Эколого-биологическая характеристика сибирского осетра (Acipenser baeri Brandt, 1869) в озере Байкал
 в озере Байкал")
Автор: Афанасьев С.Г.
Журнал: Природа Внутренней Азии @nature-inner-asia
Рубрика: Биология
Статья в выпуске: 1 (30), 2025 года.
Бесплатный доступ
Осетровые являются уникальной группой древнейших рыб, а также ценными промысловыми видами, хищнический вылов которых привел к катастрофическому сокращению численности на всем протяжении ареала. Целью исследований было изучение особенностей распространения, роста и питания сибирского осетра (Acipenser baeri Brandt, 1869), редкого исчезающего вида в реке Селенге и оз. Байкал. Показано, что в летний период распределение байкальского осетра в этих водоемах определяется особенностями донных ландшафтов. Места нереста производителей расположены на всем протяжении р. Селенги - от устья до ее верхних участков. При достижении половозрелого возраста рост в длину у байкальского осетра постепенно замедляется, а масса тела продолжает увеличиваться. Достоверных различий по длине и массе тела между самцами и самками одной возрастной группы осетра не обнаружено. Выявлена прямая корреляционная зависимость абсолютной плодовитости самок от их возраста (r = 0,71, p=0,05) и массы тела (r= 0,83, p=0,05). Относительная плодовитость снижалась по мере роста линейно-весовых показателей, а также упитанности самок. Отмечено, что байкальский осетр демонстрирует избирательность в питании, что напрямую связано с местом его обитания и сезоном года. В нагульный период на Селенгинском мелководье, в заливе Провал и дельте р. Селенги в рационе его питания преобладают амфиподы и хирономиды, а при нагуле в реке - личинки амфибиотических насекомых и молодь рыб. На основе индексов наполнения желудков сделан вывод о наличии обильной кормовой базы в Селенгинском мелководье и ее дельтовых пространствах.
Байкальская популяция, оз. байкал, распространение, линейно-весовой рост, плодовитость, питание
Короткий адрес: https://sciup.org/148331745
IDR: 148331745 | УДК: 597.423 | DOI: 10.18101/2542-0623-2025-1-6-19
Ecological and biological characteristics of the Siberian sturgeon (Acipenser baeri Brandt, 1869) in Lake Baikal
Sturgeons are a unique group of ancient fish, as well as valuable commercial species, predatory fishing of which has led to a catastrophic reduction in numbers throughout their range. The research is aimed at studying the distribution, growth and feeding characteristics of the Siberian sturgeon (Acipenser baeri Brandt, 1869), a rare endangered species in the Selenga River and Lake Baikal. It has shown that in summer the distribution of the Baikal sturgeon in these waterbodies is determined by the features of the bottom landscapes. The spawning grounds of brood fish are located along the entire length of the Selenga River - from the mouth to its upper sections. Upon reaching breeding age, the growth in length of the Baikal sturgeon gradually slows down, and the body weight continues to increase. We havn't found a reliable differences in length and body weight of male and female individuals of the same age group of sturgeon. However, there is a direct correlation of the absolute fecundity of females with their age (r=0,71, p=0,05) and body weight (r=0,83, p=0,05). Relative fecundity decreases as the linear weight indicators increases, as well as the fatness of the females. It is noted that the Baikal sturgeon demonstrates selectivity in feeding, which is directly related to its habitat and season. During the feeding period in the Selenga shallow waters, the Proval Bay and the Selenga River delta its diet is dominated by amphipods and chironomids, during fattening in the river it is dominated by larvae of amphibiotic insects and young fish. Based on the stomach fullness indices, we have concluded that there is an abundant food supply in the Selenga shallow waters and its delta.
