Эколого-биологические особенности раннецветущих растений Восточного Забайкалья

Автор: Попова О.А., Якимова Е.П., Лескова О.А.
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Ботаника
Статья в выпуске: 4, 2008 года.
Бесплатный доступ
Раннецветущие растения - это группа видов, присопосбленных к росту и развитию в экстремальных условиях. Благоприятные температурные условия, вынужденный зимний покой, резервное накопление питательных веществ, водные условия способствуют раннему цветению раннецветущих растений.
Короткий адрес: https://sciup.org/148178296
IDR: 148178296
Ecology-biological feutures sprining plants of East Transbaikalia
Early blooming plants are species group adjusted to growth and development in extreme habitat conditions. Favorable temperature conditions, forced winter rest, reserve nutrients accumulation peculiarities, water conditions peculiarities promote the early growth of the early blooming plants.
Текст научной статьи Эколого-биологические особенности раннецветущих растений Восточного Забайкалья
-
1. Растительные ресурсы СССР: Цветковые растения, их химический состав, использование; Семейства Rutaceae — Elaeangnaceae. Л., 1988. 357 с.
-
2. Работнова Т.А, Вопросы изучении состава популяций для целей фитоценологии // Проблемы ботаники. М», 19506. Вып. I. С. 112-164.
-
3» Уранов А.А. Возрастной спектр фитоценопо-пуляций как функция времени и энергетических волновых процессов // Науч. докл. Высш, школы: Биол. науки. 1975. Т.79. Вып. 5. С 23-27.
-
4. Беццеман НН. Методика фенологических наблюдений при геоботаничесикх исследованиях. М.-Л., 1954. 130 с.
-
5. Вайнагий И.В. Методика статистической обработки материалов по семенной продуктивности растений на примере Potentilla aurea L. // Растит, ресурсы. 1973- Т.9. Вып. 2, С, 2 87-296.
-
6. Голубев В.И. Эколого-биологические особенности травянистых растений и растительных сообществ лесостепи. Мд Наука, 1965,
О. А Попова, Е»П. Якимова, ОЛ, Лескова
Забайкальский государственный гуманитарно-педагогический университет им. Я. Г Чернышевского
ЭКОЛОГО-БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАННЕЦВЕТУЩИХ РАСТЕНИЙ ВОСТОЧНОГО ЗАБАЙКАЛЬЯ
Раннецветущие растения - это группа видов, присопосбленных к росту и развитию в экстремальных условиях. Благоприятные температурные условия» вынужденный зимний покой, резервное накопление питательных веществ, водные условия способствуют раннему цветению раннецветущих растений.
ОЛ. Popova, Е.Р. Yakimova, ОЛ. Leskova
ECOLOGY-BIOLOGICAL FEUTURES SPRINING PLANTS OF EAST TRANSBAIKALIA
Early blooming plants are species group adjusted to growth and development in extreme habitat conditions. Favorable temperature conditions, forced winter rest, reserve nutrients accumulation peculiarities, water conditions peculiarities promote the early growth of the early blooming plants.
Условия ранней и средней весны Забайкалья с их частыми заморозками и резкими суточными колебаниями температур воздуха и почвы, а также незначительное количество зимних и весенних осадков неблагоприятны для роста и развития растений. Именно в этот период растут и развиваются раннецветущие растения, которые адаптированы к вегетации в экстремальных условиях среды. Система адаптаций растений к условиям среды комплексна и включает как морфологические изменения биоморфы, так и физиологическую и биохимическую пластичность растений.
Для исследований были взяты 22 вида раннецветущих растений, обитающие в трех различных сообществах. В арктогероново-твердоватоосоковой степи были взяты следующие виды: Leibnitzia anandria (L.) Turcz., Arctogeron gramineum (L.) DC., Gagea pauciflora (Turcz.ex Trautv.) Ledeb., Pulsatilla turczaninovii Krylov et Serg, Androsace incana
Lam, Potentilla leucophylla Pallas, Alyssum obovatum (C.A. Meyer) Turcz., Euphorbia karoi Meinsh, Stellaria cherleriae (Fischer) Will., Eremogone capillaris (Poiret) Fenzl, Thlaspi cochleariforme DC. В разнотравношмидтоосо-ковом лугу: Ranunculus propinquus С A. Meyer, R.rigescens Turcz. ex Ovcz., Primula farinosa L., P, nutans Georgi, Ciminalis aquatica (L.) Zuev. В рододендроново-разнотравном сосновом лесу: Fragaria orientalis Losinsk, Potentilla fragarioides L., Pyrola asarifolia Michaux, Vaccinium vitis-idaea (L.) Avrorin, Oxytropis myriophylla (Pallas) DC., Pulsatilla multifida (Pritzel) Juz
Фенологические наблюдения проводились в 2001-2004 гг. согласно общепринятой методике (Бейдеман, 1960); содержание воды в тканях, фракционный состав воды - с использованием водоотнимающих веществ (Сулейманов, 1964, 1972). Характер покоя определяли, опираясь на работы П.А. Генкель, Б.Э.
