Эколого-демографический анализ ценопопуляций спорофитов листовика обыкновенного Phyllitis scolopendrium (L.) Newm. (Aspleniaceae) в Западном Закавказье
 Newm. (Aspleniaceae) в Западном Закавказье")
Автор: Державина Нина Михайловна, Ковынева Светлана Владимировна, Силаева Жанна Геннадьевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 2, 2016 года.
Бесплатный доступ
Изучены демографическая структура и пространственная организация 6 ценопопуляций (ЦП) спорофитов листовика обыкновенного Phyllitis scolopendrium (L.) Newm. в Западном Закавказье. Показано, что вид образует устойчивые дефинитивные ЦП, самоподдерживающиеся только споровым путем, требующие специфического экологического пространства. Устойчивое сосуществование с конкурентами возможно благодаря фитопатиентности, а экопатиентность обеспечивает оптимальное использование скудных условий экотопа.
Гемистенобионт, виталитет, экологическая позиция, возрастной спектр
Короткий адрес: https://sciup.org/146116646
IDR: 146116646 | УДК: 574.23:581.52
Ecologo-demographic analysis of coenopopulations of sporophyte Phyllitis scolopendrium (L.) Newm. (Aspleniaceae) in the Western Transcaucasia
The demographic structure and spatial organization of the six coenopopulations (CP) of sporophytes Phyllitis scolopendrium (L.) Newm. in the Western Transcaucasia were studied. The species forms stable definitive CPs, maintained by spore reproduction and requiring specific ecological space. Sustainable coexistence with competitors is possible due to phytopatientness; the ecopatientness ensures optimal use of scarce conditions of the ecotope.
Текст научной статьи Эколого-демографический анализ ценопопуляций спорофитов листовика обыкновенного Phyllitis scolopendrium (L.) Newm. (Aspleniaceae) в Западном Закавказье
Введение. В наши дни все чаще приходится сталкиваться с фактами угрожающего нарушения экологического равновесия биологических систем. В связи с этим актуальны не только разработки и реализация эффективных мероприятий по сохранению мирового растительного разнообразия, а также прогнозирование поведения популяций растений. В последние десятилетия появляются работы, посвященные изучению онтоморфогенеза, структуры ценопопуляций (ЦП) различных групп высших растений, позволяющие понять закономерности адаптации растений к абиотической среде и разработать эффективные мероприятия по охране, прежде всего видов, находящихся под угрозой исчезновения. Эти исследования посвящены в основном анализу ЦП цветковых растений, в то время как работы, связанные с выявлением многочисленных взаимосвязей между популяциями спорофитов папоротников и абиотическими и биотическими факторами их местообитаний, единичны (Лащинский, Шорина, 1985; Серая и др., 1985; Науялис, 1989; Шорина, Державина, 1992; Шорина, 1994; Гуреева, 1987, 2001; Криницын, 2004; Силаева, 2009).
Объект исследования - папоротник Phyllitis scolopendrium (L.) Newm. - Листовик обыкновенный. Это - короткокорневищный, радиальносимметричный, вертикально- или косорозеточный, -62- моноцентрический, многолетний, вечнозеленый гемикриптофит, слабо ветвящийся, неэффективно вегетативно размножающийся (Державина, 2006). Р. scolopendrium - декоративное и лекарственное растение (Дикорастущие..., 2001).
Целью работы стало эколого-демографическое исследование ЦП спорофитов Р. scolopendrium в Западном Закавказье и попытка прогнозирования их поведения в пессимальных условиях среды.
Материал и методы. Шесть ЦП спорофитов Р. scolopendrium изучены в горно-лесном поясе Лазаревского, Туапсинского и Хостинского р-нов Большого Сочи и в окрестностях г. Сочи Краснодарского края в июле 2012 г. Ниже приводим их краткую характеристику.
ЦП 1. Якорная щель. Лазаревский р-он г. Сочи, окр. пос. Якорная щель. Левый борт р. Хаджиек. Возле каскада 1,5-2 м водопадов. Борта высотой 10-12 м. Каштаново-буковый лес на склоне 80-85°. Сомкнутость крон 60-80 %. Характер субстрата: карбонатный флиш. Моховое покрытие 100 %. Проективное покрытие видом (ПП) - 5 %.
ЦП 2. 33 водопада. Лазаревский р-н г. Сочи, горное ущелье в долине р. Шахе, в 4-х км от аула Большой Кичмай, борта ручья Джегош высотой 2-3 м. Грабово-самшитовый лес. Сомкнутость крон - 80-85%. Характер субстрата: карбонатный флиш. Общее проективное покрытие травостоем (ОПП) -45-50 %, ПП -10 %.
