Экология питания прыткой ящерицы (Lacerta agilis L.) в популяциях Среднего Урала
 в популяциях Среднего Урала")
Автор: Галицын Дмитрий Игоревич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.16, 2014 года.
Бесплатный доступ
В данной статье рассматривается таксономическое разнообразие и биомасса кормовых объектов прыткой ящерицы на примере северных популяций. Отмечена определенная специфика и избирательность в рационе рептилий западного и восточного склона Уральского хребта. Проведен анализ перекрывания спектров питания в популяциях, а также отмечены особенности суточного рациона рептилий и его сезонной динамики.
Средний урал, трофическая ниша, спектр питания, кормовые объекты, избирательность, суточный рацион, сезонная динамика
Короткий адрес: https://sciup.org/148203347
IDR: 148203347 | УДК: 574.38:
Sand lizard (Lacerta agilis L.) nutrition ecology in the Middle Urals populations
On the northern populations example the sand lizard food items taxonomic diversity and biomass are considered in this paper. The certain nutrition features and selectivity of the reptiles in the western and eastern hillside of the Urals Mountains are noticed. The nutrition spectra overlap analysis is carried, and daily ration features of the reptiles and its season dynamics are noticed.
Текст научной статьи Экология питания прыткой ящерицы (Lacerta agilis L.) в популяциях Среднего Урала
П ищевые взаимоотношения в сообществе животных представляют первоочередной интерес с точки зрения роли определенного вида в круговороте веществ и потоке энергии. Сопряженный с этим термин «трофическая ниша» в большинстве случаев означает спектр питания вида и используется для описания его ключевых функций в биоценозах [6]. Кроме того, разнообразие пищевых объектов может определять степень экологической пластичности вида или популяции в определенных условиях, что особенно актуально для экологически разнородных внутривидовых групп [9].
Прыткая ящерица – один из таких примеров. Это широкоареальный лесостепной и степной вид, населяющий коренные и трансформированные территории с большим разнообразием местных условий. Существует достаточно много данных, описывающих трофическую нишу L. agilis , в особенности для западных частей ареала. В монографическом описании вида обобщены данные по набору кормов, сезонной и многолетней динамике и другим аспектам питания [11]. Отмечено большое разнообразие кормовых объектов, качественный и количественный состав которых определяется рядом факторов. В то же время, для популяций L. agilis , населяющих западный и восточный склоны Среднего Урала данных сведений очень немного – чаще всего они включены в общие фаунистические работы [1]. Подробных исследований, связанных с экологией питания L. agilis Среднего Урала ранее не проводилось.
В настоящей работе проведено обобщение полученных сведений о питании прыткой ящерицы Среднего Урала (на примере западного и восточного макросклонов Уральского хребта на территории Свердловской области) в дополнение к литературным данным для других частей ареала.
МАТЕРИАЛ И МЕТОДЫ
Изучение спектров питания, а именно таксономического разнообразия кормовых объектов L. agilis было основано на анализе содержимого желудочно-кишечного тракта. Выполнен анализ содержимого пищеварительных трактов 292 половозрелых особей прыткой ящерицы, в который включены сборы на протяжении трех сезонов 2011-2013 гг. Для организации маршрутных учетов [2] были выбраны локалитеты на западном и восточном склонах Уральского хребта с постоянным обитанием и достаточно большой численностью L. agilis :
-
1. Зона многоэтажной застройки в черте г. Каменска-Уральского площадью 8,96 га;
-
2. Окрестности железнодорожной станции Кунавино на территории Каменского района площадью 12,51 га;
-
3. Участок Красноуфимской лесостепи в окрестностях пос. Новый Бугалыш площадью 22,82 га.