Окниной (1964), Т.К. Горышиной (1965); ассимиляционные и запасные вещества исследовали гистохимически (Фурст, 1979).
Результаты изучения фенологического развития раннецветущих растений показали, что среди исследованных видов преобладают дли-тельновегетирующие виды - 18 видов из 22. Среди длительновегетирующих 8 (38%) зимнезеленых видов с весенним и весенне-летним циклом развития. При изучении биологии раннецветущих растений было выявлено, что среди данных видов преобладает розеточная и полурозеточная форма роста - 14 видов (64%). Образование розетки связано с торможением роста междоузлий под влиянием неблагоприятных климатических факторов и имеет большое адаптационное значение. Отсутствие вытянутого главного побега у розеточных растений способствует более рациональному использованию небольшого запаса пластических веществ, росту и развитию в более теплом приземном слое воздуха (Федоскин, 1975; Копытова, 2003). Среди экологических групп преобладают петроксерофиты - 10 видов (45%) и мезофиты - 7 видов (32%). По данным А.А Горшковой (1966, 1971), именно петроксерофиты хорошо приспособлены к сухости воздуха, недостатку влаги в почве и резким сменам температур. Сроки зацветания растений зависят от сроков заложения почек возобновления. По степени сформированное™ почек обращает на себя внимание высокий процент видов, принадлежащих к 1 группе (по классификации И.Г. Серебрякова) - 16 видов из 22. Это растения, в почках возобновления которых побеги будущего года сформированы полностью, включая соцветие и отдельные цветки. В течение весны происходит лишь раскрытие почки и рост побега, уже заложенного предыдущим летом.
Выход раннецветущих растений на поверхность почвы и начало вегетации связаны с подъемом температур воздуха. В результате исследований было обнаружено, что самое раннее развитие растений характерно для степного сообщества. Начало цветения исследованных растений степи обычно наблюдается в конце апреля, когда устанавливаются благоприятные температурные условия. Следует отметить, что в 2004 г. для Pulsatilla turczaninovii и Euphorbia karoi начало цветения было зафиксировано уже 17 апреля. В лесном сообществе начало цветения растений наблюдается в первой декаде мая. Позднее начинается развитие раннецветущих растений в луговом сообществе. Начало цветения исследованных видов отмечается во второй декаде мая, что связано с наименее благоприятными микроклиматическими условиями.
Начало развития и цветения растений зависят и от типа покоя, с которым они уходят в зиму, В литературе отмечают два типа покоя, органический, или глубокий, и вынужденный. Вынужденный покой обусловлен влиянием внешних воздействий, в частности температурой среды. Органический, или глубокий, покой характеризуется изменением обмена веществ, прекращением роста, накоплением запасных веществ (Щербаков, 1960; Генкель, Окнина, 1982). В результате наших исследований были выделены две группы растений: с вынужденным зимним покоем - сюда относится 21 вид исследованных раннецветущих растений, и с органическим зимним покоем -сюда относится один вид - Leibnitzia anandria. В лабораторных условиях растения первой группы при осеннем внесении непрерывно вегетируют или отрастают в течение ближайшей недели. Для данных видов характерно отсутствие периода глубокого органического покоя, зимний период покоя является вынужденным. Вторая группа объединяет растения, у которых рост побегов начинается через 60120 дней после снятия отрицательных температур. Если зимний период покоя для большинства раннецветущих растений является вынужденным, то совершенно иной характер имеет летний период покоя, который отмечен нами у Gagea pauciflora, Euphorbia karoi, Thiaspi cochleariforme.
Gagea pauciflora - луковичное эфемероид-ное растение. Начинает вегетировать в конце апреля, цветет в мае, К середине июня происходит обсеменение и к концу этого месяца надземный побег отмирает. Растение переходит в период глубокого летнего органического покоя, который длится до середины августа. Затем наблюдается развитие молодого побега, который отмирает с наступлением заморозков. Зимний покой является вынужденным. То есть для этого вида характерно наличие двух типов покоя: летнего - органического и зимнего - вынужденного.
