Экология питания белолобого гуся Anser albifrons на острове Колгуев в предгнездовой период

Автор: Зайнагутдинова Э.М.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 530 т.18, 2009 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140151567
IDR: 140151567
Feeding ecology of the greater white-fronted goose Anser albifrons on Kolguev island during prenesting period
Текст статьи Экология питания белолобого гуся Anser albifrons на острове Колгуев в предгнездовой период
Самым распространенным видом гусей в мире, в том числе самым многочисленным видом гусей в Европе и в России, является белолобый гусь Anser albifrons , имеющий практически циркумполярное распространение (Ely, Dzubun 1994; Mooij et al . 1999). Белолобый гусь – один из основных охотничьих видов (Mooij et al . 1999), а также важная составляющая агроценозов (Fox et al . 2005; Зимин и др. 2007) и арктических экосистем (Van der Wal et al . 2001; Mooij et al . 2005; Alisauskas et al . 2006). Для предсказания динамики численности белолобого гуся и регулирования интенсивности нагрузки на его популяции необходимо определение механизмов, влияющих на изменение численности, в первую очередь на репродуктивный успех в ключевых районах размножения.
Белолобые гуси проводят предгнездовой период в репродуктивном районе (Ely, Raveling 1984; Fox 2003). Самки за это время должны запасти значительное количество резервных веществ в теле для продукции яиц и инкубации, поэтому кормовые условия предгнездового периода оказывают существенное влияние на успех размножения птиц (Drent, Daan 1980).
Одним из наиболее важных районов размножения белолобого гуся является остров Колгуев, расположенный на юго-востоке Баренцева моря, где плотность гнездования этих гусей в десятки раз превышает плотности гнездования в других районах размножения вида (Зайнагутдинова, Кондратьев 2008). Благодаря морскому климату и близости Северо-Aтлантического тёплого течения весна на острове обычно ранняя, в результате в предгнездовой период гуси имеют возможность кормиться на обширных территориях. Уникальность Колгуева состоит в том, что это достаточно крупный арктический остров, на котором полностью отсутствуют грызуны (Морозов, Сыроечковский–мл. 2004). В связи с этим активность хищников не испытывает значительных межгодовых колебаний (Bêty et al. 2003), что благоприятно сказывается на успешности размножения гусей (Summers 1986). Однако высокие концентрации гусей на территории могут приводить к росту внутривидовой конкуренции за кормовые ресурсы и снижению успеха размножения гусей, поэтому задачей исследования было определение благоприятных и неблагоприятных факторов, воздействующих на белолобых гусей на Колгуеве в предгнездовой период.
Материалы и методы исследования
Исследования проводились на острове Колгуев в период с 29 мая по 20 июня 2006-2008 годов. Для определения распределения гусей по острову в предгнездовой период пешеходные маршрутные учёты птиц на основе учётных трансект фиксированной ширины в 600 м проводились с конца мая (времени массового прилёта белолобых гусей) до середины июня (времени массового гнездования и отлёта части размножающихся птиц с острова). При учёте птиц фиксировали их координаты, расстояние до наблюдателя, биотоп и рельеф. Упитанность птиц определяли визуально по семибалльной шкале на основе формы живота (Moriguchi 2006), пол гусей определяли, основываясь на поведенческих различиях (Black, Owen 1988). Всего в предгнездовой период было проведено 275 км маршрутных учётов и учтено более 7000 особей.
Долю снежного покрова на исследованных территориях в середине июня определяли на основе космического снимка Landsat 7 от 17 июня 2002, который в ГИС программе MapInfo Professional 9.0 был разбит на квадраты UTM сетки со стороной 1 км. Затем путём градационного анализа изображения в программе Adobe PhotoShop для каждого квадрата была определена доля озёр и доля снежного покрова на территории. Для определения суточного хода температур использованы данные с метеостанции в посёлке Бугрино.