Таксономическую принадлежность кормовых животных устанавливали при помощи определительных таблиц [5, 7, 10] до минимально возможной группы. Также проводили фаунистический учет [12] для оценки встречаемости кормовых объектов во внешней среде в районах кормодобывания. Сравнение спектров питания рептилий на западном и восточном склоне проводили по модифицированному индексу Мориситы [3, 8, 13]. Для оценки дистанцированности территориальных групп по спектрам питания был проведен кластерный анализ с учетом обобщенного Евклидова расстояния. Кроме того, учитывали среднюю биомассу кормовых объектов в пищеварительном тракте для количественной оценки потребляемого корма, суточного рациона животных и его возможных сезонных вариаций. Первичный анализ и статистическую обработку данных проводили по общепринятым методикам [4] при помощи программных пакетов MS Excel и Statistica 8.0.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В рационе прыткой ящерицы на западном и восточном склонах Среднего Урала были отмечены представители трех типов беспозвоночных: Arthropoda, Annelida, Mollusca. При этом основную роль в питании играли членистоногие. Среди них в пищеварительном тракте были найдены представители 27 семейств 9 отрядов насекомых, 1 отряда многоножек и 2 отрядов паукообразных. До видовой принадлежности было определено 118 кормовых животных. Таксономический состав (до ранга семейств), встречаемость и усредненная биомасса кормовых объектов представлены в табл. 1.
Таблица 1. Состав корма L. agilis на западном и восточном склоне Среднего Урала
|
Кормовые объекты |
Количество экземпляров |
Биомасса |
||||
|
Во внешней среде |
В пищеварительном тракте |
|||||
|
Абс. |
% |
Абс. |
% |
Абс., мг |
% |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Западный склон |
||||||
|
Arthropoda |
745 |
90,9 |
579 |
91,8 |
640,6 |
77,1 |
|
Arachnida |
28 |
3,4 |
45 |
7,1 |
84,4 |
10,2 |
|
Aranei |
17 |
2,1 |
41 |
6,5 |
76,5 |
9,2 |
|
Araneidae |
11 |
1,3 |
18 |
2,9 |
39,0 |
4,7 |
|
Thomisidae |
3 |
0,4 |
16 |
2,5 |
24,7 |
3,0 |
|
Lycosidae Acaromorpha |
3 |
0,4 |
7 |
1,1 |
12,8 |
1,5 |
|
Ixodidae Myriapoda |
11 |
1,3 |
4 |
0,6 |
7,9 |
1,0 |
|
Diplopoda |
1 |
0,1 |
3 |
0,5 |
5,1 |
0,6 |
|
Hexapoda |
716 |
87,4 |
531 |
84,2 |
551,1 |
66,3 |
|
Odonata |
37 |
4,5 |
14 |
2,2 |
36,1 |
4,3 |
|
Coenagrionidae |
24 |
2,9 |
9 |
1,4 |
15,4 |
1,9 |
|
Orthoptera |
104 |
12,7 |
56 |
8,9 |
42,1 |
5,1 |
|
Acrididae |
97 |
11,8 |
51 |
8,1 |
28,1 |
3,4 |
|
Gryllidae |
7 |
0,9 |
5 |
0,8 |
14,0 |
1,7 |
|
Homoptera |
50 |
6,1 |
19 |
3,0 |
27,7 |
3,3 |
|
Aphididae |
31 |
3,8 |
11 |
1,7 |
10,9 |
1,3 |
|
Cicadidae |
19 |
2,3 |
8 |
1,3 |
16,8 |
2,0 |
|
Heteroptera |
60 |
7,3 |
49 |
7,8 |
48,7 |
5,9 |
|
