Экология даурской пищухи на северной периферии ареала

Автор: Борисова Наталья Геннадьевна, Старков Алексей Иннокентьевич, Галиева Галина Рашидовна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
У даурской пищухи, обитающей в степях Юго-Западного Забайкалья, изучены количественные параметры вырываемых ею нор и их устройство. В норах поддерживаются температура и влажность, отличные от таковых окружающей среды. Суточная активность в жаркое время года носит бимодальный характер с пиками в утренние и вечерние часы.
Даурская пищуха, юго-западное забайкалье, устройство нор, суточная активность
Короткий адрес: https://sciup.org/146116532
IDR: 146116532 | УДК: 59(571.54)
Ecology of Daurian pika on the northern periphery of the species range
Morphology and quantitative parameters of Daurian pikas’ burrows in the steppes of South-Western Transbaikalia were studied. Temperature and humidity in the burrows are different from those outside. Daily activity of pika in hot season is bimodal with morning and evening peaks.
Текст научной статьи Экология даурской пищухи на северной периферии ареала
Введение. Степи - один из наиболее уязвимых биомов в настоящее время. Состояние степей Юго-Западного Забайкалья оценивается на сегодняшний день как относительно благополучное. Сохранение такого статуса требует грамотного управления, основанного на твердых научных знаниях структуры и процессов функционирования экосистем. В функционировании определяющая роль принадлежит ключевым видам, каковым за счет высоких потенциала размножения, кормовой, запасающей и роющей активностей для экосистем Юго-Западного Забайкалья является даурская пищуха (Ochotona dauurica Pallas, 1776). В Юго-Западном Забайкалье, где проходит северная граница ареала вида, особенности экологии даурской пищухи не исследованы. В настоящей работе приводятся результаты изучения строения нор и суточной активности зверьков в регионе. Знание этих экологических характеристик позволяет не только определить место вида в экосистеме, но и понять пределы климатической ниши вида, выявить размах приспособляемости вида к климатическим условиям среды.
Методика. При раскопках нор (N=24) зарисовывали ходы в вертикальной и горизонтальной проекциях, измеряли длину, диаметр, количество ходов, углы между ходами, глубину залегания колен, глубину расположения и диаметр камер. Измерения проводили с помощью рулетки или метровой ленты (точность измерения 1 см), измерение диаметра хода и камер в норе - штангенциркулем, углы - с
Вестник ТвГУ. Серия "Биология и экология". 2014. № 4 помощью транспортира.
Для изучения режимов температуры и влажности размещали на глубине 10 см датчики iButton DS1923-F5 в жилых норах, в нежилых и вне нор. На поверхности эти данные измеряли с помощью метеостанции Skywatch GEOS Nil.
Суточную активность регистрировали в ходе наблюдений за индивидуально распознаваемыми зверьками.
Результаты. Для формализации описания мы выделяли в системе подземных ходов наиболее протяженную ось, имеющую на обоих концах выходы на поверхность. Таковых в норах даурской пищухи оказалось не более 2. К оси (осям) под разными углами как в вертикальной, так и в горизонтальной плоскости примыкают более короткие тоннели, называемые нами боковыми ходами, или коленами; количество их колеблется от 10 до 20. Глубина залегания ходов различна. В каштановых почвах делювиального шлейфа максимальная глубина ходов в среднем составляет 0,90 м, редко - до 1 м и более. В средней части каменистых склонов пищухи закладывают ходы не глубже 40 см. Общая длина подземной системы ходов колеблется от 1,79 до 9,15 м. Даже при значительном количестве колен длина магистрального (-ых) ходов превосходит длину всех вместе взятых боковых тоннелей. Глубина наклона ходов нор варьирует в широких пределах: от 10-20° до 60-70° в некоторых участках нор. Угол наклона хода в магистральных ходах в среднем составляет 35° в норах как на склонах, так и в понижениях, но если на склонах крайние значения слабо отличимы от среднего значения, то в понижениях разброс больше - 25-50°. В боковых (особенно тупиковых) ходах наклон обычно меньше и в среднем близок к 20°.