Интенсивность питания и перемещения гусей во время питания определяли в ходе фокальных наблюдений (Madsen 1985; Martin, Bateson 1996). При помощи секундомера определяли время, затрачиваемое на 30 клевков, одновременно механическим счётчиком фиксировали число шагов, на которые переместилась птица. Для одной наблюдаемой особи такая процедура по возможности повторялась десять раз. Как только птица уходила из поля зрения, перемещалась в другой биотоп или прекращала кормиться, наблюдения прекращались. При этом фиксировали социальный ранг птицы, пол, упитанность, записывали растительную ассоциацию, рельеф, район наблюдения и расстояние до водоёма. Так как в небольших стаях между птицами на краю стаи и в её центре различий в интенсивности перемещения и времени, затраченном на настороженное поведение, выявлено не было (Carbone et al . 2003), то наблюдения проводились за любыми доступными птицами. Наблюдения велись только за кормящимися особями, если птицы сменяли активность, например, на отдых, беспокойство или целенаправленные перемещения, то наблюдения прекращались. Наблюдения за интенсивностью питания и перемещения были проведены над 92 птицами.
Статистическая обработка данных проведена в программе STATISTICA 8. Все выборки были проверены на нормальность распределения. Для выборок, распределение которых удовлетворительно аппроксимировалось нормальным, или больших выборок, содержащих более 100 переменных, применяли методы параметрической статистики. Для малых выборок, распределение которых не являлось нормальным, применяли методы непараметрической статистики. Для анализа данных использованы критерий Стьюдента, однофакторный дисперсионный анализ, и U-критерий Манна-Уитни (Dytham 2003; Халафян 2007).
Результаты
Среднесуточные температуры июня в 2007-2008 годах были значительно ниже, чем в 2006 ( Z = 3.7, P < 0.05). В 2008 году наблюдалось некоторое понижение температуры в мае ( Z = 0.1, P = 0.08) и значительное снижение температуры в апреле ( Z = 2.9, P < 0.05) по сравнению с 2007 годом, что вызвало наиболее поздние сроки схода снежного покрова.
На остров Колгуев гуси начинали прилетать в начале мая, интенсивный прилёт наблюдался в конце мая – начале июня, а в 2008 – в год с поздней весной – миграция наблюдалась до 12 июня. Распределение гусей по территории острова в предгнездовой период было достаточно равномерным и слабо зависело от рельефа [ F (2,82) = 0.67, P = 0.51] и времени схода снежного покрова в разных районах острова [ F (2,120) = 2.59, P = 0.11].
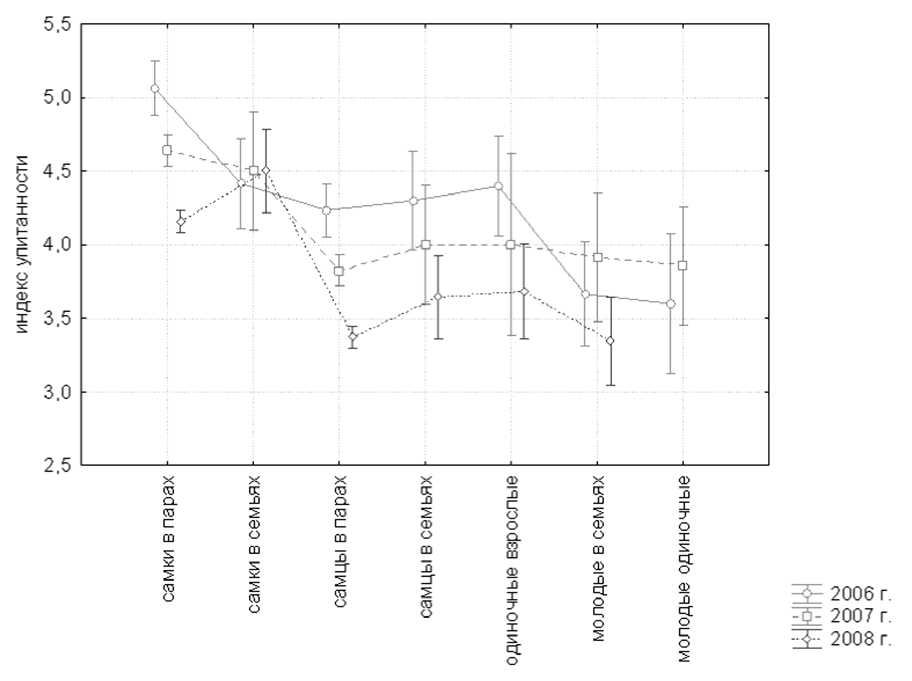
Рис. 1. Индекс упитанности белолобых гусей разного социального статуса в предгнездовой период 2006-2008 годов.
