Эколого-генетическая оценка морфологических и биохимических признаков качества у образцов коллекции Brassica rapa L. ВИР

Автор: Артемьева А.М., Соловьева А.Е., Беренсен Ф.А., Кочерина Н.В., Чесноков Ю.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Род brassica: молекулярные маркеры и in vitro методы в селекции
Статья в выпуске: 1 т.52, 2017 года.
Бесплатный доступ
Особенность химического состава культур вида Brassica rapa L. (высокое содержание воды и низкое - жиров) обусловливает низкую калорийность капустных растений. Для них характерно высокое содержание углеводов и белков, включающих все незаменимые аминокислоты. Биохимический состав растений в пределах вида B. rapa сильно варьирует. Нами впервые проведена многофакторная комплексная оценка морфологических и биохимических признаков качества, обусловливающих потребительскую ценность растений из стержневой коллекции Brassica rapa L. Всероссийского института генетических ресурсов растений им. Н.И. Вавилова (ВИР, 96 образцов) в разных эколого-географических зонах (Южный Китай и Ленинградская обл.) в полевых условиях и в теплице. С помощью отобранных SSR (simple sequence repeats) маркеров, находящихся в неравновесном сцеплении с QTL морфологических и биохимических признаков, впервые осуществлен молекулярно-генетический анализ геномов различных (листовых, корнеплодных и масличных) форм у образцов из стержневой коллекции B. rapa ВИР, в результате чего среди культур вида выявлены новые генетические источники признаков качества. При изучении особенностей проявления некоторых признаков, обусловливающих потребительскую ценность растений вида B. rapa, установлено, что различия по морфометрическим показателям образцов (длина и ширина листовой платинки, размер черешка, опушенность, окраска) при выращивании в полевых условиях Южного Китая и Ленинградской области в среднем невелики, в то время как в теплице при загущенной посадке размеры листовой пластинки уменьшаются (у пекинской, китайской, ноздреватой капусты, репы, культурной сурепицы в полевых условиях листья были на 5-12 см длиннее и шире). По биохимическим признакам в полевых условиях обнаружены значимые и высокозначимые корреляции сухое вещество (СВ)-аскорбиновая кислота (АК) ( r = 0,51, p
Морфологические и биохимические признаки качества, молекулярные маркеры, скрининг коллекции
Короткий адрес: https://sciup.org/142214004
IDR: 142214004 | УДК: 635.127:631.524.022:631.524.5/.6:577.21 | DOI: 10.15389/agrobiology.2017.1.129rus
Ecological and genetic evaluation of morphological and biochemical characters of quality in Brassica rapa L. accessions from VIR collection
Peculiarity of chemical composition of Brassica rapa L. crops (high water content and low content of fats) determine their low-calorie character. They are notable for relatively high content of carbohydrates and proteins, including all essential amino acids. Biochemical composition varies greatly among B. rapa members. We are the first to carry out a multifactor evaluation of valuable morphological and biochemical traits of B. rapa accessions from the core collection of Vavilov Institute of Plant Genetic Resources (VIR, 96 samples) in eco-geographical study (South China and Leningrad region of Russia) under field trials and under a greenhouse conditions. By means of the developed SSR (simple sequence repeats) markers which are in linked disequilibrium with QTL of morphological and biochemical traits, the molecular genetic analysis was firstly carried out in leafy, rooted, and oilseed B. rapa genotypes from the VIR core collection. As a result, new genetic sources for quality parameters have been found among the samples investigated. In average, the samples did not differ noticeably on plant morphology (leaf length, leaf width, petiole size, hairiness, colour) in field tests when growing in South China and in Leningrad region, whereas in a greenhouse at higher plant density the leaf size decreased, e.g. in Chinese cabbage, pak-choi, wutacai, and turnip the leaves were 5-12 cm longer and wider in the field tests. Under field trials, we revealed