Текст научной статьи Эколого-биологическая характеристика сибирского осетра (Acipenser baeri Brandt, 1869) в озере Байкал
Представители семейства Acipenseridae (Осетровые) являются уникальной группой древнейших рыб, способной обитать в различных экологических условиях. Они населяют пресные и соленые водоемы субтропических, умеренных и субарктических зон Евразии и Северной Америки, создавая локальные популяции или сезонные расы. В ходе эволюционного развития осетровые рыбы приобрели целый ряд анатомо-морфологических и физиологических особенностей, что дает преимущество перед костистыми рыбами. Одной из них является наличие жучек, образующих наружный скелет и обеспечивающих защиту от хищников. Кроме того, половые продукты осетровых рыб характеризуются длительной способностью к оплодотворению, а нерест может происходить многократно и в широком температурном диапазоне. К адаптациям, обеспечивающим выживаемость личинок сибирского осетра на ранних этапах онтогенеза, относятся эмбриональная эвритермность, ускоренное развитие личинок в постэмбриональный период, формирование крупного желточного мешка на ранних стадиях развития личинок, что обеспечивает их продолжительный скат без дополнительного внешнего питания [Афанасьев, 2006; Рубан, 2019].
Несмотря на эти преимущества, осетровые рыбы, являясь ценными промысловыми видами, столкнулись с серьезной угрозой исчезновения из-за катастрофического сокращения численности на всем протяжении ареала в результате хищнического вылова, особенно в конце XIX — начале XX в. Например, промысел сибирского осетра ( Acipenser baeri Brandt, 1869) во время нерестовой миграции в оз. Байкал в начале прошлого века, в сочетании с массовым истреблением молоди, привел к катастрофическим последствиям для популяции. В 1924 г. общий вылов осетра в Баргузинском и Верхнеудинском районах едва достиг четырех тонн, что свидетельствует о резком падении его численности [Егоров, 1961]. В результате в 1930–1935 гг. был введен запрет на промысел осетра в оз. Байкал, однако это не привело к заметному восстановлению численности популяции и уже в 1945 г. запрет был возобновлен и продолжает действовать до сих пор.
К сожалению, снижение численности осетровых связано не только с выловом, но и ухудшением качества водоемов, изменением их гидрологических режимов и разрушением мест нереста. Так, строительство Иркутской ГЭС, Селенгинского картонного комбината, изменение русел рек, впадающих в оз. Байкал, добыча песчано-гравийных смесей оказали существенное влияние на миграционные пути и места нерестилищ сибирского осетра [Афанасьев, 2006]. В начале 1980-х гг. было принято решение об искусственном воспроизводстве вида, однако даже такая мера поддержки не привела к восстановлению его промыслового запаса: численность сибирского осетра на Селенгинском мелководье составляла примерно 18 тыс. экземпляров, в Баргузинском заливе — 4 тыс., в р. Селенге — 140 [Афанасьев, 1997]. Из-за низкой численности сибирский осетр байкальской популяции (далее — байкальский осетр) был занесен в красные книги России (1987) и Республики Бурятия (1988) как редкий исчезающий вид.
Следует отметить, что статус редкого вида затрудняет получение актуальных данных об экологии и биологии байкальского осетра. В настоящее время это трудоемкая, но очень важная задача, особенно при разработке мероприятий по поддержанию и увеличению его численности в естественных условиях. Учитывая отсутствие современных данных о состоянии байкальской популяций сибирского осетра, целью настоящей работы было изучение особенностей его распространения, роста и питания в реке Селенге и оз. Байкал.
Объект и методы исследования
Объектом исследования являлся сибирский осетр байкальской популяции (или байкальский осетр, изолированная популяция, обитающая в оз. Байкал и его крупных притоках). Сбор материала по экологии и биологии байкальского осетра осуществлялся в р. Селенге и оз. Байкал (дельта, авандельта, Селенгинское мелководье, залив Провал) (рис. 1).
Отлов осетра на Селенгинском мелководье оз. Байкал проводили донными ставными сетями общей длиной 400 м и размером ячеи 20–200 мм. В р. Селенге в качестве орудий лова применяли плавные сети с ячеей 14, 30, 36 и 40 мм общей длиной 65–70 м. Особенности миграций разновозрастного осетра в период летнего нагула исследовали путем его мечения [Рикер, 1979]. Гидростатические метки с номером крепились на грудной или спинной плавник, места поимки и выпуска осетра после мечения регистрировались. Во всех местах проведения работ велось наблюдение за температурным режимом.