В зависимости от социального статуса птицы имели разную упитанность (рис. 1). Самки отличались наибольшей упитанностью в пред-гнездовой период, индекс их упитанности составлял от 4.2±0.04 до 5.1±0.1. Средний индекс упитанности самцов был значимо ниже, чем самок, и составлял 3.4±0.03 – 4.2±0.1 (F = 238, P < 0.05). Упитанность молодых птиц (3.8±0.03 — 4.6±0.07) была значимо ниже, чем упитанность взрослых (в 2006 г. t = 3.57, P < 0.05, n 1 = 89, n2 = 14; в 2007 г. t = 1.84, P < 0.05, n 1 = 218, n2 = 13; в 2008 г. t = 3.24, P < 0.05, n 1 = 453, n 2 = 13).
Погодные условия предгнездового периода значимо влияли на упитанность птиц. В сезон с наиболее высокими температурами июня наблюдалась наибольшая упитанность самцов (4.2 ± 0.1) и самок (5.1 ± 0.1), в сезон с самыми низкими температурами июня отмечена наименьшая упитанность самцов (3.4 ± 0.1) и самок (4.2 ± 0.1) [для самцов F (2,349) = 60.80, P < 0.05; для самок F (2,341) = 51.97, P < 0.05]. Упитанность молодых птиц также значимо снижалась в год с наиболее холодным июнем ( t = 2.41, P < 0.05, n 1 = 13, n 2 = 13).
Так как активный рост зелёных частей растений в предгнездовой период ещё не начался, гуси питались основаниями побегов осоки водной Carex aquatilis и пушицы влагалищной Eriophorum vaginatum , выкапывая и выдёргивая их из субстрата в осоково-моховых и пушицево-моховых растительных ассоциациях.
Нами была определена интенсивность питания гусей в наиболее широко распространённых биотопах острова. Интенсивность питания в 2007 году была наибольшей в бугристых редко-ивняково-моховых тундрах на склонах долины реки (70 ± 4 клевков/мин), средней на осоково-моховом болоте в долине реки (43 ± 1) и наименьшей в кустарничково-лишайниково-моховых тундрах на возвышенностях (32 ± 1) ( F = 54.52, P < 0.05). В наиболее холодном 2008 году интенсивность питания была одинаково низкой во всех биотопах: F (2,56) = 1.44, P = 0.25).
Погодные условия влияли на интенсивность питания птиц. В 2008 году, когда наблюдалось наиболее позднее таяние снега, отмечена наименьшая интенсивность питания (32 ± 1 клевков/мин). В 2007 году, когда снег сошёл в более ранние сроки, интенсивность питания была выше и составляла 44 ± 1 клевков/мин ( t = 6.8, P < 0.05, n 1 = 185, n 2 = 173). В 2008 году с более поздней весной самцы и самки питались с разной интенсивностью. Интенсивность питания самцов снижалась до 24 ± 2, а интенсивность питания самок составляла 36 ± 1 клевков/мин ( t = 5.61, P < 0.05, n 1 = 64, n 2 = 121). В 2007 году, характеризовавшимся более ранним сходом снега, самцы и самки питались с одинаковой интенсивностью ( t = 0.53, P = 0.30, n 1 = 64, n 2 = 109).
