеров 1995; Семёнов 2003; Шутова 2003; и мн. др.), то бе-лошейке почти не уделялось внимания. Наиболее известные исследования по экологии этого вида выполнены в Воронежской области более 40 лет назад (Кадочников 1957, 1960; Королькова 1954, 1963). Однако они не ставили задачей специальное сравнение F. albicollis и F. hypoleuca.
За последние полвека в центре Русской равнины в экологии мухоловок произошли заметные изменения: у них отмечены синантропные тенденции, расширение ареалов, увеличение численности. Во многих местах пеструшка и белошейка начали гнездиться бок о бок. Так, в 1920-е годы для Воронежской области считалось, что мухоловка-пеструшка представляет большую редкость (Благосклонов 1954; Кадочников 1957, 1960). В Савальском лесу в 1930-1940-е она практически не встречалась. В настоящее время она является здесь обычным видом, а гнёзд белошейки стало в четыре раза больше, чем раньше. Другой пример —Тульская область, где белошейка была редка до конца 1950-х (Швец 1998); в 1960-е произошло заметное увеличение её численности в Тульских засеках, в заповеднике “Ясная Поляна” и других местах, где она стала фоновым видом. Отмечено гнездование белошейки и в черте города Тулы. Сходные изменения происходят и в других частях ареалов этих видов. Причины этого пока не ясны. В связи с этим актуально изучение сравнительной экологии и поведения мухоловок пеструшки и белошейки при гнездовании на одной и той же территории.
Материалы и методика
Экологию белошейки и пеструшки изучали в 2002-2003 годах на многолетнем орнитологическом стационаре в Савальском лесу (Воронежская обл.). Проводили целенаправленные наблюдения и сравнения экологии этих видов в местах их сов- местного обитания для выявления конкурентных отношений, возможных механизмов разделения ресурсов и расхождения по экологическим нишам. В задачи исследований входило изучение особенностей гнездования, социального, полового и трофического поведения мухоловок, питания взрослых птиц и птенцов.
В 2002 г. изучали экологию гнездования, анализировали особенности размещения гнёзд в естественных дуплах, гнездовые территории этих двух видов, корм птенцов, кормовое поведение. В 2003 г. были также развешены искусственные гнездовья. Особенности гнездовых территорий птиц описывали по общепринятым схемам. Кормовое поведение птиц анализировали путём “непрерывных регистраций поведения” (Боголюбов, Преображенская 1991) в предгнездовой период. В таблицы заносили участки крон деревьев, где кормились птицы (всего 1234 регистрации). Проводили наблюдения и за кормлением птенцов.
Пробы птенцового корма получали методом наложением шейных лигатур (Мальчевский, Кадочников 1953). Лигатуры изготавливали из лески толщиной 0.1-0.15 мм. Корм изымали у птенцов в возрасте от 3 до 10 сут. Рассчитывали количество и долю встречаемости представителей каждого семейства от общего числа пищевых объектов, потребляемых каждым видом. Проведены расчёты сходства (коэффициент корреляции) состава пищи.
Ширину экологических ниш вычисляли по общепринятой формуле Симпсона (Мегарран 1992). Степень перекрывания ниш рассчитывали по формуле Пианки (Pianka 1973).
Результаты и обсуждение
Особенности гнездования
Савальский лес представляет искусственный лесной массив в степи общей площадью 7 000 га. Первые посадки деревьев сделаны в 1877 г. Ныне основу древесной растительности составляют посадки дуба Quercus robur и сосны Pinus sylvestris разного возраста, в основном 40-60 лет. Более подробно растительность описана нами ранее (Иванов 2003).
В Савальском лесу обитают все виды европейских мухоловок, из них массовым видом является белошейка. Численность пеструшки, малой Fice-dula parva и серой Muscicapa striata мухоловок в 3-4 раза ниже, чем бело-шейки, а в некоторых местах различия в численности еще больше. В Савальском лесу белошейка и пеструшка гнездятся повсеместно. Особый интерес представляют те участки леса, где гнёзда этих видов находятся в непосредственной близости друг от друга.