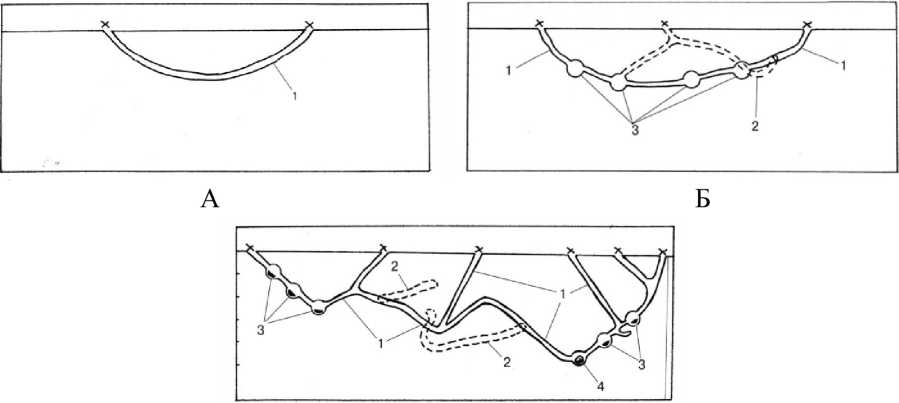
По строению можно выделить три типа нор: 1) бескамерные, 2) норы с камерами для запасов без гнезда и 3) норы с камерами для запасов и гнезда (рис. 1).
Бескамерная нора представляет собой сравнительно простую систему подземных ходов без расширений с одним-двумя входными отверстиями. Часто - это просто тоннель под землей на глубине 25-30 см, длиной от 0,3 до 2 м, с двумя отверстиями на поверхности. Диаметр хода варьирует от 5 до 10-12 см, в зависимости от характера грунта и максимальных размеров зверьков, живущих в норе. Норы этого типа чаще встречаются на окраинах колоний, в местах кормежек; вне колоний это практически единственный тип нор.
В норах с камерами в ходах имеются расширения, или камеры (до 7 на нору), нередко вблизи входных отверстий. В камерах мы находили пучки свежей растительности, в некоторых - подстилку из сухой травы. Глубина ходов варьирует в зависимости от экспозиции и характера грунта: на склонах и при значительном участии каменистого материала не превышает 40 см, в понижениях - до 60 см. Диаметр - 15- привходового отверстия находится в диапазоне от 5 до 20 см, у более глубоких ходов он обычно составляет 10 см, но могут встречаться и внекамерные расширения до 15 см. Диаметр камер доходит до 20-25 см. Протяженность ходов суммарно варьирует от 3 до 5 м, из которых на боковые ходы приходится до 1,5 м, редко - до 2 м. Норы с камерами встречаются на всей территории колонии.
В
Рис. 1 . А. Бескамерная нора даурской пищухи.
Б. Нора с камерами. В. Гнездовая нора: х — отверстия норы, 1 — магистральный ход, 2 — боковой ход, 3 — камеры с запасами, 4 — гнездовая камера
Гнездовые норы с камерами для запасов имеют до 10 входных отверстий самого разного диаметра (от 7 до 20 и более сантиметров), гнездовую камеру и до 5 камер с запасами корма. Выстилка гнезда состоит из нескольких слоев, содержащих преимущественно траву и опушенные соцветия степных растений, пух птиц, иногда клочки шерсти овец. Параметры нор практически те же, что и у предыдущего типа, а глубина может превышать 1 м (Борисова и др., 2001). Протяженность больше, чем у других типов нор, - до 9,2 м, на долю магистральных ходов приходится 5,8 м, а боковых - 3,4 м.
Температура в норах варьирует в зависимости от сезона, глубины и расстояния от привходового отверстия (табл. 1): в июне на расстоянии 35 см от привходового отверстия, на глубине 20-25 см, она составила +19°С при температуре +35°С на высоте 7-10 см от поверхности. В августе температура в том же участке норы почти та же самая, что и в июне, хотя температура воздуха на поверхности в это время уже значительно ниже, а температура грунта выше. В конце сентября температура снижается.
В 2014 г. при измерениях температуры в жилых, нежилых норах и в грунте вне нор были получены следующие данные (табл. 2): в жилых норах в августе и сентябре температуры более стабильны по сравнению с грунтом и нежилыми норами, при этом в жилых норах снижение дневной температуры к концу сентября менее резкое, чем в нежилых, а температуры в жилых норах и грунте, между которыми в начале августа существовали заметные различия (до 7°С), к концу сентября различаются незначительно (2°С).