ЦП 3. Тисо-самшитовая роща. Микрорайон пос. Хоста на восточном склоне горы Ахун, правый берег р. Хоста, скальный лабиринт, Ю-3 экспозиция, 120 м над ур. моря. Самшитовый лес с единичными грабинником, липой и ясенем. Сомкнутость крон - 95 %. Верхнемеловые известняки, поросшие мхом. Высота от 0,3-0,5 м до 1 м. Моховое покрытие около 80 %, ПП - 3 %.
ЦП 4. Зубова щель. Лазаревский р-он г. Сочи. Близ пос. Чемитоквадже. Каньон Зубова щель. Левый берег р. Чемит (Чемитоквадже). Уступ высотой 1 м из отдельных каменных глыб. Ясенево-грабово-кленовый лес. Сомкнутость крон - 95 %. Характер субстрата: карбонатный флиш. ОПП - 60 %. Моховое покрытие 40%, ПП -20%.
ЦП 5. Каньон Прохладный, г. Сочи, Сочинский нац. парк, борта ручья Прохладного - притока р. Чужая. Высота бортов - 3,5 - 7-10 м. На бортах: вяз, ольха, клен, бук, самшит. Деревья самшита полностью покрыты мхом. Сомкнутость крон над правым бортом - 60-70 %, над левым - 30-40 %. Характер субстрата: слоистый известняк. Моховой покров - 20-30 % на левом берегу, на правом - 90 %. ПП -10 %.
ЦП 6. Каткова щель. Лазаревский р-он г. Сочи, окр. пос. Каткова щель. Ущелье Каткова щель. Правый борт р. Чухукт, высотой до 10-15 м. Кленово-грабовый лес. Сомкнутость крон - 75-80 %. Характер субстрата:
слоистый известняк. ОПП - 50-60 %. Камни замшелые на 50%. ПП-5%.
Экологическая оценка местообитаний папоротника проведена на основе геоботанических описаний по традиционным методикам. Флористические списки обработаны по индикаторным шкалам Д.Н. Цыганова (1983) в компьютерной программе EcoScaleWin (Грохлина, Ханина, 2006). Рассчитаны: реализованная (REV) экологическая валентность, которая сопоставлена с потенциальной (PEV) по шкалам Д.Н. Цыганова (1983), а также коэффициенты экологической эффективности (К ес. Eff), индексы толерантности по климатическим (It клим.) и эдафическим факторам (It почв.) и обобщенный индекс толерантности (Ito) (Жукова, 2004).
Проанализировано 75 пробных площадок размером 1 м2. Исследованы основные демографические параметры ЦП: плотность (М), онтогенетический (возрастной) спектр, жизненность и способ самоподдержания. При этом использованы различные демографические показатели: индекс возрастности (А) (Уранов, 1975), индекс эффективности (со) (Животовский, 2001), индекс восстановления (1В), индекс замещения (13) (Жукова, 1995). Тип ЦП определен по классификациям А.А. Уранова и О.В. Смирновой (1969) и «дельта-омега» (Животовский, 2001). Жизненность (виталитет) ЦП оценена по методике Ю.А. Злобина (1989).
Результаты и их обсуждение. Проведенные нами наблюдения показали, что в местах исследования папоротник обитает как в тенистых, так и в осветленных кленово-ясенево-грабовых, каштаново-буковых, грабово-самшитовых, самшитовых и др. лесах. Под пологом леса, как известно, особенности влаго- и теплообеспеченности растений значительно смягчаются. Здесь складываются и благоприятные условия почвенного питания, т.к. к отмирающим частям самого папоротника добавляется опад растений разных ярусов сообщества.
Спорофиты Р. scolopendrium населяют скопления гумуса в трещинах и расселинах известняковых и доломитовых тенистых скал (хазмофит), ниши под глыбами горных пород, мшистые камни на склонах ущелий, их днищах или водораздельных гривах, осыпи, проникает глубоко в пещеры. В то же время они способны расти и на субстратах с низким содержанием кальция, и в условиях общей нехватки питательных веществ.