Pentatomidae |
48 |
5,9 |
31 |
4,9 |
27,4 |
3,3 |
|
Miridae |
12 |
1,5 |
18 |
2,9 |
21,3 |
2,6 |
|
Coleoptera |
312 |
38,1 |
267 |
42,3 |
217,0 |
26,1 |
|
Carabidae |
47 |
5,7 |
104 |
16,5 |
34,8 |
4,2 |
|
Количество экземпляров |
||||||
|
Кормовые объекты |
Во внешней среде |
В пищеварительном тракте |
Биомасса |
|||
|
Абс. |
% |
Абс. |
% |
Абс., мг |
% |
|
|
Scarabaeidae |
27 |
3,3 |
10 |
1,6 |
21,0 |
2,5 |
|
Sylphidae |
19 |
2,3 |
27 |
4,3 |
19,1 |
2,3 |
|
Staphylinidae |
7 |
0,9 |
12 |
1,9 |
16,5 |
2,0 |
|
Elateridae |
21 |
2,6 |
19 |
3,0 |
19,7 |
2,4 |
|
Cantharidae |
13 |
1,6 |
5 |
0,8 |
11,0 |
1,3 |
|
Coccinellidae |
34 |
4,2 |
11 |
1,7 |
18,1 |
2,2 |
|
Lagriidae |
22 |
2,7 |
9 |
1,4 |
9,8 |
1,2 |
|
Cerambycidae |
49 |
6,0 |
31 |
4,9 |
30,8 |
3,7 |
|
Chrysomelidae |
41 |
5,0 |
29 |
4,6 |
24,5 |
2,9 |
|
Curculionidae |
32 |
3,9 |
10 |
1,6 |
11,7 |
1,4 |
|
Neuroptera |
14 |
1,7 |
3 |
0,5 |
8,9 |
1,1 |
|
Hymenoptera |
50 |
6,1 |
47 |
7,4 |
68,3 |
8,2 |
|
Tenthredinidae |
9 |
1,1 |
19 |
3,0 |
18,3 |
2,2 |
|
Vespidae |
15 |
1,8 |
8 |
1,3 |
21,1 |
2,5 |
|
Apidae |
19 |
2,3 |
7 |
1,1 |
14,9 |
1,8 |
|
Formicidae |
7 |
0,9 |
13 |
2,1 |
14,0 |
1,7 |
|
Lepidoptera |
43 |
5,3 |
31 |
4,9 |
39,0 |
4,7 |
|
Lycaenidae |
19 |
2,3 |
8 |
1,3 |
13,3 |
1,6 |
|
Nymphalidae |
24 |
2,9 |
17 |
2,7 |
19,0 |
2,3 |
|
Diptera |
46 |
5,6 |
45 |
7,1 |
63,3 |
7,6 |
|
Tabanidae |
19 |
2,3 |
28 |
4,4 |
40,9 |
4,9 |
|
Syrphidae |
27 |
3,3 |
17 |
2,7 |
22,4 |
2,7 |
|
Annelida |
||||||
|
Oligochaeta |
||||||
|
Lumbricidae |
6 |
0,7 |
7 |
1,1 |
35,1 |
4,2 |
|
Mollusca |
||||||
|
Gastropoda |
||||||
|
Pulmonata |
||||||
Продолжение таблицы 1
|
1 |
2 |
3 |
4 |
5 I |
6 1 |
7 |
|
Limacidae |
9 |
1,1 |
4 |
0,6 1 |
41,3 |
1 5,0 |
|
Chordata |
59 |
7,2 |
24 |
3,8 |
55,7 |
6,7 |
|
Reptilia |
||||||
|
Squamata |
47 |
5,7 |
24 |
3,8 |
55,7 |
6,7 |
|
Lacertidae |
38 |
4,6 |
24 |
3,8 |
55,7 |
6,7 |
|
Растительность |
– |
– |
9 |
1,4 |
19,2 |
2,3 |
|
Камни |
– |
– |
8 |
1,3 |
38,7 |
4,7 |
|
Всего |
819 |
100 |
631 |
100 |
830,6 |
100 |
Восточный склон
|
Arthropoda |
1730 |
96,4 |
493 |
90,6 |
684,4 |
81,4 |
|
Arachnida |
31 |
1,7 |
10 |
1,8 |
30,7 |
3,7 |
|
Aranei |
31 |
1,7 |
10 |
1,8 |
30,7 |
3,7 |
|
Araneidae |
10 |
0,6 |
5 |
0,9 |
15,7 |
1,9 |
|
Thomisidae |
14 |
0,8 |
1 |
0,2 |
4,1 |
0,5 |
|
Lycosidae |
7 |
0,4 |
4 |
0,7 |
10,9 |
1,3 |
|
Hexapoda |
834 |
46,5 |
483 |
88,8 |
653,7 |
77,8 |
|
Odonata |
47 |
2,6 |
29 |
5,3 |
35,1 |
4,2 |
|
Coenagrionidae |
18 |
1,0 |
29 |
5,3 |
35,1 |
4,2 |
|
Количество экземпляров |
||||||
|
Кормовые объекты |
Во внешней среде |
В пищеварительном тракте |
Биомасса |
|||
|
Абс. |
% |
Абс. |
% |
Абс., мг |
% |
|
|
Orthoptera |
48 |
2,7 |
94 |
17,3 |
120,9 |
14,4 |
|