Из-за высокой численности белолобых гусей на Колгуеве, ведущей к росту внутривидовой конкуренции, интенсивность питания птиц более низкого социального ранга с низкой упитанностью была ниже, чем у птиц более высокого социального ранга, имеющих большую упитанность (рис. 2). В 2007 году на осоково-моховом болоте в долине реки интенсивность питания самцов снижалась с 52 ± 5 до 37 ± 2 клевков в 1 мин в зависимости от их индекса упитанности [ F (1, 28) = 9.05,
P = 0.01] и с 47 ± 4 до 36 ± 2 клевков/мин в зависимости от индекса упитанности партнёра по паре [ F (1,27) = 6.33, P = 0.02]. Интенсивность питания самок снижалась с 42 ± 2 до 39 ± 1 клевков/мин в зависимости от их индекса упитанности [ F (2,67) = 3.37, P = 0.04] и с 44 ± 2 до 38 ± 1 клевков/мин в зависимости от индекса упитанности партнёра по паре [ F (1, 63) = 4.50, P = 0.04]. В 2008 году интенсивность питания самцов с индексом упитанности 3 (21 ± 2 клевков/мин) была ниже, чем у самцов с индексом упитанности 4 (32 ± 3) [ F (1,62) = 5.67, P = 0.02] и не зависела от индекса упитанности самки [ F (1,61) = 0.61, P = 0.44]. Аналогично интенсивность питания самок с индексом упитанности 4 (34 ± 1 клевков в 1 мин) была ниже, чем у более упитанных самок с индексом 5 (41 ± 4) [ F (1,125) = 5.51, P = 0.02] и не зависела от индекса упитанности самца [ F (1,129) = 0.01, P = 0.92].
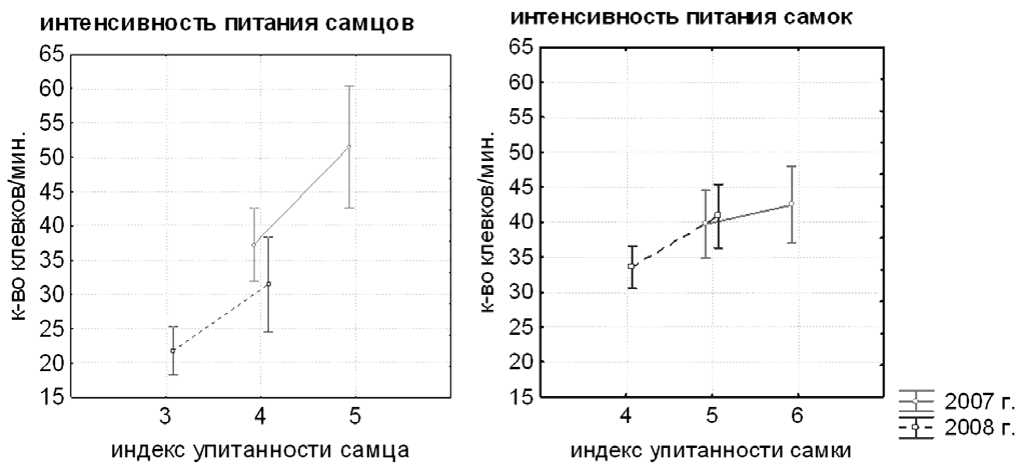
Рис. 2. Изменение интенсивности питания белолобых гусей (количество клевков в 1 мин) в зависимости от их индекса упитанности в 2007-2008 годах.
Нами также была определена интенсивность перемещения гусей во время питания (число шагов в 1 мин), которая, как и интенсивность питания, характеризует доступность кормов. Так как проективное покрытие кормовых растений не зависит от сроков схода снега, интенсивность перемещения птиц во время питания не различалась значимо в сезон с менее холодным июнем 2007 г. и в сезон с наиболее холодным июнем 2008 г. ( F = 2.65, P = 0.10).