На дне углублений между камнями особи Р. scolopendrium выглядят несколько лучше (имеют большее число ваий), чем на бортах каменных развалов. Зато особи, обитающие под пологом леса в небольших накоплениях перегноя, но в атмосфере повышенной влажности (например, на бортах водопадов в Лазаревском р-не), имеют даже более крупные вайи, чем живущие в глубине леса на богатых почвах. Вероятно, гидратура почвы и воздуха - более важный фактор в жизни этого вида, чем качество субстрата.
Реже вид встречается на почве и стволах как поваленных (ЦП 2), так и живых деревьев на небольшой высоте. Г.М. Кадыров и А.М. Аскеров (1972) приводят этот папоротник в Талыше в качестве эпифита, часто встречающегося на стволах Alnus subcordata С.А. Mey., изредка -на стволах Parrotia persica С.А. Mey., Acer velutinum Boiss., Pterocarya pterocarpa (Michx.) Knuth., Zelkova carpinifolia (Pall.) Dipp.
Благоприятные условия для обитания Р. scolopendrium складываются за счет высокой влажности воздуха и почвы, которые обеспечивают близость горных рек, ручьев и водопадов и, соответственно, морской субтропический климат. Еще одна характерная черта гигротермического режима местообитаний Р. scolopendrium - круглогодичное достаточное увлажнение за счет обилия стекающих со склонов в ущелья и долины атмосферных и грунтовых вод. Сезонное увлажнение обеспечивают таяние снегов весной, затяжные дожди и интенсивные ливни в начале осени. Для горных районов весьма характерны сильные туманы.
Спорофиты и гаметофиты этого вида отличаются разной стратегией в ответ на малую интенсивность света. Споры Р. scolopendrium, как известно, могут прорастать в полной темноте (Gams, 1938). Неудивительно, что гаметофиты приспособлены к существованию с низкой точкой компенсации (Gams, 1938; Porter, 1994) и, вероятно, являются облигатными сциофитами. У спорофитов же более динамичный фотосинтетический аппарат и более широкий диапазон толерантности по отношению к свету. По положению точки компенсации и точки насыщения на кривой фотосинтеза спорофиты вида больше напоминают листья теневыносливых растений (Porter, 1994). Особи, встреченные нами на поверхности известняковых скал на открытом свету, были с деформированными ваиями желтоватого цвета. Подобные изменения ваий отмечает и J. Porter (1994). Будучи вечнозеленым папоротником Р. scolopendrium способен вегетировать в листопадных лесах до выпадения снега, компенсируя тем самым нехватку света во время летнего затенения.
Анализ амплитуд факторов по шкалам Д.Н. Цыганова (1983) показал (табл. 1), что Р. scolopendrium выступает в качестве стеновалента по криоклиматическому фактору и фактору увлажнения почвы. По термоклиматическому фактору обеспеченности почв азотом и их кислотности - занимает позиции гемистеновалента. По шкалам континентальности и богатства почв он гемиэвривалентен. По омброклиматическому фактору папоротник мезовалентен и лишь по шкале освещенности эвривалентен, охватывая достаточно широкий диапазон условий.
Результаты обработки геоботанических описаний местообитаний ЦП вида с помощью программного комплекса EcoScaleWin, показали что исследованные ЦП приурочены к достаточно узкому диапазону как климатических так и почвенных условий (рис. 1). Об этом свидетельствуют коэффициенты экологической эффективности. Наибольшее экологическое пространство освоено видом по фактору кислотности и увлажнения почвы, что объясняется спецификой занимаемых им местообитаний. Обобщенный индекс толерантности (Ito=O,45) позволяют заключить, что этот вид гемистенобионтен (ГСБ) по отношению к комплексу факторов (табл.1).
Для того, что бы вид мог реализовать основные потенциальные возможности в местах нашего исследования, необходимо совпадение комплекса факторов. Гаметофиты должны дать начало спорофитам на каменистом субстрате с нейтральной или щелочной реакцией, либо на бурых оподзоленных почвах с подстилающими их известняками, либо на перегнойно-карбонатных почвах, почвообразовательный процесс в которых протекает в условиях избытка растворов кальция. Важна высота над уровнем моря: оптимальна нижняя часть горно-лесного пояса. При этом мозаичные и разнообразные скалистые участки должны быть ранее заселены пионерными растениями, среди которых преобладают мхи, поскольку их синузии хорошо удерживают влагу. При высокой атмосферной влажности и нормально увлажненном элементе мезорельефа еще важнее в качестве косвенного фактора - микрорельеф: в виде нарушенных участков почвы, трещин, ниш, карнизов в каменных глыбах и между ними и т.п., в которых может накапливаться гумус. Кроме этого, важно учитывать, что местообитания, доступные этому виду по его экологическим возможностям, могут быть уже заняты конкурентами, что, несомненно, сужает или сдвигает его экологический оптимум. В этом случае проявляет себя правило конкурентного исключения.