Температурный режим нор даурских пищух
Таблица 1
|
^""^Температурный ^"\пежим Период |
t°C на поверхности |
t°C в 30-35 см от входа |
t°C в глубине |
|
1-я пол. июня 11ч утра |
35 |
19 |
- |
|
34 |
19 |
- |
|
|
30 |
17 |
- |
|
|
28 |
16 |
- |
|
|
25 |
15 |
- |
|
|
27 |
17 |
- |
|
|
2-я пол. августа 11ч утра |
27 |
21 |
20 |
|
25 |
18 |
17 |
|
|
24 |
16 |
17 |
|
|
2-я пол. августа 11ч утра |
25 |
18 |
16 |
|
20 |
16 |
15 |
|
|
22 |
17 |
17 |
|
|
27 |
19 |
18 |
|
|
21 |
18 |
15 |
|
|
22 |
18 |
19 |
|
|
21 |
18 |
11 |
|
|
2-я пол. сентября 11ч утра |
13 |
10 |
8 |
|
12 |
7 |
6 |
|
|
12 |
8 |
7,5 |
|
|
10 |
8,5 |
6,9 |
|
|
5 |
1 |
3 |
|
|
5 |
2,5 |
3 |
|
|
15 |
12 |
5 |
|
|
8 |
5 |
6,5 |
|
|
9 |
7 |
7 |
|
|
15 |
10,5 |
8 |
|
|
11 |
8 |
7 |
|
|
13 |
7 |
7 |
|
|
12 |
9 |
3 |
|
|
10 |
7 |
8 |
|
|
14 |
9 |
6 |
|
|
10 |
7 |
7 |
Почвы в 2014 г. характеризовались высоким уровнем влажности: на склоновых каштановых почвах она редко опускалась ниже отметки 85% (в ночное время), а днем доходила до 93%. В первой половине сентября дожди стали реже, влажность почв начала снижаться; в нежилых норах и грунте она оставалась максимальной лишь в дневные и вечерние часы, а ночью и утром падала до 90%. В жилых норах влажность все это время постоянно держалась на уровне 83-85%.
Таблица 2
Средняя температура в жилых, нежилых норах и в грунте вне нор по сезонам 2014 г. и в разное время суток, °C
|
Сезон |
Заселенность нор |
||||||||
|
Жилые |
Нежилые |
Грунт без нор |
|||||||
|
t суток |
8:00 |
16:00 |
24:00 |
8:00 |
16:00 |
24:00 |
8:00 |
16:00 |
24:00 |
|
1-я пол. августа |
15,6 |
18,5 |
17,2 |
17,4 |
22,2 |
19,8 |
18,7 |
25,5 |
22,1 |
|
2-я пол. августа |
И,7 |
21,5 |
15,7 |
16,7 |
24,3 |
19,3 |
19,5 |
26,4 |
22,8 |
|
1-я пол. сентября |
10,4 |
15,9 |
14,3 |
10,2 |
16,5 |
13,2 |
9,8 |
16,1 |
13,3 |
|
2-я пол. сентября |
9,8 |
16,4 |
12,6 |
8,1 |
12,5 |
10,6 |
9,5 |
18,6 |
12,5 |
С середины сентября влажность стала ниже - 75-80%; влажность грунта изменилась менее значительно (до 85-90%). В жилых норах влажность в ночное время опускалась до 85%, а в дневные доходила до 96-98%; в нежилых норах она ночью не была ниже 90%, а в дневные составляла 100%, мало отличаясь от показателей в грунте (93% ночью и 100% днем).
Суточная активность.
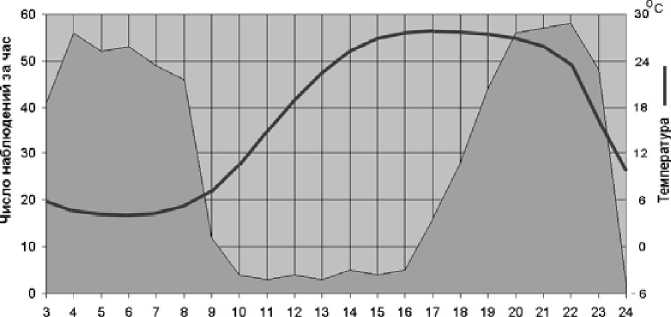
В летний период дневные температуры в Юго-Западном Забайкалье высоки; активность пищух в это время на поверхности носила ярко выраженный бимодальный характер: с пиками активности в часы после восхода и перед закатом солнца (рис. 2).