Поскольку во время кормления самец и самка всегда держатся вместе и перемещаются одновременно, то интенсивность перемещений самцов и самок также не различалась значимо (в 2007 г. F = 2.65, P = 0.10; в 2008 г. F = 0.26, P = 0.61).
В разных биотопах в связи с различной встречаемостью осок и пушиц интенсивность перемещения белолобых гусей существенно различалась. Для того, чтобы найти наиболее благоприятные для кормления биотопы, интенсивность перемещения гусей была определена в наиболее распространённых биотопах острова. В 2007 году интенсивность перемещений птиц на кустарничково-лишайниково-моховой тундре на возвышенности (22 ± 2 шага/мин) значимо превышала этот показатель на осоково-моховом болоте в долине реки (17 ± 1, t = 3.06, t кр = 1.68, P < 0.05, n 1 = 30, n 2 = 116) и на бугристой редко-ивняково-моховой тундре на склонах (14 ± 1, t = 3.06, t кр = 3.16, P < 0.05, n 1 = 30, n 2 = 21). В 2008 году различия также были значимыми ( F = 11.43, P < 0.05).
Изменение интенсивности перемещений в течение июня может характеризовать доступность кормов в предгнездовой период, поэтому данный показатель был определён для птиц, кормящихся на осоковомоховом болоте в долине реки – биотопе, где наблюдения проводились за наибольшим количеством птиц. В 2008 год с наиболее поздней весной в связи с истощением кормовых ресурсов интенсивность перемещения значительно возрастала во вторую декаду июня по сравнению с первой декадой: от 5 ± 1 до 12 ± 2 шага в 1 мин ( t = 3.84, t кр = 1.68, P < 0.05, n 1 = 28, n 2 = 29). В 2007 год с более ранним сходом снега интенсивность перемещений снижалась до 8 ± 1 шагов/мин во вторую декаду июня в сравнении с первой декадой, когда она составляла 17 ± 1, однако эти различия не были значимыми ( t = 0.77, t кр = 1.69, P = 0.22, n 1 = 100, n 2 = 16).
Внутривидовая конкуренция может вести также к различиям в интенсивности перемещения особей разного социального ранга, поэтому интенсивность перемещений пар разного социального ранга была измерена в 2007 году для птиц, кормящихся на осоково-моховом болоте в долине реки (в этом биотопе наблюдения велись за наибольшим числом особей). Было выявлено незначительное увеличение интенсивности перемещения во время кормления у менее упитанных птиц: с 11 ± 2 до 18 ± 1 шагов/мин в зависимости от индекса упитанности самца ( F = 2.43, P = 0.12) и с 13 ± 2 до 19 ± 2 шагов/мин в зависимости от индекса упитанности самки ( F = 1.18, P = 0.31).
Обсуждение результатов
На остров Колгуев гуси начинают прилетать в начале мая, интенсивный прилёт наблюдается в конце мая – начале июня, а в год с поздней весной миграция продолжалась до середины июня. Схожие сроки прилёта белолобых гусей отмечены на Колгуеве в 1990-х годах В.В.Морозовым и Е.Е.Сыроечковским-младшим (2004), а также в других частях ареала (Кречмар 1986; Mooij et al . 1999; Рябицев 2002).
Предгнездовой период белолобых гусей на Колгуеве длится несколько недель, что позволяет предположить, что рост фолликулов начинается в районе размножения, и что здесь эти гуси в значительной мере «полагаются» на набор массы на местах гнездовий, поэтому ситуация, которая складывается на острове в предгнездовой период, может влиять на успех размножения гусей. В других районах белолобые гуси также проводят около 2 недель до начала гнездования в районе размножения (Ely, Raveling 1984; Fox 2003). Сроки весенней миграции белолобых гусей на Колгуеве варьируют в зависимости от погодных условий сезона, что было показано и для подвида A. a. frontalis (Ely, Raveling 1984; Кречмар 1986).
