Эколого-фаунистический обзор пауков-волков (Aranei, Lycosidae) Северного Алтая
 Северного Алтая")
Автор: Волковский Евгений Владимирович
Журнал: Вестник Красноярского государственного педагогического университета им. В.П. Астафьева @vestnik-kspu
Рубрика: Биология
Статья в выпуске: 1 (19), 2012 года.
Бесплатный доступ
В результате проведенных стационарных исследований выявлен 21 вид семейства Lycosidae различающихся биотопов горных котловин Алтая, которая во многом определяется структурой растительного покрова и степенью теплообеспеченности биотопов. Установлено, что в большинстве биотопов формируются своеобразные комплексы, поэтому общность фаун имеет невысокую степень сходства.
Фенология, алтай
Короткий адрес: https://sciup.org/144153420
IDR: 144153420
Ecological and faunistic review of wolf-spiders (Aranei, Lycosidae) of the Northern Altai
As a result of stationary studies 21 species of Lycosidae family of different biotopes of mountain hollows of the Altai, which is mainly determined by the structure of the vegetative cover and the degree of the heat supply of habitats, are identified. It is established, that in the majority of biotopes the unique complexes are formed, so the community of faunas have a low degree of similarity.
Текст научной статьи Эколого-фаунистический обзор пауков-волков (Aranei, Lycosidae) Северного Алтая
Пауки выполняют важную роль в динамике популяций насекомых в различных биоценозах, поскольку являются неспецифическими хищниками [Шаров и др., 1984; Neffeler, 1999]. За последние 70 лет проведены значительные исследования по выявлению фауны пауков некоторых районов Западной Сибири [Харитонов 1932; 1936; Ermolajev, 1937; Лобанова, 1976; 1977; 1987; Еськов, 1988; Marusik et all, 1996; 2003; Романенко, 2007], в то же время исследований фаунистической и экологической направленности арахнонаселения гор Алтая еще недостаточно.
Важнейшими показателями для оценки роли пауков в биоценозах являются динамика численности и фенология развития, некоторые данные по ним содержатся в работах более общего характера. Специальные исследования проводились в основном на территории европейской части России, в то время как по Западной Сибири таких исследований практически нет. Отсутствуют не только региональные определители, но даже достаточно полные фаунистические списки.
Материалом для наших исследований послужили сборы, проведенные в окрестностях с. Чемал Республики Алтай и г. Бийска Алтайского края в течение вегетационных периодов 2003 и 2006 гг. Оба исследованных района имеют сходные черты по ландшафтным характеристикам, так как находятся в больших котловинах окруженных горами, а также обладают почти одинаковым по составу и структуре растительным покровом.
Сбор обитателей напочвенного яруса производился по общепринятым в арахнологии методикам, таким как ручной сбор и почвенные ловушки Барбера. В качестве ловушек использовались пластиковые стаканы емкостью 500 мл. В каждом биотопе устанавливалось по 5–10 ловушек с фиксатором, в роли которого мы использовали тосол. В отличие от формалина, в его состав входит многоатомный спирт (этиленгликоль), который на открытом воздухе практически не испаряется и препятствует отвердеванию покровов беспозвоночных. В зависимости от микрорельефа и грунта в биотопах использовалось разное количество ловушек, но не менее 5. Ловушки открывались лишь на 5 дней в течение декады. Это было необходимо для снижения влияния на наземную фауну беспозвоночных.
Сборы проводились в разнообразных биотопах, отличающихся составом растительности, экспозицией склонов и соответственно микроклиматическими условиями.
Биотоп 1. Зрелый густой сосновый лес с развитым подлеском и хорошо развитым травяным злаковым ярусом (при доминировании папоротника орляка). Горизонт А0 состоит из опавшей хвои, которая перегнивает более двух лет, и травянистых остатков. В результате этого формируется сравнительно мощный слой лиственного опада. В биотопе высота горизонта А0 составляет от 7 до 15 см. Средняя высота трав составляет 40 см, проекционное покрытие (пп) – до 90 %. Кустарниковый ярус в основном представлен акациями. Древесный ярус – зрелыми соснами, имеющими высокую сомкнутость крон (80 %). Лес расположен на не затапливаемом речными водами острове в русле реки Катунь.