Thiaspi cochleariforme - небольшое эфеме-роидное растение, начинает вегетировать в середине апреля. В мае отмечается массовое цветение, которое продолжается весь май и захватывает первую декаду июня. Затем быстро происходит созревание семян и обсеменение, а к концу июня надземные побеги от- мирают, Растение вступает в летний органический покой. Во второй половине августа ярутка вновь отрастает и уходит в зиму с зеленой розеткой прикорневых листьев. То есть у данного вида отмечено два вида покоя: летний - органический и зимний - вынужденный.
Euphorbia karoi - небольшое эфемероидное растение, начинает вегетировать в начале апреля и уже через 7-10 дней после начала вегетации зацветает. Цветение наблюдается весь апрель и май. В конце мая - начале июня созревают семена и происходит обсеменение. К концу июня надземные побеги отмирают. Следует отметить, что данный вид не отрастает осенью, то есть органический покой охватывает летние и осенние месяцы. Зимний покой является вынужденным. У молочая Кара выделены два типа покоя: органический -летне-осенний и вынужденный - зимний.
Современными климатическими условиями совершенно невозможно объяснить наличие летнего периода покоя у Gagea pauciflora, Thlaspi cochleariforme, Euphorbia karoi, так как вторая половина лета наиболее благоприятна для вегетации растений. В этот период максимальные летние температуры совпадают с максимумом выпадения осадков, что является характерной климатической особенно стью степей вообще. Вполне возможно, что летний период покоя у данных видов связан с далеким климатическим прошлым этого района и указывает. на особенности бывшего климата в летний период (Федоскин, 1974).
Важное значение в устойчивости растений к экстремальным условиям обитания имеет накопление и превращение запасных веществ. В растениях распространенными запасными веществами являются крахмал, растворимые сахара и липиды. Образование крахмала и жиров сопровождается обезвоживанием клеток и способствует перезимовке растений. Установлено, что крахмал гидролизуется с образованием сахаров, главным образом моносахаров, и в более суровых условиях - дисахаров. Наши исследования показали, что немногие растения (Leibnitzia anandria, Euphorbia karoi, Gagea pauciflora, Potentilla leucophylla, Alyssum obovatum) осенью в тканях накапливают крахмал, который затем (октябрь - ноябрь) превращается в сахара и жир. В зиму все исследованные растения уходят с сахарами и липидами. Весной, к моменту активного роста и развития растения, в исследуемых органах растений обнаруживаются сахара (табл. 1).
Таблица 1
Особенности накопления н превращения запасных веществ у исследованных видов
|
Группы видов по содержанию крахмала в осенне-зимний период |
Виды ' |
|
I. Ненакапливающие крахмал |
Eremogone capiitaris, Arctogeran gramineum, Androsace incana, Pulsatilla turczaninovii, P. mullifida, Steliaria cherleriae, Oxytropis myriophyila, Pyrola asarifolia, Vaccinium vitis-idaea, Fragaria orientaiis, Potentilla fragarioides. Primula farinosa, P. nutans, Ranunculus rigescens, R. propinquus |
|
II. Накапливающие крахмал, но быстро его превращающие |
Leibnitzia anandria, Euphorbia karoi, Gagea pauciflora, Potentilla leucophylla. Alyssum obovatum |
Одним из физиологических факторов, обеспечивающих устойчивость растений к неблагоприятным факторам среды, является состояние водного режима их тканей. Было установлено (рис. 1), что самое высокое содержание воды в листьях (более 80% сырой массы) отмечено у видов, обитающих в луговом сообществе (Primula farinosa, Р,nutans, Ranunculus propinquus, R.rigescens). Среди лесных видов максимальная оводненность листьев отмечена у Pyrola asarifolia - 81,04 в 2002 г. Среди степных видов максимальная оводненность листьев отмечена у Leibnitzia anandria - 78,99%. Самая низкая оводненность листьев (не превышает 50%) отмечена у двух степных видов - Alyssum ■ obovatum, Pulsatilla turczaninovii и лесного вида -Vaccinium vitis-idaea.