the significant and high significant correlations between dry matter (DM) and ascorbic acid (AA) level ( r = 0.51, p < 0.05), DM and chlorophyll a (Chla) content ( r = 0.59, p < 0.05), DM and carotenoids (Cd) ( r = 0.55, p < 0.05), DM and carotins (Cn) ( r = 0.67, p < 0.05), DM and b-carotene (b-C) ( r = 0.59, p < 0.05), DM and protein (P) level ( r = -0.49, p < 0.05); Chla and Chlb ( r = 0.93, p < 0.001), Chla and Cd ( r = 0.59, p < 0.05), Chla and Cn ( r = 0.49, p < 0.05), Chla and b-C ( r = 0.99, p < 0.001); Chlb and b-C ( r = 0.92, p < 0.001); Cd and Cn ( r = 0.49, p < 0.05), Cd and b-C ( r = 0.63, p < 0.05); Cn and b-C ( r = 0.49, p < 0.05). In the greenhouse, the significant and high significant r values were as follows: r = -0.59 (p < 0.05) for DM and AA, r = 0.58 (p < 0.05) for DM and Chlb, r = -0.53 (p < 0.05) for DM and Cd, r = 0.71 (p < 0.001) for DM and Cn; r = -0.59 (p < 0.05) for AA and Chlb, r = 0.83 (p < 0.001) for AA and Cd, r = 0.58 (p < 0.05) for AA and P; r = 0.74 (p < 0.001) for Chla and Chlb, r = 0.67 (p < 0.05) for Chla and Cn, r = 0.95 (p < 0.001) Chla and b-C; r = -0.48 (p < 0.05) for Chlb and Cd, r = 0.87 (p < 0.001) for Chlb and Cn, r = 0.64 (p < 0.05) for Chlb and b-C; r = 0.63 (p < 0.05) for Cn and b-C. The semi-headed Chinese cabbage Syaobaikou and Dunganskaya, pak-choi Mayskaya, and especially Ching Pang Yu Tsain with a distinctly high level of chlorophylls and carotene, are indicated as new promising genetic sources for valuable biochemical parameters under both field and greenhouse conditions. Their indices in the field trials and greenhouse tests were 5.44-7.03 and 4.20-5.40 %, respectively, for DM (that is higher as compared to mean value for the crop), 12.61-24.66 and 24.12-33.23 % for P, 32.56-46.46 and 30.00-61.47 mg/100 g for AA, 33.35-110.64 and 53.82-95.99 mg/100 g for Chla, 12.20-53.80 and 17.89-44.78 mg/100 g for Chlb, and 1.80-6.75 and 3.00-6.04 mg/100 g for b-C. It is confirmed that the SSR markers BRMS051, KS51082, BRMS043 and KS50200 may effectively screen collection accessions and breeding material for desired morphological and biochemical traits. Our original data allow to practically implement an association mapping strategy and identify genetic determinants of morphological and biochemical quality characteristics using unique B. rapa collection preserved in VIR.
Текст научной статьи Эколого-генетическая оценка морфологических и биохимических признаков качества у образцов коллекции Brassica rapa L. ВИР
Репа ( Brassica rapa L.) — широко распространенный на Земном шаре вид, который включает экономически важные скороспелые и продуктив-
Работа выполнена при частичной финансовой поддержке гранта РФФИ ¹ 13-04-00128-а.
ные культуры (масличные, овощные, кормовые) с ценным химическим составом. Низкое содержание жиров и высокое — воды делает их малокалорийными, а значительное количество биологически активных веществ (витаминов, ферментов и др.) у культурных представителей вида положительно влияет на здоровье человека, стимулирует иммунную систему и препятствует развитию сердечно-сосудистых заболеваний. Во Всероссийском институте генетических ресурсов растений им. Н.И. Вавилова (ВИР) ведется поиск и привлечение в коллекции образцов B. rapa с улучшенными морфологическими и биохимическими признаками качества. Генетический анализ этих важных для человека количественных признаков (quantitative trait loci — QTL) возможен благодаря наличию у культур вида значительной генетической изменчивости. Биохимический состав растений в пределах вида B. rapa сильно варьирует (1-5). В последнее десятилетие установлены особенности накопления основных метаболитов, в том числе биологически активных веществ, у ранее малоизученных восточноазиатских капустных культур B. rapa , включая различные сортотипы (6, 7). Объяснение широчайшей изменчивости морфологических признаков у B. rapa предложено K. Lin с соавт. (8), обнаружившими уникальные гены у трех морфотипов B. rapa — корнеплодной репы и короткостадийной формы, геномы которых секвенировали и сравнили с геномом кочанной пекинской капусты.