В обследованных участках леса гнёзда белошеек располагались на высоте 4-12, чаще 4.5-8 м. Хотя много дупел располагалось ниже 4 м (около 45%), белошейки предпочитали им более высоко расположенные дупла. Пеструшки селились в среднем на высоте 4-6 м. Во всех случаях максимальное число гнёзд приходилось на участки спелого дубового или смешанного леса, где было много естественных дупел.
Для привлечения мухоловок в 2003 г. было развешено 100 искусственных гнездовий (88 синичников и 12 “уголков”) на расстоянии 15-25 м друг от друга. Развешивание проводили на 4 участках на разных видах деревьев (на сосне 62, на вязе 15, на дубе 9 , на клёне 1, на ясене 1). Из 88 домиков было занято 45, из них 31 белошейкой, 7 пеструшкой, 4 большой синицей Parus major и 3 гнезда не определено. Заселяемость по участкам колебалась от 35 до 84%. Максимум приходился на зрелые и хорошо осветлённые уча- стки со слабым подлеском, минимум отмечен на открытом участке без подлеска и подроста.
Пространственное распределение птиц во время сбора корма
Изучали распределение птиц во время поиска ими корма в древеснокустарниковой растительности. Регистрировали положение птицы в разных частях крон деревьев (930 регистраций по белошейке и 304 по пеструшке). Белошейки ловили насекомых: на первичных ветвях — 0.3%, на вторичных ветвях — 6.9%, на третичных — 44.45%, на четвертичных — 27.8% и на концевых — 14.1%. Ширина пространственной ниши белошейки в 2002 г. была равна 0.51, в 2003 г. — 0.43 (Табл. 1). Высотные зоны птицы использовали следующим образом: 10-15 м — 43.2%, 15-20 м — 44.3%, 5-10 м — 14.5%. Зона 0-5 м ими не посещалась. Пеструшка более полно использовала пространство в кронах. На основных ветвях сделано 0.9% регистраций, на толстых ветвях — 6.27%, на “скелетных” ветвях крон — 33%, на тонких ветвях — 40.2%, на концевых — 19.5%. Более широко пеструшка использовала и высотные зоны: 0-5 м — 9.9%, 5-10 м — 45.53%, 10-15 м — 33.1%, 15-20 м — 11.6%. Ширина пространственной ниши пеструшки в 2002 г. была равна 0.62, а в 2003 г. — 0.52. Перекрывание пространственных ниш обоих видов по всему объёму экологического пространства равно 0.64 в 2002 г. и 0.49 в 2003 г. (Табл. 2).
Опираясь на результаты двухлетних исследований, можно сделать предположение о том, что выбранные модельные виды собирают корм в разных участках крон деревьев. Их пространственные ниши различаются, несмотря на заметное перекрывание.
Таблица 1. Ширина пространственной ниши мухоловок
|
Виды |
2002 год |
2003 год |
|
Ficedula albicollis |
0.51 |
0.43 |
|
Ficedula hypoleuca |
0.62 |
0.52 |
Таблица 2. Перекрывание пространственных ниш мухоловок
F. hypoleuca, 2002 F. albicollis, 2003
|
Ficedula albicollis, 2002 |
0.64 |
— |
|
Ficedula hypoleuca, 2003 |
— |
0.49 |
Территориальное поведение
На модельных площадках гнездилась 31 пара белошеек и 7 пар пеструшек. Наблюдение проводили за теми участками, где гнезда этих видов располагались на расстоянии 15-20 м друг от друга.