Acrididae |
42 |
2,3 |
94 |
17,3 |
120,9 |
14,4 |
|
Homoptera |
63 |
3,5 |
28 |
5,1 |
31,5 |
3,7 |
|
Aphididae |
34 |
1,9 |
8 |
1,5 |
10,5 |
1,2 |
|
Cicadidae |
29 |
1,6 |
20 |
3,7 |
21,0 |
2,5 |
|
Heteroptera |
55 |
3,1 |
29 |
5,3 |
30,4 |
3,6 |
|
Pentatomidae |
38 |
2,1 |
29 |
5,3 |
30,4 |
3,6 |
|
Coleoptera |
355 |
19,8 |
208 |
38,2 |
258 |
30,7 |
|
Carabidae |
34 |
1,9 |
11 |
2,0 |
35,1 |
4,2 |
|
Scarabaeidae |
28 |
1,6 |
18 |
3,3 |
22,9 |
2,7 |
|
Sylphidae |
9 |
0,5 |
21 |
3,9 |
20,1 |
2,4 |
|
Elateridae |
37 |
2,1 |
14 |
2,6 |
19,5 |
2,3 |
|
Coccinellidae |
44 |
2,5 |
20 |
3,7 |
31,0 |
3,7 |
|
Lagriidae |
14 |
0,8 |
12 |
2,2 |
11,0 |
1,3 |
|
Cerambycidae |
54 |
3,0 |
31 |
5,7 |
45,8 |
5,5 |
|
Chrysomelidae |
61 |
3,4 |
43 |
7,9 |
42,9 |
5,1 |
|
Curculionidae |
74 |
4,1 |
38 |
7,0 |
29,7 |
3,5 |
|
Hymenoptera |
116 |
6,5 |
39 |
7,2 |
69,8 |
8,3 |
|
Tenthredinidae |
34 |
1,9 |
12 |
2,2 |
19,4 |
2,3 |
|
Vespidae |
11 |
0,6 |
3 |
0,6 |
18,0 |
2,1 |
|
Apidae |
49 |
2,7 |
15 |
2,8 |
22,4 |
2,7 |
|
Formicidae |
22 |
1,2 |
9 |
1,7 |
10,0 |
1,2 |
|
Lepidoptera |
51 |
2,8 |
14 |
2,6 |
49,4 |
5,9 |
|
Pieridae |
32 |
1,8 |
6 |
1,1 |
20,4 |
2,4 |
|
Nymphalidae |
19 |
1,1 |
8 |
1,5 |
29,0 |
3,5 |
|
Diptera |
99 |
5,5 |
42 |
7,7 |
58,6 |
7,0 |
|
Tabanidae |
68 |
3,8 |
25 |
4,6 |
37,5 |
4,5 |
|
Syrphidae |
31 |
1,7 |
17 |
3,1 |
21,1 |
2,5 |
|
Chordata |
||||||
|
Reptilia |
||||||
|
Squamata |
65 |
3,6 |
51 |
9,4 |
97,8 |
11,6 |
|
Lacertidae |
59 |
3,3 |
51 |
9,4 |
97,8 |
11,6 |
|
Растительность |
– |
– |
4 |
0,7 |
10,3 |
1,2 |
|
Камни |
– |
– |
5 |
0,9 |
47,8 |
5,7 |
|
Всего |
1795 |
100 |
544 |
100 |
840,3 |
100 |
Исходя из результатов анализа, основу питания для популяций L. agilis Среднего Урала составляют насекомые. Они встречаются в пищеварительном тракте рептилий на протяжении всего активного периода. В целом преобладают представители жесткокрылых и прямокрылых, другие группы встречаются значительно реже. Кроме того, заметную роль в питании играют представители своего семейства и вида (особенно на восточном склоне). Кроме того, в рационе ящериц на восточном склоне чаще встречаются быстро передвигающиеся формы жертв. Это, прежде всего, двукрылые, прямокрылые, активные виды жест- кокрылых, жалящие перепончатокрылые и представители своего вида. Основную часть кормовых объектов ящерицы хватают непосредственно с поверхности почвы или с низкорослой травянистой растительности, реже добывают пищу в густом кустарнике на высоте 30-80 см от земли. В ряде случаев в пищеварительном тракте ящериц были обнаружены растительные остатки (чаще фрагменты листьев и семян), вероятно, попавшие с животной пищей, а также мелкие частицы поч- вы и камни, вероятно, играющие роль гастролитов.