Час суток
Рис. 2. Суточная активность пищух в конце мая — начале июня 1995 г. и среднесуточный ход температур воздуха
К концу лета, по мере понижения дневных температур, активность принимала все более диффузный характер без ярко выраженных пиков. В сумеречные нежаркие дни летом активность становилась также более диффузной (рис. 3).
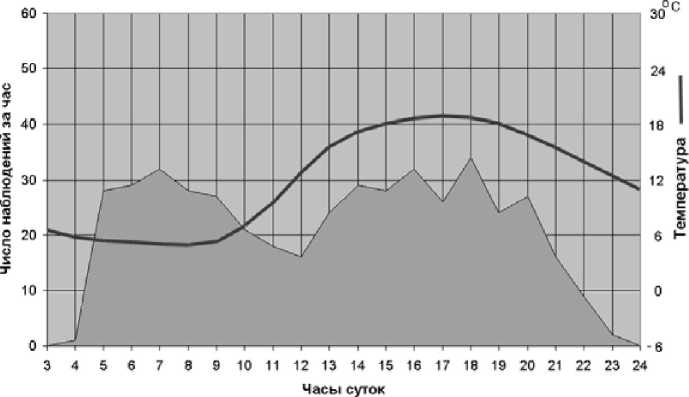
Рис. 3 . Суточная активность пищух в августе 1995 г. и среднесуточный ход температур воздуха, нежаркий день
С наступлением осени период активности сместился на все более узкий временной интервал; утренние и вечерние пики почти исчезли (рис. 4).
°с
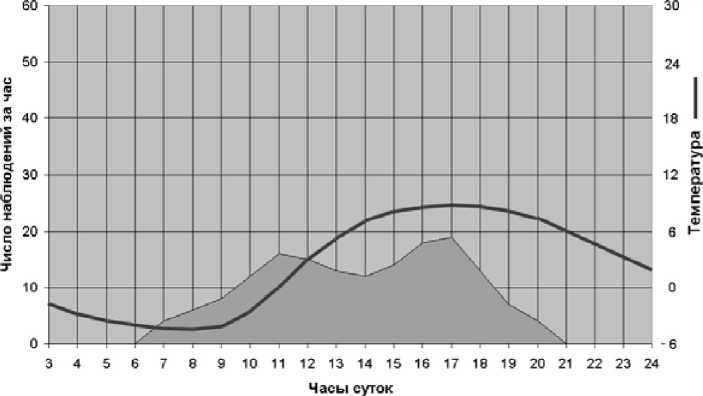
Рис. 4. Суточная активность пищух в начале октября 1997 г. и среднесуточный ход температур воздуха
Обсуждение. Несмотря на то, что строение нор даурской пищухи достаточно хорошо изучено во многих частях ареала вида (Фетисов, 1942; Некипелов, 1954; Швецов, Московский, 1961, Хабаева, 1975; Матурова, Доржиев, Иванова, 1977 и др.), описания, сделанные ранее, преимущественно качественные, поэтому количественное сравнение с нашими данными невозможно. Что касается климатических условий в норах даурской пищухи, то ранее они не оценивались, за исключением влажности в норах даурских пищух в Убсу-Нурской котловине (Ондар, 2001). Там установлено, что во влажные периоды она поддерживается на уровне от 10% в верхних горизонтах нор до 15% на глубине 70 см и от б до 18% - в засушливые, что выше, чем в почвах вне колоний.
В жилых норах другой обитательницы степей, черногубой пищухи, О. curzoniae, на глубине 10 см влажность была выше по сравнению с нежилыми: 44,5-54% против 39,5-41%. На глубине от 31 до 50 см влажность в жилых и нежилых норах выравнивалась и составляла 30% (Li, Zhang, 2006).
Количественных данных по суточной наземной активности даурских пищух до сих пор опубликовано не было. Нами установлено, что даурская пищуха активна преимущественно в светлое время суток. Основные пики активности приходятся на время восхода и заката и окружающие 1-2 часа. В пасмурные дни изменения активности были менее выраженными.