Во многих районах размножения белолобые гуси сразу после прилёта агрегируются на ограниченных участках, освободившихся от снега, и лишь после схода снегового покрова их распределение по территории становится более равномерным (Минеев 1995; Рябицев 2002; Кречмар, Кондратьев 2006). На Аляске и в Канаде белолобые гуси в предгнездовой период кормятся на приморских маршах (Fox, Madsen 1981; Fox, Ridgill 1985; Carriere et al . 1999; Ely et al . 2006). На Колгуеве приморские марши заняты колониями белощёких казарок Branta leucopsis (Anisimov 2007), поэтому белолобые гуси выбирают другие биотопы. Так как уже в мае значительная часть тундры свободна от снега и доступна для кормления, гуси равномерно распределяются по острову. Поэтому важным преимуществом острова Колгуев является широкая доступность кормовых биотопов в предгнездовой период по всему острову и возможность кормиться в районе гнездования. Для белолобых гусей характерна высокая пластичность в выборе кормовых местообитаний (Розенфельд 2005), однако важно, чтобы грунт не промерзал глубоко, так как основу питания в этот период составляют подземные части растений (Розенфельд 2005).
Ранний сход снега на равнинах и плато Колгуева позволяет гусям кормиться на обширных территориях, однако несколько чаще гуси на острове встречаются в травяно-моховой тундре на склонах холмов и долин, так как почва на них успевает оттаять раньше, а проективное покрытие осок выше, чем на возвышенностях, поэтому и интенсивность кормёжки гусей в этих биотопах выше, чем на плато. В других регионах гуси также предпочитают кормиться в сходных осоково-пушицево-моховых и кустарниково-пушицево-моховых сообществах (Жаркова, Боржонов 1972; Минеев 1995). С увеличением доступности кормов в весенний период интенсивность питания гусей возрастает (Prop, De Vries 1993), поэтому на Колгуеве в сезон со средними сроками схода снега интенсивность питания в течение июня возрастала, а интенсивность перемещений в течение июня снижалась. В годы с неблагоприятными погодными условиями предгнездового периода кормовые ус- ловия существенно ухудшаются (Fox et al. 2006), поэтому в сезон с наиболее поздней весной на Колгуеве интенсивность питания гусей в предгнездовой период не возрастала, а гнездование задерживалось. В сезон со средними сроками схода снежного покрова интенсивность питания белолобых гусей значительно варьировала в различных растительных ассоциациях, а в сезон с наиболее поздней весной интенсивность питания во всех растительных ассоциациях была низкой, так как почва не протаивала в значительной мере. Менее благоприятные кормовые биотопы играют буферную роль и используются, когда условия в предпочитаемых биотопах не оптимальны, в том числе, когда трава медленно возобновляется из-за плохой погоды (Black et al. 2007). Так как погодные условия предгнездового периода значимо влияют на упитанность гусей (Bauer et al. 2006), в годы с наиболее поздней весной упитанность белолобых гусей на Колгуеве была наименьшей. Таким образом, на острове наблюдаются достаточно благоприятные кормовые условия в предгнездовой период, которые, впрочем, могут существенно ухудшаться в годы с поздней весной.