В большинстве случаев частота встречаемости определенной группы кормовых животных в пи- щеварительном тракте рептилий была пропорциональна встречаемости их в природе. Однако для некоторых групп эта закономерность не наблюдалась. Так, в пище L. agilis на западном склоне характерна повышенная встречаемость пауков и жужелиц. У животных восточного склона наблюдается сходная тенденция для саранчовых, листоедов и молодых представителей своего вида. В ряде случаев некоторые группы потенциальных кормов, наряду с заметной встречаемостью в природе, не были обнаружены в рационе ящериц. Таким образом, имеет место определенная степень избирательности рептилий в питании, в том числе, предпочитаемость кормов определенного размерно-весового класса. Так, в рационе взрослых ящериц западного и восточного склона наиболее часто встречались животные размером 13-17 и 24-28 мм соответственно. Помимо прочего, выбор данного размерного класса жертвы может быть обусловлен длиной тела и ритмами активности самого хищника.
Результаты анализа дистанцированности спектров питания с учетом количественного и качест- венного состава корма выделяют некоторую специфику рациона ящериц западного склона (рис 1).
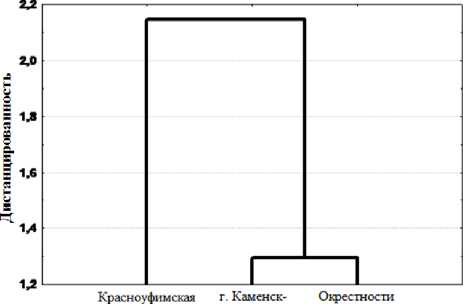
лесостепь Уральский ст. Кунавино
Локалнтеты
Рис. 1. Дистанцированность популяций L. agilis по спектрам питания
Отмечено, что спектр питания рептилий на западном склоне несколько шире, чем на восточном - за счет присутствия в рационе ящериц таких групп кормов, как: клещи, сетчатокрылые, наземные моллюски и дождевые черви. Тем не менее, в отношении большинства групп кормов, спектры питания взрослых ящериц на западном и восточном склонах достаточно сходны и перекрываются на 79,4% (табл. 2). Известно, что чем ниже таксономический ранг кормовых объектов, тем меньше степень перекрывания их потребителей [3]. Исходя из этой закономерности, перекрывание спектров оценивали на уровне отрядов и семейств. На видовом уровне спектры питания L. agilis могут различаться в гораздо большей степени, что является возможным следствием как особенностей локального фаунистического состава кормов в биотопах, так и определенной избирательности ящериц при кормодобывании.