Для характеристики кормовой базы осетра отбирались пробы зообентоса в основных местах его обитания и на всем протяжении путей ската искусственно выращенной молоди. Отбор проб осуществлялся дночерпателем Петерсена (площадь захвата 0,025 м2), бентометром Леванидова (площадь захвата 0,0625 м2).
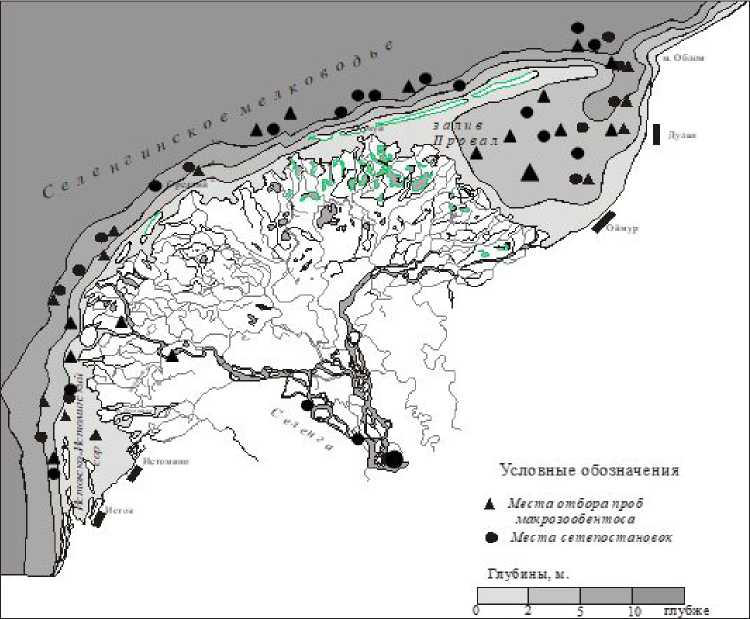
Рис. 1. Карта-схема мест сетепостановок и отбора проб зообентоса в р. Селенге и оз. Байкал
Статистическую обработку данных проводили с использованием стандартных методов [Лакин, 1990] и пакета программ Microsoft Office 2013 и Statistica 11.0.
Результаты и обсуждение
Распространение и миграции
Для оценки численности популяции рыб и изучения их миграционных путей широко используется метод мечения. Данные, полученные с помощью этого метода, показали, что основными местами обитания байкальского осетра являются р. Селенга, ее обширная дельта, Селенгинское мелководье и прилегающий к ним залив Провал.
В летний период распределение байкальского осетра в этих водоемах во многом определяется особенностями донных ландшафтов. Скопление рыб было зафиксировано преимущественно на песчано-илистых и илистых грунтах, где отмечается наиболее высокая плотность зообентоса, являющегося одним из основных источников питания для осетра. Интересно, что разновозрастный осетр в нагульный период демонстрирует определенные предпочтения в выборе мест обитания. В отличие от большинства представителей прибрежно-соровых сообществ рыб он выбирает свободные от высшей водной растительности открытые пространства и избегает заросших участков, причем чем крупнее особь, тем реже она заплывает в заросли. Вероятно, густая водная растительность затрудняет поиск пищи, а также ориентацию в пространстве. Важно также отметить, что байкальский осетр может вести оседлый образ жизни, о чем свидетельствуют результаты повторного вылова меченых рыб: некоторые особи были пойманы через год на расстоянии 1–30 км от места их первоначального выпуска. Это свидетельствует о том, что осетры могут оставаться в определенной зоне продолжительное время, что позволяет им эффективно использовать пищевые ресурсы. Когда температура воды начинает понижаться до 8–10 °С, байкальский осетр перемещается в оз. Байкал, где зимует на глубине от 20 до 40 метров.
Нерестовая миграция байкальского осетра также представляет собой важный аспект его жизненного цикла. Этот процесс начинается, как правило, в начале весны и продолжается до начала лета. Анализ литературных данных, а также собственные наблюдения за расположением нерестилищ байкальского осетра позволяют сделать заключение о том, что его нерестовая миграция в р. Селенге происходит по протокам Харауз, Тугариха, Лобановская, Толстоножиха, Средняя (в период высокого уровня воды), Хаустик, Новый и Нижний промой, Красноярская. О том, что байкальский осетр нерестится на нижних (30–40 км от устья), средних и верхних (180–400 км) участках р. Селенги, свидетельствуют случаи поимки здесь производителей с текучими гонадами (5-я стадия зрелости).