Наибольшее количество контактов особей этих двух видов приходилось на гнездовой и предгнездовой периоды. Столкновения наблюдали как между самцами белошейки и пеструшки, так и между самцами белошейки. Особой разницы между ними в период столкновений заметить не удалось. Обычно хозяин территории, на которую вторгался пришелец, прогонял конкурирующего самца за пределы своих владений. Несколько раз наблюдали, как самец белошейки заглядывал в дупло, занятое пеструшкой, и каждый раз его выгоняли. Чаще конкурирующие самцы демонстративно перемещались вдоль границ своих участков, активно сигнализируя пением о том, что территория занята.
Величина кладки и размеры яиц
Величина кладок и размеры яиц у пёстрых мухоловок хорошо изучены (Haartman 1951; Лихачёв 1961, 1966, 1978; Головань 1982; Пекло 1987; Зимин 1988; Климов 2003; и др.). Всеми исследователями отмечено значительное сходство белошейки и пеструшки по этим показателям. Однако материалов для сравнения не достаточно.
Таблица 3. Размеры яиц Ficedula albicollis albicollis (Temminck, 1815)
|
Источник |
п |
Длина, мм |
Диаметр, мм |
||||
|
min |
max |
М ± т |
min |
max |
М+т |
||
|
Савальский лес |
167(28) |
17.29 |
21.0 |
17.72±0.07 |
12.98 |
"15.4 |
13.24±0.03 |
|
(наши данные, 2003) Саратовская область (Завьялов и др. 1996) |
17.4 |
20.0 |
17.97±0.10 |
13.0 |
13.8 |
13.39±0.05 |
|
|
Европейская часть России (Пекло 1987) |
55(11) |
15.3 |
20.7 |
17.8±0.14 |
12.1 |
14.0 |
13.2±0.07 |
|
Украина (Марисова, Холина 1959) |
78(18) |
16.7 |
17.9 |
17.5 |
12.8 |
14.0 |
13.4 |
|
Европейская Часть России (Благосклонов 1954) |
14 |
16.5 |
17.8 |
17.3 |
12.0 |
13.2 |
12.85 |
|
Западная Европа (Makatsch 1976) |
30 |
16.9 |
19.4 |
18.21 |
13.0 |
14.3 |
13.66 |
|
Примечание: число перед скобками — количество промеренных яиц, |
|||||||
|
число в скобках — количество кладок, из которых они взяты. |
|||||||
|
Таблица 4. Размеры яиц Ficedula hypoleuca hypoleuca (Pallas, 1764) |
|||||||
|
Источник |
п |
Длина, мм |
Диаметр, мм |
||||
|
min |
max |
М + т |
min |
max |
М+т |
||
|
Савальский лес (наши данные, 2003) |
44(7) |
16.1 |
19.5 |
17.78±0.07 |
12.50 |
14.0 |
13.34±0.025 |
|
Европейская часть России (Пекло 1987) |
158(25) |
15.9 |
19.5 |
17.7±0.05 |
12.30 |
14.4 |
13.3±0.03 |
|
Европейская часть России (Благосклонов 1954) |
100 |
16.00 |
19.5 |
17.93 |
12.10 |
14.4 |
13.42 |
|
Подмосковье (Егорова, |
538 |
16.60 |
19.7 |
18.08 ± |
12.50 |
14.1 |
13.44±0.5 |
|
Константинов 2003) |
0.14 |
0 |
|||||
|
Западная Европа (Makatsch 1976) |
268 |
16.1 |
20.3 |
17.80 |
12.4 |
14.3 |
13.39 |
Примечание: число перед скобками — количество промеренных яиц, число в скобках — количество кладок, из которых они взяты.
Наши наблюдения 2003 года позволили получить следующие сведения. Из 31 полной кладки белошейки 2 содержали по 8 яиц, 12 — 7 яиц, 12 — 6 яиц, 3 — 5 яиц и 2 — 4 яйца. В одной повторной кладке было 3 яйца. Из 7 кладок пеструшки 3 кладки содержали по 6 яиц, 3 — 7 яиц и 1 — 5 яиц.
Размеры яиц белошейки и пеструшки приведены в таблицах 3 и 4 в сравнении с некоторыми литературными данными.