Пространственная структура изученных ЦП объясняется двойственностью популяционной жизни папоротников, связанной с чередованием двух гетероморфных поколений - спорофитов и гаметофитов, находящихся на разных уровнях морфологической организации и, соответственно, имеющих разные экологические потребности.
У Р. scolopendrium поселения спорофитов и гаметофитов могут быть, как изолированы экологически, например, спорофиты обитать на гумусе в расселинах скал на склоне, а гаметофиты - у подножия склона в микроуглублениях обнаженной почвы, так и сосуществовать в одной нише ценоза: и те и другие - в трещинах скальной породы, либо под каменным карнизом в моховом покрове. Гаметофиты имеют тенденцию расти плотными скоплениями разной численности, обеспечивающими
Вестник ТвГУ. Серия "Биология и экология". 2016. № 2 межгаметофитные взаимодействия, включая ответы, индуцированные гормоном антеридиогеном (отвечающим за образование антеридиев, ингибирование вегетативного роста и прорастанию спор в темноте). Спорофиты же отличает контагиозное размещение по площади ценоза.
Тип размещения сначала гаметофитов, а затем и спорофитов создается под действием ряда факторов: 1) наличия недолговечных микросайтов на эродированной лесной почве, моховых подушках, осыпях, голых камнях, под каменными карнизами, на обочинах дорог и т.п.; 2) плотности и дальности разноса спор; 3) смыва с крутых склонов во время дождя, как части спор, так и самих гаметофитов и даже молодых спорофитов; 4) возможности части спор быть съеденной почвенными животными.
Таблица 1
Потенциальные, реализованные экологические валентности (PEV, REV), коэффициенты экологической эффективности (K.ec.eff.)
и индексы толерантности (It) по климатическим и эдафическим факторам Phyllitis scolopendriwn по шкалам Д.Н. Цыганова (1983)
|
Фактор |
PEV |
REV |
K.ec.eff. (%) |
It |
|
Тт |
0,4 ГСВ |
0,05 |
12,5 |
0,45 ГСБ |
|
Кп |
0,6 ГЭВ |
0,067 |
И,2 |
|
|
От |
0,47 МВ |
0,042 |
8,9 |
|
|
Сг |
0,33 св |
0,054 |
16,36 |
|
|
Hd |
0,22 СВ |
0,04 |
18,2 |
0,36 ГСБ |
|
Тг |
0,37 ГЭВ |
0,026 |
7 |
|
|
Nt |
0,45 ГСВ |
0,06 |
13,3 |
|
|
Rc |
0,39 ГСВ |
0,08 |
20,5 |
|
|
Ес |
0,78 ЭВ |
0,12 |
15,38 |
- |
Примечание. Факторы: Тт - термоклиматический; Кп - континентальности климата; От - омброклиматический; Ст - криоклиматический; Hd - увлажнения почвы; Nt -обеспеченности почвы азотом; Тг - богатства почвы; Rc - кислотности почвы; Ес -освещенности-затенения. ГСВ - гемистеновалент, ГЭВ - гемиэвривалент, МВ -мезовалент, СВ - стеновалент
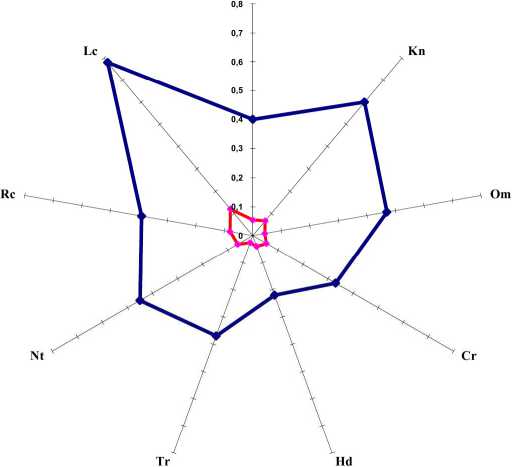
-
• PEV - потенциальная экологическая валентность
-
• REV -реализованная экологическая валентность
Рис. 1. Потенциальная и реализованная экологические валентности ЦП Phyllitis scolopendrium
Примечание. Буквенные обозначения приведены в табл. 1
Это обеспечивает постоянные волны возобновления и отмирания, омоложения и старения поселений папоротника. Свидетельством этого служит сочетание разновозрастных, разноразмерных локусов в пределах каждой изученной ЦП, а так как локусы развиваются асинхронно во времени и пространстве, каждая ЦП претерпевает флуктуационные изменения. Учитывая сказанное, тип динамики изученных ЦП можно определить как волнообразно-флуктуационный.