Биотоп 2. Разнотравно-злаковый луг с редкими соснами, окруженный зрелым сосновым лесом. Из-за особенностей рельефа данный биотоп имеет слабо развитый слой лиственного опада. Этот биотоп находится ниже, чем окружающий рельеф, в результате чего дождевыми и снеговыми водами смывается его гумусовый слой. Средняя высота довольно густого травянистого яруса – 25 см. Кустарниковый ярус представлен единичными кустами акации и малины, произрастающими по краям биотопа. Территория исследования находится на горном склоне северо-западной экспозиции.
Биотоп 3. Лесные поляны, расположенные в лесном массиве на западном склоне горы. Слабо развит слой лиственного опада. Так как исследованный биотоп находится на склоне, происходит смыв этого опада к подножию горы. Относительно густой травостой – 20 см. Кустарниковый ярус отсутствует. Территория окружена зрелым сосновым лесом.
Биотоп 4. Зрелый густой сосновый лес со слабо развитым подлеском на холмистом ландшафте. В результате особенностей рельефа, способствующих смыву, формирование гумусового слоя при разреженном травяном ярусе затруднено. Проекционное покрытие травянистого покрова менее 50 %, средняя его высота 30 см. Кустарник отсутствует.
Биотоп 5. Зрелый сосновый лес с развитым подлеском, с явными следами низового пожара (трехлетней давности) на равнинной части рельефа. Биотоп обладает среднеразвитым горизонтом А 0 который сформирован в основном опавшей хвоей и останками травостоя. Достаточно густой травяной ярус со средней высотой 40 см. Кустарниковый ярус в основном состоит из акаций и небольшого количества подроста сосны.
Биотоп 6. Зрелый густой сосновый лес со слабо развитым подлеском и травяным ярусом. Слой лиственного опада развит слабо, составляет 1–3 см. Травянистая растительность редкая, средняя высота – 30 см. Кустарник представлен небольшим числом кустов акаций.
Биотоп 7. Сосново-березовый лес с хорошо развитым кустарниковым и травяным ярусами. Обладает сравнительно массивным слоем перегнивающей растительности (4–10 см). Достаточно густой (пп 80 %) травяной ярус – 40 см. Кустарник представлен в основном акцией и малиной, а также подростами березы и небольшого количества сосны.
Биотопы 1–5 находились в окрестностях с. Чемал, а биотопы 6 и 7 – в районе г. Бийска.
Наибольшее значение для биоценозов имеют массовые виды пауков. В исследуемых районах был обнаружен 21 вид из семейства Lycosidae, из которых Pardosa lugubris (Walckenaer, 1802) является самым многочисленным видом, встречающимся на протяжении всего вегетационного периода. В напочвенном ярусе в общей сложности в сборах было обнаружено 103 вида пауков (2489 экземпляров). На долю семейства Lycosidae приходится 1781 экземпляр.
В результате проведенных исследований нами было выявлено два пика динамической активности пауков семейства Lycosidae (рис.). Первый пик приходится на I декаду июня, второй отмечен во II декаде июля, и небольшое увеличение численности отмечено в I декаде августа.