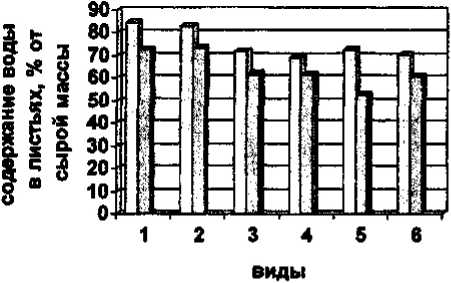
□ абсолютный максимум
0 абсолютный минимум
Рис.1. Содержание воды в листьях раннецветущих растений (средние данные за май 2001 - 2004 гг.) 1 - Primula farinosa; 2 - Р. nutans; 3 - Pyrola asarifolia; 4 - Pulsatilla multifida; 5 - Leibnitzia anandria;
6 - Thlaspi cochleariforme
По количеству воды в листьях в различные фенологические фазы выявлено, что наибольшая оводненность листьев у луговых видов отмечена во время вегетации до цветения, бутонизации. К началу цветения запас влаги в листьях неуклонно снижается. Для видов, обитающих в лесном сообществе, эта закономерность не наблюдается. Максимум овод-ненности листьев во время вегетации до цветения отмечен только у Potentilla fragarioides. У четырех видов (Pyrola asarifolia, Vaccinium vitis-idaea, Oxytropis myriophylla, Fragaria orientalis) максимум оводненности листьев отмечен во время цветения. Для исследованных нами 11 видов, обитающих в степном сообществе, выявлена следующая закономерность (рис. 2). Практически у всех изученных видов данного сообщества максимум оводненности листьев отмечен в фазу вегетации до цветения и бутонизации. К началу цветения оводненность тканей уменьшается и только у двух видов (Gagea pauciflora, Androsace incana) оводненность тканей выше в фазу цветения.
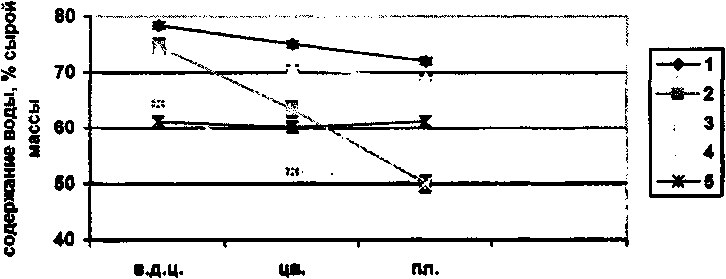
Рис.2. Содержание воды в надземной фитомассе степных растений в различные фенофазы (средние данные за май 2002 - 2004 гг.)
1 - Thlaspi cochleariforme; 2 - Arctogeron gramineum; 3 - Leibnitzia anandria; 4 - Potentilla leucophylla; 5 - Pulsatilla turczanlnovii
Важное значение имеют данные по диапазону влажности, поскольку они характеризуют степень подвижности водного режима, а следовательно, и способность растений адаптироваться к самым разнообразным условиям среды. Наши исследования показали, что изученные раннецветущие растения характери зуются достаточно лабильным водным режимом. При изучении фракционного состава воды в листьях и корнях растений было обнаружено, что у всех исследованных видов преобладает фракция связанной воды (табл.2), которая составляет более половины общего запаса влаги.
Таблица 2
Группы видов по количеству общей, связанной и свободной воды в надземной фнтомассе (средние данные за май 2001-2004гг), % сырой массы
|
Группы |
Виды |
Общая |
Связанная |
Свободная |
|
1 |
primulafarmosa |
83,90*1,23 |
64,86*1,72 |
19,04*1,13 |
|
Primula nutans |
82,59*0.82 |
62,43*1,19 |
20,16*1,25 |
|
|
Ranunculus propinquus |
79,74*0,41 |
60,32*1,85 |
19,42*1,76 |
|
|
Ranunculus rigescens |
76,73*1,02 |
56,46*2,35 |
20,27*1,86 |
|
|
Thlaspi cochlearijorme |
74,86*0,61 |
62,10*1,44 |
12,76*1,04 |
|
|
Pulsatilla multifida |
74,51*0,28 |
61,89*1,52 |
12,63*1,49 |
|
|
Ciminalis aquatica |
74,38*0,73 |
63,38*0,82 |
11,00*0,73 |
|
|
Leibnitzia anandria |
73.15*0,53 |
58,27*0,56 |
14,88*0,97 |
|
|
II |
Euphorbia karoi |
70,73*2,90 |
54,49*1,19 |
16,19*1,45 |
|
Stellaria cherleriae |
70,68*0,33 |
58,39*1,34 |
12,29*1,11 |
|
|
Gagea pauciflora |
70.60*0,18 |
55.18*1,82 |
15.42*1,99 |
|
|
111 |
Potentilla fragarioides-луг |
69,76*1,15 |
47,31*2,00 |
22,45*1,58 |
|
Pyrola asarifolia |
66,17*0,49 |
42,32*1,07 |
23,85*1,10 |
|
|
Potentilla fragarioides-jtec |
65,19*1,39 |
50,43*2,25 |
14,76*1,11 |
|
|
Oxytropis myriophylla |
64,01*1,15 |
55,15*1,32 |
8,86*0,59 |
|
|
Arctogeron gramineum |
63,00*2,50 |
50,47*1,60 |
12,53*1,64 |
|
|
Eremogone capiliaris |
62,63*0,71 |
46,43*1,22 |
16,20*1,56 |
|
|
Androsace incana |
61,77*0,66 |
52,86*0,68 |
8,91*0.46 |
|
|
Pulsatilla turczaninovii |
61,13*1,00 |
35,54*0,69 |
25,59*1,69 |
|
|
Fragaria orientalis |
60,97*1,02 |
44,87*1,17 |
16.10*0,84 |
|
|
Potentilla leucophylla |
60,38*0,09 |
48,80*1,83 |
11,58*1,74 |
|
|
IV |
Alyssum obavatum |
59,75*0,63 |
53,15*0,95 |
6,59*0,59 |
|
Vaccinium vitis-idaea |
46,92*0,53 |
37,44*1,08 |
9,48*0,90 |
Известно, что содержание свободной или активной воды определяет интенсивность течения физиологических процессов (транспирации, ростовых процессов и т.п.), а содержание связанной или трудноотнимаемой воды - устойчивость растений к неблагоприятным условиям среды (к засухе, пониженным или повышенным температурам и т.п.), большое содержание связанной воды в тканях исследованных видов свидетельствует о том, что раннецветущие растения хорошо приспособлены к недостаточному увлажнению и резким колебаниям содержания воды в почве.