Поскольку в качестве основного демографического параметра ЦП служит ее онтогенетический спектр, приводим рисунок онтогенетических состояний спорофита (рис. 2).
На фоне достаточно сходных абиотических параметров местообитаний Р. scolopendrium все же формируются разноликие, и в то же время во многом подобные ЦП. Анализируя их демографические параметры (табл. 2), можно заключить, что все они - нормальные, дефинитивные, т.к. способны к возобновлению и находятся в относительном равновесии с экотоном. Поскольку в большинстве ЦП преобладают репродуктивные растения, то их можно охарактеризовать как зрелые, за исключением ЦП 1, которая является молодой.
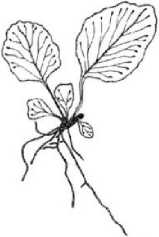
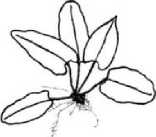
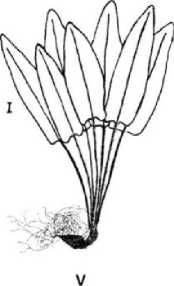
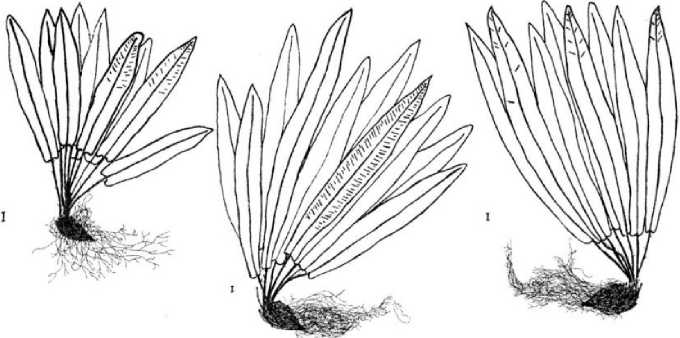
Spl Sp2 Sp3

Рис. 2. Онтогенетические состояния спорофита Phyllitis scolopendrium. Масштаб I - 0,01 м
Анализ онтогенетической структуры ценопопуляций Р. scolopendrium позволяет выделить следующие типы онтогенетических спектров (рис. 3):
-
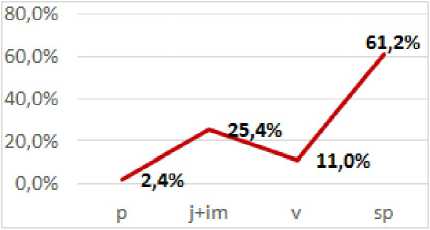
1) двувершинный с пиками на прегенеративной и генеративной группах особей (ЦП 2, 5). Обе ЦП обитают в сходных экологических условиях: на бортах лесных ручьев. Максимум спектра в обеих ЦП приходится на группу спороносящих, а также ювенильных и имматурных растений, обеспечивающих возобновление спорофитов. Это свидетельствует о зрелости ЦП и относительно высокой плотности
особей. Специфика спектра объясняется значительной продолжительностью онтогенетических состояний и длительностью жизни спороносящих растений, а также постоянной инвазией спор. Отсутствие гаметофитов в ЦП 2 связано с тем, что, сформировавшись весной, они успели оплодотвориться и дать начало ювенильным и имматурным спорофитам, положив начало волне возобновления ЦП.
В ЦП 5 репродуктивные спорофиты растут либо по краям водотока, на каменистом замшелом грунте почти спускаясь в воду, либо на моховом покрове крупных валунов чуть выше. При этом во время осенних дождей и весенних паводков споры и молодые растения легко смываются и уносятся потоками воды. Этим объясняется отсутствие в ней гаметофитов и пререпродуктивных растений (проростков, ювенильных и имматурных). По классификации «дельта-омега» ЦП 2 является зрелой, а ЦП 5 - зреющей.