У черногубой пищухи, населяющей степные биотопы Тибета, также обнаружено два пика активности (Zong, Xia, 1987). Было установлено, что чем выше освещенность, тем чаще пищухи активны на поверхности, но более короткое время. Соотношение продолжительности периодов активности и покоя по часам в январе составляло 11/13 ч, в апреле оно было обратным, а летом - 15/9. По-
Вестник ТвГУ. Серия "Биология и экология". 2014. № 4 видимому, имеют значение и определенные температуры: судя по лабораторным данным, при температуре ЗО°С и 25°С в течение 24,16 ч и 24,45 ч, соответственно, пищухи проявляли активность 5761 и 3116 раз, тогда как, например, при температуре 15°С за 23,33 ч - всего 962 раза. Хоккайдский подвид северной пищухи, О. hyperborea yesoensis, имеет сходный, с двумя пиками, циркадианный тип активности (Kawamichi, 1969). Период как утренней, так и вечерней активности по отдельности составляет около 2,5 ч. Этот ритм практически не меняется в течение всего бесснежного периода, однако часы активности могут варьировать по месяцам вслед за временем захода и восхода солнца, причем наблюдается отрицательная корреляция между инсоляцией и поверхностной активностью, в пасмурную погоду период активности удлиняется на 1-2 часа. У видов, обитающих в Гималаях, наблюдаются различия в суточной активности. Если у пищухи Ройли, О. roylei, двухфазный тип активности, то у большеухой, О. macrotis, он однофазный, с 9 до 17 ч (Kawamichi, 1971). Автор объясняет это различиями в местообитаниях двух видов: большеухая пищуха живет в нагромождениях каменных глыб и валунов в высокогорье, где всегда царит полумрак; солнечные лучи проникают туда, и то не всегда, в основном в полуденные часы, при высоком положении солнца. В местах обитания пищухи Ройли (горные хвойные, смешанные и тропические дождевые леса с выходами скал) сложность рельефа гораздо меньше, и пищухи утром начинают деятельность в 5 ч утра, в темноте, с утренним пиком в 7 ч, а на 17 ч приходится вечерний пик активности.
У американской (Smith, 1974), рыжеватой (Yang, 1990), черногубой (Wang, Wang, 1989) установлена физиологическая непереносимость высоких температур.
В сравнении с более крупными зайцеобразными пищухи имеют более высокий уровень обмена веществ, что обеспечивает большую выносливость их к холоду, чем к летней жаре. У черногубых пищух термогенез, осуществляемый в бурой жировой ткани, происходит в течение всего года и варьирует по сезонам. По сравнению с кроликами у черногубых пищух плохо развита теплоотдача: скорость дыхания у них не повышается даже при 35°С, не регулируется эндогенно повышение ректальной температуры, незначительна иррадиация тепла от наружного уха за счет меньшего отношения поверхности уха к поверхности тела. Кроме того, у пищух почти не вырабатываются специфические белки теплового шока (Yang, 1990).
Разные виды пищух имеют сходные физиологические особенности. Было отмечено, что у черногубых пищух средняя ректальная температура составляет 39,8±1,2°С (п=17), а у рыжеватых (О. rufescens) - 39,6°С (Kosaka et ah, 1985), средняя скорость дыхания у этих двух видов была так же сходна, в среднем составляя 110,5±14,1 в минуту, что говорит о том, что общие особенности циркуляции крови и " " -21 - '
терморегуляции являются адаптацией к обитанию в условиях высокогорий с их низкими температурами.
Мы полагаем, что физиологическая неприспособленность к высоким температурам присутствует и у даурских пищух и определяет в значительной степени режим их наземной активности летом. Из отрицательной корреляции с температурой в жаркие сезоны года следует, что именно температура, а не фотопериод определяет характер временного распределения суточной наземной активности даурской пищухи в это время года. Примечательно, что у близкого вида, черногубой пищухи, обнаружена (в отличие от другого незимоспящего мелкого млекопитающего Тибетского плато - полевки-экономки (Microtus oeconomus), подготовка к зиме. Заключающаяся в изменении скорости метаболизма, появлении термогенезиса без дрожи, увеличении содержания протеина и оксидазной активности цитохрома С в митохондриях бурого жира, подготовка запускается низкими температурами, а не изменениями фотопериода (Wang et al., 1999).