Рост, созревание и плодовитость байкальского осетра в естественных условиях
Среди осетровых рыб в естественных условиях обитания сибирский осетр — это один из наиболее медленно растущих видов, при этом особи могут достигать значительной массы. Известны случаи поимки обского осетра массой 180–200 кг [Берг, 1949; Дрягин, 1949] и байкальского осетра массой 150–200 кг [Егоров, 1961].
Как известно, интенсивность роста осетра тесно связана с экологическими условиями, преимущественно с температурным режимом водоемов и их трофно-стью. Так, в условиях искусственного разведения в бассейнах, где температура воды и обеспеченность пищей гораздо выше по сравнению с естественной средой, темпы роста у сибирского осетра становятся в 7–9 раз выше [Смольянов, 1987]. Байкальский осетр при выращивании в садковом хозяйстве Гусиноозерской ГРЭС за первый год содержания достигал массы почти четыреста грамм, что примерно в четыре раза больше, чем в естественных водоемах [Афанасьев, 2006].
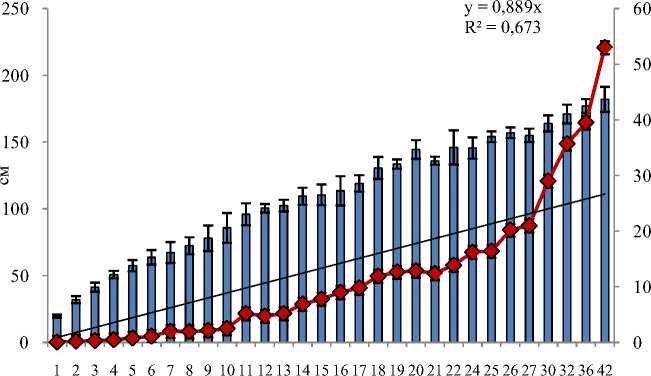
Анализ полученных нами данных показал, что в естественной среде обитания интенсивный прирост в длину у особей байкальского осетра наблюдается в период 1–17 лет (до половозрелого возраста), затем он постепенно замедляется, но рыбы продолжают расти, хоть и медленно (рис. 2). Средняя длина половозрелых особей может достигать 1,5–1,8 м. Значение массы тела также увеличивается в первые годы жизни, затем несмотря на замедление линейного роста масса особей продолжает расти и к 40 годам может достигать 60 кг. Коэффициент вариации линейно-весовых показателей в пределах возрастных групп имеет высокие значения, что может быть связано с длительным периодом нереста, т. е. сеголетки поздненере-стующих рыб по своим размерам бывают меньше сеголетков рыб, нерестующих рано весной. С другой стороны, молодь осетра и взрослые рыбы, распространяясь по водоему, попадают в различные по экологическим факторам (температура, плотность бентоса, насыщенность воды кислородом и др.) условия обитания, что также влияет на уровень варьирования линейно-весовых показателей. Следует также отметить отсутствие достоверных различий по длине и массе между самцами и самками одной возрастной группы.
Анализ наших данных, а также фондовых материалов за 1972–2011 гг. показал, что половозрелость самцов байкальского осетра наступает в 13–15 лет, самок — 16–18 лет. Показатели упитанности по Фультону у обоих полов половозрелого осетра сопоставимы — 0,5–0,7 у самок и 0,5–0,8 у самцов. Упитанность молоди осетра в возрасте 0+– 3+ в нагульный период также была высокой и находилась в пределах 0,6–0,8.