-
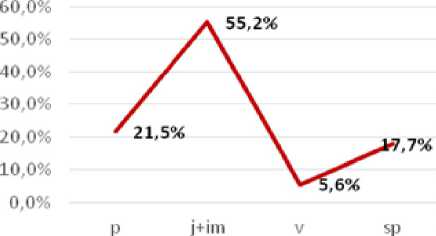
2) левосторонний, с преобладанием пререпродуктивных особей (ЦП 1). Механизм формирования такого спектра связан со споровым способом самоподдержания и накоплением молодых особей. Абсолютный максимум в спектре приходится на группу ювенильных и имматурных растений.
Успешному развитию гаметофитов под пологом леса, процессу оплодотворения и формированию спорофитов благоприятствуют моховые синузии, которые обеспечивают удержание воды в сухой сезон. Неудивительно, что именно в этой ЦП сложилась максимальная экологическая плотность особей - 15,3 шт/м2. По классификации «дельтаомега» ЦП 1 является молодой.
-
3) одновершинный, центрированный, с пиком на репродуктивной фракции особей - средневозрастных спороносящих растениях (ЦП 3, 4, 6). В этих ЦП обнаружены многорозеточные субсенильные спорофиты с 6-8 розетками ваий.
В ЦП 3 в тисо-самшитовой роще затруднено споровое возобновление. Несмотря на то, что проективное покрытие мха составляет около 80%, это местообитание отличает значительная сухость воздуха в связи с удаленностью от реки. Плотность ЦП довольно низкая за счет отсутствия молодых скоплений. Гаметофиты в этой ЦП не обнаружены, зато несколько скоплений до полутора сотен особей мы нашли на обнаженной почве вдоль водотока и на голых влажных глыбах известняка на высоком борте лесного ручья в этом же фитоценозе.
ЦП 4 отличает относительно высокая экологическая плотность особей. Отсутствие фракции гаметофитов и проростков связано с тем, что к июлю уже успевают сформироваться локусы возобновления. Проективное покрытие средневозрастных спорофитов достигает 20 %.
Якорная щель ЦП 1
Тнсо-сампштовая роща ЦП 3
33 водопада ЦП 2
Зубова щель ЦП 4

Каньон прохладный ЦП 5
Каткова Щель ЦП 6
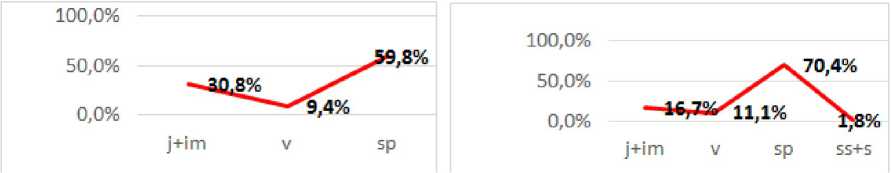
Рис. 3 . Онтогенетические спектры Phyllitis scolopendrium.
р - проростки, j+im - ювенильные и имматурные растения, у - виргинильные растения, sp - спороносящие растения, 55+5 - субсенильные и сенильные растения
В ЦП 6 отмечена минимальная экологическая плотность особей, связанная с небольшим процентом пререпродуктивных особей. Это объясняется высоким проективным покрытием травостоя (50-60 %) и большой крутизной склона. При этом, если спороносящие особи, развившие мощные корневые системы, удерживаются в смываемом во время сезонных дождей субстрате на крутом склоне, то представители локусов возобновления легко уносятся потоками воды.
По классификации «дельта-омега» ЦП 3, ЦП 4, ЦП 6 относятся к стареющим.
Индексы восстановления (1в) и замещения (1з) достаточно высокие для большинства ЦП (табл. 2), что указывает на благоприятные условия для их самоподдержания. В самоподдержапии ЦП Р. scolopendrium, как моноцентрического растения, главная роль принадлежит споровому возобновлению, которое активно почти во всех местообитаниях. Удержание и расширение площади ценопопуляционных локусов, а также расселение вида в другие местообитания происходит исключительно споровым путем.