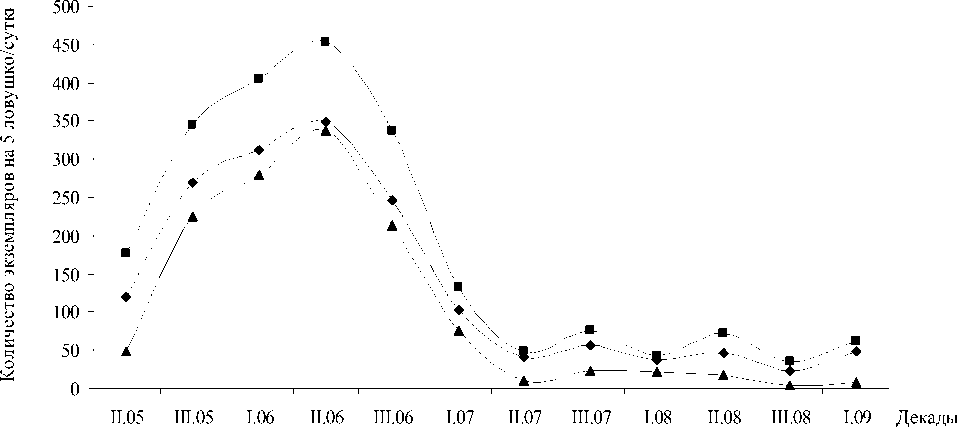
—■— Количество экземпляров пауков напочвенного яруса ♦ Количество пауков семейства Lycosidae
▲ Количество экземпляров вида Pardosa lugubris
Рис. Динамика численности пауков напочвенного яруса
Наиболее массовыми и распространенными видами являются Pardosa lugubris и Trochosa terricola [Волковский, 2010 а]. Они встречаются во всех исследованных нами биотопах. Alopecosa accentuata и Alopecosa pulverulenta отмечены нами в 5 биотопах, Alopecosa aculeata, Alopecosa cuneata, Alopecosa schmidti и Pardosa bi-fasciata встречаются в 3 различных биотопах, Pirata hygrophilus – в двух. Самым наименьшим распространением обладают Alopecosa albofasciata, Alopecosa inquili-na, Alopecosa mariae, Alopecosa sulzeri, Alopecosa taeniopus, Arctosa lutetiana, Par-dosa anchoroides, Pardosa paludicola, Pardosa palustris, Pardosa plumipes, Xerolyco-sa miniata и Xerolycosa nemoralis . Данные виды встречаются только в одном из исследованных биотопов.
Наибольшее количество видов семейства пауков-волков было отмечено нами в биотопах 1 и 3, несколько меньшим количеством видов обладают биотопы 2, 4 и 5 (по 9, 8 и 7 видов соответственно), самое наименьшее количество видов нами обнаружено в биотопах 6 и 7 (по 3 вида) (табл.).
При этом наибольшее количество видов приходится на род Alopecosa – 10 видов, Pardosa – 6, Xerolycosa – 2, а на Arctosa , Pirata и Trochosa – по 1.
Распределение экземпляров пауков-волков по биотопам неравномерно и значительно отличается от видового распределения.
Видовой состав семейства Lycosidae в течение вегетационного периода не одинаков [Волковский, 2010 б]. В результате проведенных исследований нами было выявлено три пика видового разнообразия пауков семейства Lycosidae. Первый пик приходится на I декаду июня, второй отмечен во II декаде июля и третий в первой декаде августа. При сопоставлении динамической активности пауков исследуемого семейства и его видового разнообразия выявлено, что пики активности и видового разнообразия совпадают. Возможно, это связано с однотипностью биологии развития большей части часто встречающихся видов пауков-волков.
Таблица
Биотопическое распределение видов Lycosidae в исследованных биотопах
|
№ п/п |
Вид |
Биотоп |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
||
|
1 |
Alopecosa accentuata (Latreille, 1817) |
+ |
+++ |
+ |
+ |
+++ |
||
|
2 |
Alopecosa aculeata (Clerck, 1758) |
+ |
+ |
+ |
||||
|
3 |
Alopecosa albofasciata (Brullé, 1832) |
+ |
||||||
|
4 |
Alopecosa cuneata (Clerck, 1758) |
+ |
+ |
+ |
||||
|
5 |
Alopecosa inquilina Clerck, 1758 |
+ |
||||||
|
6 |
Alopecosa mariae (Dahl, 1908) |
+ |
||||||
|
7 |
Alopecosa pulverulenta (Clerck, 1757) |
+ |
+ |
++ |
+ |
|||
|
8 |
Alopecosa schmidti (Hahn, 1835) |
+ |
+ |
+ |
||||
|
9 |
Alopecosa sulzeri (Pavesi, 1873) |
+ |
||||||
|
10 |
Alopecosa taeniopus (Kulczyn'ski, 1895) |
+ |
||||||
|
11 |
Arctosa lutetiana (Simon, 1876) |
+ |
||||||
|
12 |
Pardosa anchoroides Yu & Song, 1988 |
+ |
||||||
|
13 |
Pardosa bifasciata (C.L. Koch, 1834) |
+ |
+ |
+ |
||||
|
14 |
Pardosa lugubris (Walckenaer, 1802) |
+++ |
+ |
+++ |
++ |
+++ |
++ |
+++ |
|
15 |
Pardosa paludicola (Clerck, 1758) |
+ |
||||||
|
16 |
Pardosa palustris (Linnaeus, 1758) |
+ |
||||||
|
17 |
Pardosa plumipes (Thorell, 1875) |
+ |
||||||
|
18 |
Pirata hygrophilus Thorell, 1872 |
+ |
+ |
|||||
|
19 |
Trochosa terricola Thorell, 1856 |
+ |
+ |
+++ |
+ |
+++ |
+ |
+ |
|
20 |
Xerolycosa miniata (C.L. Koch, 1834) |
+ |
||||||
|
21 |
Xerolycosa nemoralis (Westring, 1861) |
+ |
||||||
Примечание:
+ – до 20 экземпляров;
++ – от 20 до 50 экземпляров;
+++ – более 50 экземпляров.