Ранее (9-12) мы картировали локусы хромосом, отвечающие за проявление семи морфологических и пяти биохимических признаков качества в популяциях линий удвоенных гаплоидов листовых, корнеплодных и масличных культур вида B. rapa L. Всего (с учетом контроля одним локусом одних и тех же признаков в разных условиях выращивания или нескольких признаков одновременно) было картировано 140 QTL, детерминирующих формирование указанных хозяйственно значимых свойств у линий удвоенных гаплоидов B. rapa в полевых условиях и при выращивании в теплице. Установлены молекулярные маркеры, генетически сцепленные с изучаемыми QTL, дискутируется блочная геномная структура построения генетических компонент (локусов хромосом и групп сцепления), вовлеченных в экспрессию морфологических и биохимических признаков качества. Однако достигнутые результаты до конца не раскрывают характер наследования и молекулярно-генетический контроль этих признаков, представляющих интерес для изучения генетического разнообразия у вида B. rapa .
Отличительная особенность настоящей работы заключается в том, что впервые проведена многофакторная комплексная оценка морфологических и биохимических признаков качества, которые обусловливают потребительскую ценность растений B. rapa . Анализ геномов различных форм B. rapa с использованием молекулярно-генетических маркеров, находящихся в неравновесном сцеплении с QTL морфологических и биохимических признаков, позволил выявить перспективные образцы — генетические источники для селекции на качество у B. rapa . При этом найденные молекулярные маркеры могут служить эффективным инструментом при массовом скрининге образцов коллекции и селекционного материала.
Цель работы заключалась в морфологической, биохимической и молекулярно-генетической оценке коллекции Brassica rapa L. ВИР в различных эколого-географических зонах в полевых условиях и в теплице.
Методика . Исследовали 96 образцов разного происхождения из стержневой коллекции ВИР, отражающей ботаническое разнообразие вида B. rapa . Использовали таксономическое деление по C.E. Specht и A. Diederichsen (13), за исключением выделения ssp. sylvestris (Lam.) Janchen в отдельный от ssp. oleifera подвид. Все образцы оценивали в многолетних полевых испы-130
таниях в Пушкинском филиале ВИР (г. Санкт-Петербург) и на Южнокитайской опытной станции фирмы «Enza Zaden» (г. Гуанчжоу, провинция Гуандун, Китай) в 2010 году, а также в 2013-2016 годах в условиях зимней теплицы (Пушкинский филиал ВИР). Схема посева в полевых тестах 70½30 см, в теплице — 25½20 см. Морфологическое описание и биохимическую оценку растений проводили по представленным ранее методикам (3, 4, 14, 15), используя 20 растений каждого образца.
ДНК выделяли из молодых зеленых листьев растений по методике Д.Б. Дорохова и Э. Клоке (16). Молекулярно-генетический скрининг образцов методом полимеразной цепной реакции (ПЦР) проводили согласно описанному ранее протоколу (17). В реакционную смесь (25 мкл) вносили 10½ инкубационный буфер (2,5 мкл), 0,5 мкл каждого dNTP (10 мМ), по 1 мкл праймера (10 пмоль/мкл), 0,5 мкл Taq ДНК-полимеразы (5 ед/мкл) («Сибэнзим», Россия) и 20 нг геномной ДНК. ПЦР-амплификацию осуществляли в ДНК-термоциклере («Bio-Rad», США) с индивидуальной программой для каждого типа маркеров в соответствии с базой данных генетических маркеров овощных культур . Результаты визуализировали при помощи ДНК-электрофореза в 1,8 % агарозном геле с окрашиванием бромистым этидием. Для регистрации паттернов использовали документационную систему («Bio-Rad», США).