Таблица 2 Основные демографические показатели ЦП Phyllitis scolopendrium
|
№ ЦП |
М шт/м2 |
А |
со |
1в |
ц |
Тип популяции по классификации «дельта-омега» |
Тип популяции, полночленность |
Виталитетный спектр / Тип популяции по жизненному состоянию |
|
1 |
15,3 |
0,13 |
0,34 |
3,44 |
- |
молодая |
Нормальная, дефинитивная, молодая, не полночленная |
43,3%:30%:26,7% Процветающая Q=11 |
|
2 |
13,9 |
0,36 |
0,72 |
0,97 |
- |
зрелая |
Нормальная, дефинитивная, зрелая, не полночленная |
43,3%:26,7%:30% Процветающая Q=10,5 |
|
3 |
5,73 |
0,83 |
0,97 |
0,18 |
0,15 |
стареющая |
Нормальная, дефинитивная, стареющая, не полночленная |
36,7%:30%:33,3% Равновесная Q=10 |
|
4 |
10,73 |
0,83 |
0,85 |
0,6 |
0,47 |
стареющая |
Нормальная, дефинитивная, зрелая, полночленная |
36,7%:30%:33,3% Равновесная Q=10 |
|
5 |
11,7 |
0,34 |
0,71 |
0,96 |
- |
зреющая |
Нормальная, дефинитивная, зрелая, не полночленная |
33,3%:36,7%:30% Процветающая Q=10,5 |
|
6 |
5,4 |
0,79 |
0,8 |
1,38 |
1,09 |
стареющая |
Нормальная, дефинитивная, зрелая, полночленная |
50%:23,3%:26,7% Процветающая Q=11 |
При оценке виталитетной структуры изученных популяций выявлено доминирование спорофитов с высоким (класс «а») (ЦП 1, ЦП 2, ЦП 3, ЦП 4, ЦП 6) и средним (класс «Ь») уровнями жизненности (ЦП 5), что указывает на благоприятные экологические и фитоценотические условия местообитаний вида (Табл. 2). При этом уровень жизненного состояния повышается у особей, обитающих вблизи водоемов. Присутствие особей с низким уровнем жизненности (класса «с») способствует формированию резерва, обеспечивающего устойчивость ЦП. Виталитетные спектры ЦП папоротника характеризуется левосторонней асимметрией и, учитывая Q-критерий (Табл. 2), позволяющий оценить жизненность популяции в целом, изученные ЦП можно назвать процветающими (ЦП 1, ЦП 2, ЦП 5, ЦП 6) и равновесными (ЦП 3, ЦП 4).
Заключение. Учет специфики местообитаний Р. scolopendrium, его структурных характеристик и некоторых особенностей физиологии позволяют заключить, что этот папоротник физиологически и структурно приспособлен к жизни в затенении и в условиях повышенной гидратуры, т. е. требует специфического качества экологического пространства. Однако он демонстрирует высокую адаптируемость при обитании в открытых местах, где погружается в трещины между камнями. При этом он развивает меньших размеров более плотные и жесткие пластинки ваий с ббльшим числом устьиц по сравнению с затененными особями.
В отношении богатства почв у этого папоротника обнаружен широкий экологический диапазон. С одной стороны, предпочтительными местообитаниями для Р. scolopendrium являются ниши между россыпями камней со значительными накоплениями гумуса под пологом леса, с другой стороны, вид может заселять практически обнаженные скальные склоны, используя мельчайшие трещины в породе.
В исследованных ценозах складывается диапазон условий и ресурсов, позволяющих виду занимать второстепенное положение и образовывать нормальные, дефинитивные ЦП, существующие на протяжении длительного времени благодаря высокой энергии спорового размножения. Устойчивое сосуществование с конкурентами возможно благодаря фитопатиентности, а экопатиентность обеспечивает оптимальное использование скудных условий экотона.
Сравнительно узкое экологическое пространство, занимаемое видом, позволяет предполагать его уязвимость в случае наступления пессимальных условий: вырубки леса, пересыхания близ расположенных водоемов и др., что повлечет за собой общее неблагоприятное изменение условий существования.
ECOLOGO-DEMOGRAPHIC ANALYSIS
OF COENOPOPULATIONS OF SPOROPHYTE
PHYLLITIS SCOLOPENDRIUM (L.) NEWM. (ASPLENIACEAE) IN THE WESTERN TRANSCAUCASIA
N.M. Derzhavina1, S.V. Kovineva1, Z.G. Silaeva2
'Turgenev Orel State University, Orel
2Orel State Agrarian University, Orel
The demographic structure and spatial organization of the six coenopopulations (CP) of sporophytes Phyllitis scolopendrium (L.) Newm. in the Western Transcaucasia were studied. The species forms stable definitive CPs, maintained by spore reproduction and requiring specific ecological space. Sustainable coexistence with competitors is possible due to phytopatientness; the ecopatientness ensures optimal use of scarce conditions of the ecotope.