Вместе с тем исследованные биотопы обладают относительно разным видовым составом. Согласно индексу общности Чекановского – Сьеренсена, по количественным данным наибольшей степенью сходства обладают биотопы 1 и 6 (0,906±0,018). Чуть меньшим сходством обладают биотопы 6 и 7 (0,852±0,018), 1 и 7 (0,846±0,043),
1 и 4 (0,784±0,052), 4 и 6 (0,722±0,019), 4 и 7 (0,720±0,086), 2 и 5 (0,618±0,081), 3 и 5
(0,490±0,087), 5 и 7
(0,334±0,060), 3 и 7
(0,251±0,019), 2 и 3
(0,404±0,073), 5 и 6
(0,343±0,018), 1 и 5
(0,279±0,095), 1 и 3
(0,095±0,066), 2 и 6
(0,341±0,044), 4 и 5
(0,269±0,050), 3 и 6
(0,034±0,018), 1 и 2
(0,310±0,118), 3 и 4
(0,212±0,083), 2 и 7
(0,032±0,026), 2 и 4 (0,025±0,030).
Ряд исследователей [Зюзин, 1976; Измайлова, Вержутский, 1981], изучающих фауну и экологию пауков, указывают на важное значение в их распространении таких факторов, как: типологические особенности, световая структура насаждений, тепловой режим и степень увлажненности местообитаний. Наше исследование также подтверждает влияние экологических факторов на видовой состав и численность пауков различных биотопов. Кроме того, фауна пауков похожих лес- ных биотопов может иметь существенные различия, что указывает на своеобразие каждого из исследованных биотопов [Романенко, 1998].
В результате проведенных исследований на различающихся лесных и луговых биотопах нами был выявлен 21 вид пауков, относящихся к 6 родам семейства Lyco-sidae. Доминирующими видами являются Pardosa lugubris (72 % от общего количества пауков-волков), Alopecosa accentuata (11,2 %), Trochosa terricola (9,3 %) и Alopecosa pulverulenta (1,4 %). Эти виды составляют в наших сборах 93,9 % от всего количества экземпляров пауков-волков. При этом наибольшее количество видов нами было отмечено в биотопах 7 (33,2 % от общего количества экземпляров пауков-волков) и 5 (27 %). Выявлено три пика видового разнообразия пауков семейства Lycosidae. В процессе исследования наблюдалось два пика динамической активности ликозид, которые связаны с физиологическими процессами (половое созревание и поиск полового партнера) и заботой о потомстве (откладка яиц и формирование кокона с последующей заботой о его температурном режиме) [Волковский, 2010 б]. Установлено, что в большинстве исследованных биотопов формируются своеобразные комплексы видового состава пауков, поэтому общность фаун имеет невысокую степень сходства. В то же время встречаются биотопы с высокой степенью общности даже при значительном удалении друг от друга, что объясняется сходными элементами структуры растительного сообщества и некоторым совпадением экологических условий [Волковский, 2010 а].