Таким образом, наблюдения за ростом раннецветущих растений показали, что и вынужденный зимний покой, и особенности накопления запасных питательных веществ в тканях, и преобладание фракции связанной воды в листьях и корнях способствует раннему развитию растений в весенний период.
Список литературы Эколого-биологические особенности раннецветущих растений Восточного Забайкалья
- Бейдеман И.Н. Изучение фенологии растений/И.Н. Бейдеман//Полевая геоботаника. -М.-Л.: Изд-во АН СССР. Т. 12. -1960. -С. 333 -366.
- Генкель П.А. Физиология жаро-и засухоустойчивости растений/П. А. Генкель -М: Наука, 1982. -280 с.
- Генкель П.А.Состояние покоя и морозоустойчивость плодовых растений/П.А. Генкель, Е.З. Окнина М.: Наука, 1964.-242с.
- Горшкова А.А. Биология степных пастбищных растений Забайкалья/А.А. Горшкова -М.: Наука, 1966. -272 с.
- Горшкова А.А. Экология водного режима степных растений Забайкалья/А.А. Горшкова -Иркутск: Изд-во АН СССР, 1971.-215с.
- Горышина Т.К. О характере осенне-зимнего покоя у различных сезонных групп травянистых растений дубравы/Т.К. Горышина//Бюллетень МОИП. f.XLH(5).-1965.-C.46-52.
- Копытова Л.Д. Основные показатели водного режима степных растений в связи с их экологией и биоморфологией/Л.Д. Копытова//Бот. журн. -Т.88. -№ 5.-2003.-С. 111-119.
- Сулейманов И.Г. Состояние и роль воды в растении/И.Г. Сулейманов: Казань: Изд-во Казанского унта, 1972.-С. 3-38.
- Сулейманов И.Г. Структурно-физические свойства протоплазмы и ее компонентов в связи с проблемой морозоустойчивости культурных растений/И.Г. Сулейманов. -Казань. 1964.-С. 139-165.
- Федоскин Н.В. Жизненные формы травянистых растений степей юго-восточного Забайкалья/Н.В. Федоскин//Бюллетень МОИП. -Отд. биологии. Т. LXXIX (6).-1974.-С. 98-108.
- Федоскин Н.В. Жизненные формы растений степей юго-восточного Забайкалья/Н.В. Федоскин//Флора, раститагыюсть и растительные ресурсы Забайкалья и сопредельных областей. -Чита. -Вып. 5. -1975. -С. 31 -34.
- Федоскин Н.В. Побегообразование растений степей юго-восточного Забайкалья/Н.В. Федоскин//Флора, растительность и растительные ресурсы Забайкалья и сопредельных областей. -Чита -Вып. 5. -1975. -С.52-53.
- Федоскин Н.В. Характер зимнего и летнего периодов покоя у некоторых травянистых растений/Н.В. Федоскин//Природные ресурсы западной области РСФСР и их рациональное использование -Смоленск. 1974.-С.134-139.
- Фурст Г.Г. Методы анатомо-гистохимического исследования растительных тканей/Г.Г. Фурст. -М.: Наука, 1979.-128 с.
- Щербаков И.П. Об изучении зимнего покоя у древесных растений/И.П. Щербаков//Известия Сиб. отделения АН СССР. -№4. -1960. -С. 87-96.