Державина Н.М. Эколого-демографический анализ популяций спорофитов листовика обыкновенного Phyllitis scolopendrum (L.) Newm. (Aspleniaceae) в Западном Закавказье / Н.М. Державина, С.В. Ковынева, Ж.Г. Силаева // Вести. ТвГУ. Сер.: Биология и экология. 2016. №2. С. 62-75.
Список литературы Эколого-демографический анализ ценопопуляций спорофитов листовика обыкновенного Phyllitis scolopendrium (L.) Newm. (Aspleniaceae) в Западном Закавказье
- Грохлина Т.И., Ханина Л. 2006. Автоматизация обработки геоботанических описаний по экологическим шкалам. Принципы и способы сохранения биоразнообразия//Материалы II Всероссийской научной конференции. Йошкар-Ола: Мар. гос. ун-т. C. 87-89.
- Гуреева И.И. 1987. Структура ценопопуляций крупнокорневищных папоротников//Популяционная экология растений. М.: Наука. С. 125-128.
- Гуреева И.И. 2001. Равноспоровые папоротники Южной Сибири (систематика, происхождение, биоморфология, популяционная биология). Томск. 158 с.
- Державина Н.М. 2006. Биоморфология и анатомия равноспоровых папоротников (эпилитов, эпифитов, земноводных и водных) в связи с адаптациогенезом: автореф. дис.. д-ра биол. наук. М. 50 с.
- Дикорастущие полезные растения России. 2001/под ред. А.Л. Буданцева и Е.Е. Лесиовской. СПб: Изд-во СП ХВА. 663 c.
- Животовский А.А. 2001. Онтогенетические состояния, эффективная плотность и классификация растений//Экология. № 1. С. 3-7.
- Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола: РИИК «Ланар». 224 с.
- Жукова Л.А. 2004. Оценка экологической валентности видов основных эколого-ценотических групп//Восточно-европейские леса: история в голоцене и современность. М.: «Наука». Кн. 1, раздел 3,5. С. 256-270.
- Злобин Ю.А. 1989. Принципы и методы изучения ценотических популяций растений. Казань. 146 с.
- Кадыров Г.М., Аскеров А.М. 1972. Эпифитные папоротники Талыша//Уч. Зап. Азерб. ун-та. Сер. Биол. науки. №1. С. 8-10.
- Криницын И.Г. 2004. Онтогенез и структура популяций спорофитов некоторых видов рода Botrychium Sw. в подзонах южной тайги и подтайги Европейской России: автореф. дис. … канд. биол. наук. Барнаул. 19 с.
- Лащинский Н.Н., Шорина Н.И. 1985. Онтогенез спорофита и структура ценопопуляций Polystichum braunii (Spenn.) Fee в черневой тайге Салаирского кряжа//Изв. СО АН СССР. Сер. биол. наук. Вып. 2. № 13. С. 35-44.
- Науялис И.И. 1989. Организация популяций гаметофитов папоротников//Бот. журн. Т. 74. № 7. С. 994-1003.
- Серая Г.П. и др. 1985. Сравнительный анализ структуры ценопопуляций крупнокорневищных папоротников Висимского заповедника//Структура и динамика биогеоценозов Урала. Свердловск. С. 49-57.
- Силаева Ж.Г. 2009. Структура ценопопуляции Polypodium vulgare L. в урочище «Чертово Городище» Национального парка «Угра»//Вестн. ТвГУ. Сер.: Биология и экология. Вып. 16. С. 111-118.
- Уранов А.А. 1975. Возрастной спектр ценопопуляций как функция времени и энергетических волновых процессов//Биол. науки. № 2. С 7-34.
- Уранов А.А., Смирнова О.В. 1969. Классификация и основные черты развития популяций многолетних растений//Бюл. МОИП. Отд. биол. Т. 74. Вып. 2. С. 119-134.
- Цыганов Д.Н. 1983. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука. 196 с.
- Шорина Н.И. 1994. Экологическая морфология и популяционная биология представителей подкласса Polypodiidae: дис.. д-ра биол. наук. М. 359 с.
- Шорина Н.И., Державина Н.М. 1992. Структура и динамика ценопопуляционных скоплений Polypodium vulgare L. в лесах Западного Закавказья//Бот. журн. Т. 77. № 2. С. 46-54.
- Gams S.H. 1938. Ökologie der extratropischen Pteridophyten//Verdoorn F. (ed.). Manual of Pteridology. The Hague: M. Nijhoff. P. 382-419.
- Porter J.L. 1994. A study of the ecology of aspleniums in Limestone Grikes//Fern. Gaz. V. 14. № 7. P. 245-254.