Данные обрабатывали с помощью программы Statistica 6.0 («StatSoft Inc.», США). Использовали уровень значимости p ≤ 0,05.
Результаты . Образцы коллекции B. rapa (табл. 1) оценивали в условиях теплицы (посев 1 февраля) и в полевых тестах (посев в Гуанчжоу 25 мая, в Пушкине 25 июня) по ряду морфологических признаков качества, обусловливающих продуктивность и потребительскую привлекательность растений: длина, ширина, окраска, опушенность, характер поверхности ткани листовой пластинки, а также длина и ширина черешка наиболее развитого листа среднего яруса.
1. Список образцов стержневой коллекции вида Brassica rapa L. из мировой коллекции Всероссийского института генетических ресурсов растений им. Н.И. Вавилова (ВИР )
|
Подвид, разновидность |
Сортотип |
Номер в каталоге ВИР |
Название |
Происхождение |
|
ssp. pekinensis (Lour.) Hanelt |
Дунганская |
139 |
Дунганская |
Казахстан |
|
(пекинская) |
Сяо |
53 |
Местный |
Казахстан |
|
74 |
Сяо-бай-коу |
Китай |
||
|
89 |
Доу-образная раннеспелая |
Китай |
||
|
Чиримен |
100 |
Hikoshima spring |
Япония |
|
|
Нагасаки |
238 |
Nagoya Market |
Япония |
|
|
Шантунг |
58 |
Би-це |
Киргизия |
|
|
210 |
Kiriba Santo |
Япония |
||
|
108 |
Местный |
Китай |
||
|
Касин |
132 |
Kasin |
Япония |
|
|
247 |
Хасинбечу |
Корея |
||
|
Чосен |
122 |
Лен-син-дзон |
Китай |
|
|
207 |
Chosen |
Япония |
||
|
Аити |
63 |
Местный |
Китай |
|
|
131 |
Aichi |
Япония |
||
|
Нозаки |
111 |
Nozaki early |
Япония |
|
|
327 |
Nozaki Harumaki |
Япония |
||
|
Кага |
103 |
Kaga |
Япония |
|
|
88 |
Цзюй-син-бао-тоу-бай-цай |
Китай |
||
|
Хоторен |
127 |
Hotoren |
Япония |
|
|
Чи-фу |
48 |
Wong-Bok |
Нидерланды |
|
|
110 |
Matsushima |
Япония |
||
|
Кенсин |
222 |
Kensin |
Япония |
|
|
Гранат |
164 |
Michihli |
Канада |
|
|
71 |
Хэ-тоу-вень |
Китай |
||
|
Да-цин-коу |
56 |
Да-цин-коу |
Китай |
|
|
128 |
Цужита |
Япония |
||
|
198 |
Местный |
Китай |
||
Продолжение таблицы 1
|
ssp. chinensis (L.) Hanelt |
Пиорбай |
75 |
Пиорбай |
Китай |
|
(китайская) |
Сыюсман |
77 |
Сыюсман |
Китай |
|
Вр.930 |
Майская 8 |
Китай |
||
|
Тайсай |
46 |
Тай-на |
Россия |
|
|
106 |
Янцай |
Китай |
||
|
214 |
Nicanme Jukijiro Taisai |
Япония |
||
|
Ю-тсай (var. utilis) |
195 |
Местный |
Китай |
|
|
203 |
Ching Pang Ju Tsai |
Китай |
||
|
var. rosularis (Tsen & Lee) |
Та-гу-цай |
84 |
Хэе-ю-та-цай |
Китай |
|
Hanelt (розеточная) |
129 |
Та-гу-цай |
Китай |
|
|
var. narinosa (Bailey) Hanelt |
Хризантемум |
154 |
Chrysanthemum heart |
Китай |
|
(ноздреватая) |
213 |