Размеры яиц у изучаемых видов очень схожи. При сравнении данных по Савальскому лесу можно заметить, что яйца белошеек несколько мельче. Но для изучения ооморфологических признаков важны не только сами линейные показатели, но и степень их изменчивости. Результаты последних исследований С.М. Климова (2003) показали существование заметной разницы в степени изменчивости морфологических признаков яиц у птиц. Он анализировал длину, диаметр, индекс формы яиц многих видов, в т.ч. и мухоловок. Оказалось, что у белошейки наблюдается более высокая изменчивость по всем перечисленным показателям, чем у пеструшки.
Питание и выкармливание птенцов
В 2002 г. питание птенцов белошейки изучали в 12 гнёздах (собрано 50 кормовых объектов), пеструшки — в 2 гнёздах (33 объекта). У белошейки преобладали взрослые насекомые (хрущик садовый 48%, таракан лесной 14%, бабочки пяденицы и цикадки по 8%). У пеструшки преобладали: хрущик садовый (21%), пауки (21%), личинки цикадок (15%), муравьи (9%). В 2003 г. собран более полный материал. Под наблюдением находились 7 гнёзд белошейки (31 проба, 127 пищевых объектов) и 7 гнёзд пеструшки (42 пробы, 104 объекта). Корм изымали практически одновременно в гнёздах, находящихся в непосредственной близости друг от друга. В состав пищи птенцов белошейки входили представители 21 семейства из 5 отрядов (см. рисунок), причём доминировали личинки (76.4 %), среди которых гусеницы составляли 60.6%, личинки равнокрылых 14.2%, далее следовали имаго жесткокрылых — 11%. Среди пищи птенцов пеструшек обнаружены представители 24 семейств из 6 отрядов. Преобладали имаго (75%), из них перепончатокрылые составили 27.8%, пауки — 14.4%, двукрылые — 12.5%, жуки 8.6%. Среди личинок преобладали гусеницы (14.4% от общего числа объектов). Значения мер ширины экологической ниши, перекрывания экологических ниш и сходства состава пищи птенцов белошейки и пеструшки приведены в таблицах 5, 6 и 7.
Таблица 5. Ширина трофической ниши мухоловок
|
Виды |
2002 год |
2003 год |
|
Ficedula albicollis |
0.31 |
0.13 |
|
Ficedula hypoleuca |
0.35 |
0.45 |
Таблица 6. Перекрывание трофических ниш мухоловок
F. hypoleuca, 2002 F. albicollis, 2003
|
Ficedula albicollis, 2002 |
0.55 |
— |
|
Ficedula hypoleuca, 2003 |
— |
0.188 |
Таблица 7. Сходство состава пищи двух видов мухоловок (коэффициент корреляции)
F. hypoleuca, 2002 F. albicollis, 2003
|
+0.43 |
— |
|
— |
-0.0153 |
Ficedula albicollis, 2002
Ficedula hypoleuca, 2003
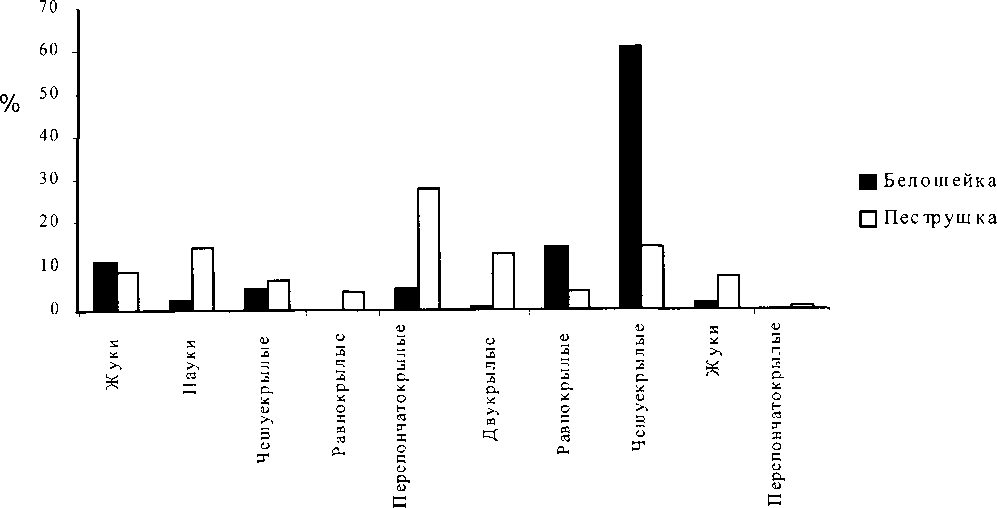
Рис. 1
Питание белошейки и пеструшки в 2003 г.