Таблица 2. Перекрывание спектров питания L. agilis на западном и восточном склонах по модифицированному индексу Мориситы
|
Географические районы |
Западный склон |
Восточный склон |
|
Западный склон |
////////////////////////// |
0,794 |
|
Восточный склон |
0,794 |
////////////////////////// |
Учет биомассы кормовых животных в пищеварительном тракте рептилий в течение сезона позволил оценить величину суточного рациона ящериц. На протяжении всего активного периода данная величина может колебаться в широких пределах. В первую очередь, она сопряжена с суточными и сезонными ритмами активности ящериц, а также с пространственно-временными особенностями жизненного цикла кормовых объектов. Несмотря на индивидуальные вариации, сезонная динамика суточного рациона подчиняется определенной общей закономерности. Эта закономерность неразрывно связана с метеоусловиями на протяжении сезона. Так, наибольшая масса суточного рациона наблюдается во второй половине июня (на восточном склоне – 2,8 г) и в конце июня – начале июля (на западном склоне – 2,4 г). В этот период, после спаривания, животные наиболее активно и регулярно питаются, запасая энергию для компенсации ее потерь в репродуктивный период. Часто комплекс внешних условий (температура воздуха и почвы, влажность, доля солнечного УФ-излучения и др.) также благоприятен для наиболее полного извлечения энергии при усвоении корма. С апреля по конец мая, а также с начала августа по сентябрь ящерицы питаются неравномерно и величина суточного рациона при этом заметно ниже (в среднем, на западном и восточном склоне 1,6 и 1,9 г соответственно). Сравнительный анализ суточного рациона самцов и самок не обнаруживает существенных различий в массе, однако, качественный состав корма самцов (в особенности, на западном склоне) более разнообразен: сетчатокрылые, моллюски и олигохеты были обнаружены именно в составе их рациона. Отмечено, что степень активности и агрессивности крупных территориальных самцов значительно выше, чем у самок. При этом самцы расходуют значительное количество энергии и более склонны нападать на представителей своего вида и пола, а также на крупную добычу.
Обобщая полученные сведения о питании L. agilis Среднего Урала, можно отметить общее сходство качественного состава кормов на западном и восточном склонах. Доминирующую роль в питании играют жесткокрылые и прямокрылые. Существует определенная избирательность в рационе, сопряженная с рядом особенностей рептилий и кормовых объектов. При описании аспектов питания L. agilis как активного хищника, помимо состава кормов, необходимо учитывать популяционную динамику их потребления, а также величину суточного рациона – для более полного понимания роли прыткой ящерицы в регуляции численности кормовых объектов, которые отличаются большим разнообразием.
Список литературы Экология питания прыткой ящерицы (Lacerta agilis L.) в популяциях Среднего Урала
- Вершинин В.Л. Амфибии и рептилии Урала. Екатеринбург: УрО РАН, 2007. 170 с.
- Даревский И.С. и др. Систематика и внутривидовая структура//Прыткая ящерица. Монографическое описание вида . М.: Наука, 1976. С. 53-92.
- Кузьмин С.Л. Трофология хвостатых земноводных: экологические и эволюционные аспекты. М.: Наука, 1992. 167 с.
- Лакин Г.Ф. Биометрия. М: Высш. школа, 1990. 352 с.
- Малоземов Ю.А., Малоземова Л.А. Краткий определитель беспозвоночных животных Среднего Урала. Екатеринбург: Изд-во Урал. ун-та, 2005. 277 с.
- Одум Ю. Экология: в 2 т. Т. 2. М.: Мир, 1986. 376 с.
- Определитель насекомых европейской части СССР: в 5 т. Т. 2. Жесткокрылые и веерокрылые М.-Л.: Наука, 1965. 668 с.
- Песенко Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 1982. 288 с.
- Пианка Э. Эволюционная экология. М.: Мир, 1981. 400 с.
- Плавильщиков Н.Н. Определитель насекомых. М, 1994. 544 с.
- Прыткая ящерица. Монографическое описание вида . М: Наука, 1976. 384 с.
- Шляхтин Г.В., Голикова В.Л. Методика полевых исследований экологии амфибий и рептилий. Саратов: Изд-во Сарат. ун-та, 1986. 78 с.
- Hurlbert S.H. The measurement of niche overlap and some relatives//Ecology. 1978. Vol. 59, № 1. P. 67-77.