Средний индекс упитанности самок на Колгуеве в предгнездовой период был достаточно высок, что характерно для гусей, так как самки должны накопить резервы тела для продуцирования яиц и периода инкубации (Raveling 1979; Boyd et al. 1998; Hübner 2006). Упитанность самцов белолобых гусей на Колгуеве в предгнездовой период была ниже упитанности самок, что является обычной тенденцией для гусей (Raveling 1979; Boyd, Fox 1995; Boyd 2000; Hübner 2006) и объясняется тем, что самец должен значительную часть времени тратить на охрану самки (Budeau et al. 1991; Gauthier, Tardif 1991; Johnson, Sibly 1993; LeSchack et al. 1998). Упитанность самцов белолобых гусей на Колгуеве весной в семьях с молодыми птицами прошлого года рождения была несколько выше, чем упитанность самцов в парах без сопровождения молодых особей. На миграционных остановках белолобых гусей в Исландии также отмечено, что упитанность самок и самцов в семьях с молодыми особями или со взрослыми потомками предыдущих лет также выше, чем упитанность птиц в парах (Boyd et al. 1998), что объясняется высоким социальным рангом семей и меньшими затратами на настороженное поведение (Ely 1993; Fox et al. 1995; Black et al. 2007). Упитанность одиночных белолобых гусей на Колгуеве в целом была ниже, чем упитанность птиц в парах и семьях, так как одиночные птицы имеют более низкий социальный ранг, чем птицы в семьях и парах (Иваницкий 2002). Наименее упитанными были молодые белолобые гуси. Это связано с тем, что социальный ранг молодых птиц ниже, чем взрослых (Иваницкий 2002). Аналогичная зависимость показана также для белощёких казарок на Шпицбергене (Black et al. 2007). Таким образом, в связи с социальной иерархией и внутривидо- вой конкуренцией упитанность низкоранговых особей существенно ниже, чем высокоранговых.
С ростом численности популяции гусей возрастает внутривидовая конкуренция. В результате доступ к лучшим кормовым ресурсам получают самые высокоранговые гусиные семьи, и снижается доля птиц, способных успешно размножаться (Black et al . 2007). Поэтому различия в упитанности гусей и в доступе к кормовым ресурсам могут характеризовать интенсивность внутривидовой конкуренции. Самки белолобых гусей на Колгуеве в предгнездовой период кормятся больше, чем самцы, что характерно для гусей в целом (Sedinger, Raveling 1990; Ка-рагичева 2005; Black et al . 2007). Интенсивность перемещений во время кормёжки самцов и самок не различается, так как во время кормления самец и самка всегда держатся вместе и перемещаются одновременно. Так как агрессивные взаимодействия среди кормящихся белолобых гусей достаточно редки, мы попробовали использовать индекс упитанности в качестве показателя социального ранга пары. Наиболее упитанные белолобые гуси на острове Колгуев кормились с большей интенсивностью, чем менее упитанные птицы. Было выявлено также незначительное снижение интенсивности перемещения во время кормёжки у более упитанных птиц. Полученные результаты свидетельствуют о наличии внутривидовой конкуренции за пищевые ресурсы не только у колониальных видов или у птиц в условиях высокой скученности на зимовках (Teunissen et al . 1985; Sedinger et al . 1995; Klaassen et al . 2006), но и у неколониальных видов в условиях высокой численности в период размножения.
Заключение
Прилетев на остров Колгуев весной, белолобые гуси могут проводить здесь до нескольких недель до начала гнездования. Основной задачей птиц в этот период является восстановление резервов тела, необходимых для размножения. Климатические условия территории существенно влияют на эффективность накопления жировых резервов. Так, в сезон с более тёплой погодой на острове миграция проходила раньше, а упитанность птиц и интенсивность питания были выше, чем в сезоны с более холодной весной. А в сезон с поздним приходом весны интенсивность перемещения гусей во время кормёжки в течение июня существенно возрастала. Среди районов размножения белолобого гуся остров Колгуев, благодаря тёплому Северо-Атлантическому течению, является одной из тех территорий, где весна наступает наиболее рано. Так как климатические условия предгнездового периода существенно влияет на кондицию гусей, приступающих к размножению, более благоприятные климатические условия в сравнении с другими регионами являются одной из причин высокой численности белолобых гусей на Колгуеве. Однако из-за высокой численности здесь этих птиц среди них возрастает внутривидовая конкуренция и отмечается снижение интенсивности питания и возрастание интенсивности перемещения во время кормёжки у птиц низкого социального ранга, имеющих низкую упитанность, в сравнении с птицами более высокого социального ранга, упитанность которых выше.