Анализируя собранные материалы можно отметить, что видовой набор пищевых объектов мухоловок белошейки и пеструшки схож, хотя у каждого вида есть свои предпочтения. В пищевом рационе отмечено большое разнообразие членистоногих при относительно высокой численности отдельных видов, которые были обильны и легко доступны. Это, видимо, приводило к переключению белошеек на одни виды насекомых, а пеструшек — на другие. На этом основании можно сделать предположение о существовании различий в рационах питания птенцов. И хотя данные за разные годы несколько различались, трофические ниши мухоловок чётко расходились. Благодаря этому птицы двух видов селятся в непосредственной близости друг от друга и в сходных местообитаниях, возможно, избегая конкуренции. По мнению некоторых авторов, избыточность необходимых пищевых ресурсов может приводить к снижению конкуренции даже при высокой степени перекрывания трофических ниш (Лэк 1957; Джиллер 1988). Именно это могло происходить в нашем случае.
Выкармливание птенцов
В кормлении птенцов у белошейки и пеструшки принимают участие оба родителя. “Рабочий” день мухоловок начинается с 4 ч 30 мин и продолжается до 20 ч 30 мин. В среднем белошейки приносили корм в гнездо 27 раз в час, а пеструшки — 25. Между утренними и вечерними часами особой разницы в частоте приноса корма птенцам не обнаружено. Самки белошейки приносили корм чаще самцов, а у пеструшки родители приблизительно в равной мере участвовали в выкармливании. В одном случае, когда у самца белошейки было две самочки, птенцов кормили только они. Самец лишь изредка появлялся у обоих гнёзд, даже не заглядывая в них.
Успешность выкармливания птенцов — хороший показатель приспособленности птиц к условиям среды. Однако в разные годы доля погибших птенцов в гнёздах птиц может меняться. Это зависит как от возрастных особенностей родителей, так и, в большей степени, от конкретных условий места и сезона. В 2003 г. при наблюдении за 22 гнездами белошейки мы установили, что в 3 кладках успешность выкармливания составила 100%, в 3 выводках птенцы полностью погибли. В среднем успешность выкармливания птенцов составила 68.1%.
Выводы
-
1. Мухоловка-белошейка и мухоловка-пеструшка в Савальском лесу во время поисков корма по-разному используют одни и те же участки крон. Белошейки в меньшей степени, чем пеструшки осваивали кроны. В результате показатели пространственных ниш двух видов заметно различались.
-
2. У изучаемых видов схожи элементы видового территориального поведения и выкармливания птенцов. Различия по этим показателям между видами минимальные.
-
3. Размеры яиц белошейки и пеструшки отличаются незначительно, что свидетельствует об одинаковых размерах самок и близком родстве птиц. Более существенные различия обнаруживаются при сравнении изменчивости оометрических показателей, таких как длина, ширина и индекс формы.
-
4. Питание птенцов изучаемых видов различается. Белошейка и пеструшка, по нашему мнению, в это время практически не конкурируют за пищевые ресурсы, что позволяет им гнездиться в экологически сходных местообитаниях.