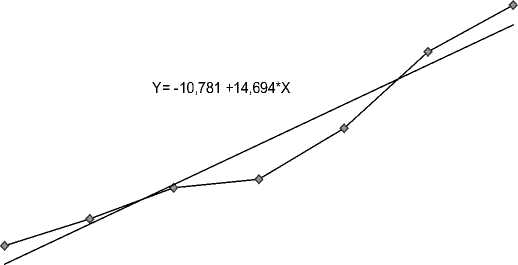
Важным показателем репродуктивной способности самок рыб является их плодовитость. Известно, что абсолютная плодовитость (АП, число ооцитов/ самка) пропорциональна массе рыбы, поэтому этот показатель может значительно варьировать даже в пределах одной возрастной группы. Так, у самок байкальского осетра, выловленных на Селенгинском мелководье, в заливе Провал и р. Селенге в период преднерестовых и нерестовых миграций, АП изменялась в довольно широких пределах (180–740 тыс. шт.) (рис. 3), что соответствует литературным данным, приводимым для этого вида (210–830 тыс. шт.) [Егоров, 1961]. Установлена достоверная прямая корреляционная зависимость этого показателя от возраста самок (r=0,71, p=0,05) и их массы тела (r=0,83, p=0,05).
Возраст, лет
Длина Масса Линейная (Масса)
Рис. 2. Возрастное изменение длины и массы тела байкальского осетра в естественной среде обитания
I
I 600
H
£ 500
о о
I 400
z
§ 100
10-14,9 15-19,9 20-24,9 25-29,9 30 - 34,9 35 - 40 53
Масса, кг
Рис. 3. Зависимость абсолютной плодовитости самок байкальского осетра от массы тела
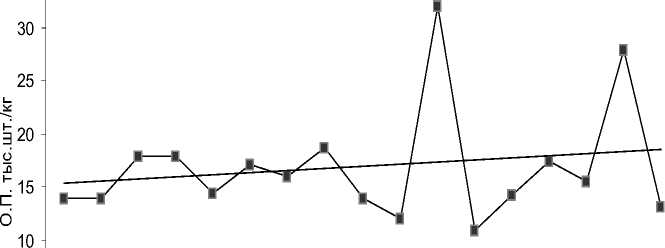
Более постоянной величиной считается относительная плодовитость (ОП, количество ооцитов, приходящихся на единицу массы тела). У самок байкальского осетра этот показатель варьировал в пределах 9,4–32,4 тыс. шт./кг (рис. 4), причем ОП в меньшей степени зависела от возраста особей (r=0,21, p=0,05) по сравнению с АП. Установлено также, что более высокие темпы роста ОП характерны для возрастного диапазона 20–30 лет, у старших возрастных групп этот показатель не обнаруживает достоверных различий. Полученные нами результаты свидетельствуют о том, что при увеличении АП по мере роста линейно-весовых показателей, а также упитанности самок происходит постепенное снижение ОП, что согласуется с данными других авторов [Егоров, 1961; Беспалова, Кошелев, 2007; Беляев и др., 2022].
18 20 21 22 23 25 26 27 28 29 30 31 32 33 34 36 38
Возраст, лет
Рис. 4. Зависимость относительной плодовитости самок байкальского осетра от возраста
Питание байкальского осетра
Анализ литературных источников показал, что сведения о питании байкальского осетра, учитывая его статус редкого вида, фрагментарны и единичны. Из данных А. Г. Егорова (1961) следует, что байкальский осетр является типичным бентофагом, характер питания которого зависит от видового разнообразия зообентоса и его численности. Считается, что основными компонентами пищи осетра являются амфиподы, личинки амфибиотических насекомых, широколобки.
Известно, что важным показателем, определяющим численность популяций промысловых рыб, является состояние кормовой базы водоемов, в которых они обитают. Количественная и качественная оценка зообентоса в р. Селенге показала, что основу донного комплекса беспозвоночных животных слагают личинки амфибиотических насекомых четырех отрядов: поденки, веснянки, ручейники и двукрылые [Базова, 2004]. В продольном профиле реки минимальные показатели зообентоса (средняя биомасса 0,18 г/м2) отмечены в ее верхнем участке, что может быть связано с господством здесь больших скоростей течения и легко переносимых песчаных и галечных грунтов. Средний участок реки, где доминируют каменисто-галечные грунты и также наблюдаются высокие скорости течения, донной фауной заселен неравномерно (средняя биомасса 1,07 г/м2). Максимальные значения биомассы зообентоса обнаружены на нижних участках реки, а также в дельте и заливе Провал, где преобладают малые скорости течения и гравийнокаменистые и заиленные грунты. Доминирующей группой зообентоса на нижних участках реки по численности являются личинки двукрылых (38%), по биомассе – личинки ручейников (37%), а в дельте и заливе Провал – олигохеты, хирономиды и амфиподы [Базова, 2004; Рожкова и др., 2016; Колпакова и др., 2021]. Биомасса бентоса варьирует от 0,64 до 44,02 г/ м2 при среднем значении 4,46 г/м2 в нижней части р. Селенги и 18,12 г/м2—на илистых биотопах залива Провал.