Авторы выражают глубокую благодарность сотруднику Музея природы И.В. Шимову и сотрудникам ИОЭБ СО РАН А.В. Сотниковой - за помощь в проведении работ и к.б.н. А.Ц. Мангатаеву - за участие в обработке материалов.
Список литературы Экология даурской пищухи на северной периферии ареала
- Борисова Н.Г., Старков А.И., Сазонов Г.И., Руднева Л.В. 2001. К экологии даурской пищухи в Юго-Западном Забайкалье//Тр. ЗИН РАН. Т. 288. С. 163-179.
- Матурова Р.Т., Доржиев Ц.З., Иванова Г.И. Грызуны и зайцеобразные западной части Заганского хребта и Тугнуйской котловины//Насекомые и позвоночные Забайкалья. Улан-Удэ. С. 83-102.
- Некипелов Н.В. 1954. Изменение численности даурской пищухи в юго-западном Забайкалье//Изв. Иркут. н.-и. противочум. ин-та. Т. 12. С. 171-180.
- Ондар С.О. 2001. Механизмы функционирования ультраконтинентальных степей: устойчивость и динамические процессы: автореф. дисс.. д-ра биол. наук. М. Изд-во МГУ. 46 с.
- Фетисов А.С. 1942. Зимние кормовые объекты куницеобразных Забайкалья//Изв. Биол.-геогр. НИИ при Иркут. ун-те. Т. 9. Вып. 3-4. С. 145-151.
- Хабаева Г.М. 1975. О распространении и экологии грызунов северной части юго-западного Забайкалья//Зоологические исследования в Забайкалье. Улан-Удэ: Бурят. кн. изд-во. С. 238-241.
- Швецов Ю.Г., Московский А.А. 1961. Распространение и численность основных видов грызунов в пограничной полосе Юго-Западного Забайкалья//Биологический сборник. Иркутск. С. 96-106.
- Kawamichi T. 1969. Behavior and daily activities of the Japanese Pika, Ochotona hyperborea yesoensis//J. Fac. Sci. Hokkaido Univ., Japan, Ser. VI. Zool. № 17. P. 127-151.
- Kawamichi T. 1971. Daily activities and social pattern of two Himalayan pikas, Ochotona macrotis and O. roylei, observed at Mt. Everest//J. Fac. Sci. Hokkaido Univ., Japan, Ser. VI. Zool. V. 17. № 4. P. 587-609.
- Kosaka M., Ohwatari N., Iwamoto J., Tsuchiya K., Fujiwara M., Fan Yu-J., Matsuo T., Moriuchi T., Matsuzaki T. 1985. Studies of temperature regulation on pika (Ochotona rufescens rufescens); an old-fashioned rabbit//Trop. Med. V. 27. № 4. P. 289-294.
- Li W.J., Zhang Y.M. 2006. Impacts of plateau pikas on soil organic matter and moisture content in alpine meadow//Acta Theriol. Sinica. V. 26. № 4. P. 331-337.
- Smith A.T. 1974. The distribution and dispersal of pikas: influences of behavior and climate//Ecology. V. 55. P. 1368-1376.
- Wang D., Wang Z. 1989. Strategies for survival of small mammals in a cold alpine environment I. Seasonal variations in the weight and structure of brown adipose tissue in Ochotona curzoniae and Microtus oeconomus (in Chinese with English summary)//Acta Theriol. Sinica. V. 9. P. 176-185.
- Wang D.H., Sun R.Y., Wang Z.W., Liu J.S. 1999. Effects of temperature and photoperiod on thermogenesis in plateau pikas (Ochotona curzoniae) and root voles (Microtus oeconomus)//J. Comp. Physiol. B. V. 169. No. 1. P. 77-83.
- Yang G.-J. 1999. Physiological characteristics of pika (Ochotona rufescens rufescens) as a weak heat tolerant animal//Trop. Med. V. 32. № 4. P. 129-140.
- Zong H., Xia W. 1987. Circadian activity rhythms of plateau pikas, Ochotona curzoniae//Acta Theriol. Sinica. V. 7. № 3. P. 211-223.