Примечание: *здесь и в таблице 2 ... — отсутствуют в пищевом комке.
Состав пищи байкальского осетра в р. Селенге и заливе Провал в летний период
Таблица 2
Питание молоди байкальского осетра на нижних участках р. Селенги в летний и осенний период
Полученные нами данные свидетельствуют о том, что особи байкальского осетра младших возрастов отдают предпочтение личинкам амфибиотических насекомых (рис. 3). По мере увеличения возраста и линейно-весовых размеров (примерно до 60 см) в рационе увеличивается доля личинок поденок и снижается доля хирономид. У старших возрастных групп длиной тела свыше 80 см основным компонентом питания становится рыба, а доля амфибиотических насекомых резко сокращается. Как и другие виды осетровых рыб, байкальский осетр проявляет избирательность в питании в зависимости от возрастного состояния. Переключение питания с бентосных организмов на рыбный рацион, а также питание теми и другими одновременно в зависимости от их наличия и доступности отражает пластичность и адаптационные возможности байкальского осетра.
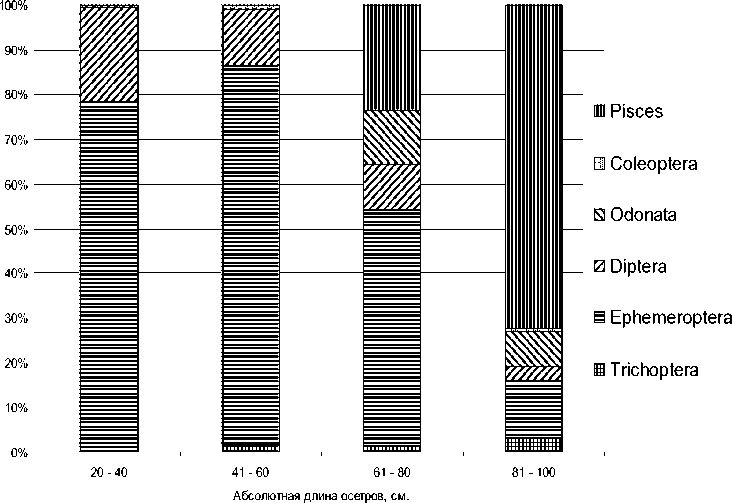
Рис. 3. Пищевая избирательность байкальского осетра в зависимости от его линейного размера в дельте р. Селенги (% по массе)
Заключение
В результате проведенных исследований получены данные о распространении, росте и питании байкальского осетра в р. Селенге и оз. Байкал. Места нереста производителей сибирского осетра байкальской популяции расположены на всем протяжении р. Селенги — от устья (20 км) до ее верхних участков.
При достижении половозрелого возраста рост в длину у байкальского осетра постепенно замедляется, а масса тела продолжает увеличиваться. Достоверных различий по длине и массе тела между самцами и самками одной возрастной группы осетра не обнаружено.
Установлена прямая корреляционная зависимость абсолютной плодовитости самок от их возраста (r=0,71, p=0,05) и массы тела (r=0,83, p=0,05). Относительная плодовитость снижалась по мере роста линейно-весовых показателей, а также упитанности самок.
Установлено, что в нагульный период в заливе Провал, Селенгинском мелководье оз. Байкал осетр предпочитает наиболее продуктивные песчано-илистые и илистые биотопы с низкой скоростью течения. Подобно сибирскому осетру на большей части его ареала байкальский осетр демонстрирует избирательность в питании, что напрямую связано с местом его обитания и сезоном года. В нагульный период на Селенгинском мелководье, в заливе Провал и дельте р. Селенги в рационе его питания преобладают амфиподы и хирономиды, а при нагуле в реке — личинки амфибиотических насекомых и молодь рыб.