Bitamin na |
Япония |
|
|
var. purpuraria (Bailey) Bailey |
||||
|
(пурпурная) |
391 |
Xing Yang |
Китай |
|
|
ssp. nipposinica (Bailey) Hanelt Мибуна |
115 |
Mibuna |
Япония |
|
|
(японская) |
Мизуна |
159 |
Mizuna |
Япония |
|
241 |
Shiroguki Kyona |
Япония |
||
|
ssp . rapa L. f. Komatsuna |
Комацуна |
215 |
Uzuki Komatsuna |
Япония |
|
(листовая репа комацуна) |
242 |
Goseki Late |
Япония |
|
|
Курона |
264 |
Kurona |
Япония |
|
|
Японские листовые овощи |
Мана |
372 |
Bansei Mana |
Япония |
|
Сирона |
98 |
Osaka Market |
Япония |
|
|
217 |
Okute Osaka Shirona |
Япония |
||
|
Хиросимана |
335 |
Hiroshimana |
Япония |
|
|
Стабильные гибриды между |
96 |
Шантай |
Китай |
|
|
подвидами |
302 |
Гурин Дэбюу |
Япония |
|
|
331 |
White Long Petiole |
Япония |
||
|
436 |
Benrina |
Япония |
||
|
ssp . rapa L. |
Китайский |
163 |
Местный |
Китай |
|
(репа корнеплодная) |
Остерзундомский |
307 |
Остерзундомский |
Россия |
|
Бортфельдский |
385 |
Бортфельдский |
Украина |
|
|
Карельская |
738 |
Карельская |
Россия |
|
|
Гробовская |
821 |
Гробовская |
Россия |
|
|
Миланская белая |
826 |
Миланская белая |
Россия |
|
|
Петровская |
830 |
Петровская |
Россия |
|
|
Тельтовский |
894 |
Тельтовская |
Германия |
|
|
Норфолькский |
||||
|
фиолетовоголовый |
984 |
Норфолькский |
Франция |
|
|
Волынский |
1050 |
Волынский |
Украина |
|
|
Золотой шар |
1283 |
Золотой шар |
Нидерланды |
|
|
ssp . oleifera (DC.) Metzger |
68 |
Местный |
Китай |
|
|
f. annua (сурепица яровая) |
1 |
Kun Min ai u-zai |
Китай |
|
|
2 |
Hue Zin u-zai |
Китай |
||
|
11 |
Gute |
Финляндия |
||
|
13 |
Местный |
Аргентина |
||
|
25 |
Zsjan Su U uan-uzai 5082 |
Китай |
||
|
63 |
Pahsi |
Индия |
||
|
106 |
Lotni mustard |
Индия |
||
|
108 |
Arlo |
Швеция |
||
|
114 |
Local (tetraploid) |
Пакистан |
||
|
163 |
LGL |
Пакистан |
||
|
192 |
Mustard |
Непал |
||
|
248 |
Local |
Испания |
||
|
251 |
Vat-cawte |
Танзания |
||
|
301 |
BHLS |
Непал |
||
|
339 |
Jui-cai-tai |
Китай |
||
|
374 |
Local 88/47 |
Бутан |
||
|
ssp . oleifera (DC.) Metzger |
166 |
Root mustard |
Тунис |
|
|
f. biennis (сурепица озимая) |
337 |
U-zai-zsi |
Китай |
|
|
ssp . dichotoma (Roxb.) Hanelt |
53 |
Local toria |
Индия |
|
|
(коричневый сарсон) |
100 |
Local |
Непал |
|
|
135 |
Ds 17 |
Индия |
||
|
161 |
Toria selection |
Пакистан |
||
|
205 |
Sarson |
Пакистан |
||
|
ssp . trilocularis (Roxb.) Hanelt |
131 |
Type 1 |
Индия |
|
|
(желтый сарсон) |
188 |
Palton sarson 66 |
Индия |
|
|
299 |
Sangam |
Индия |
||
|
338 |
Chen-du-ai-u-zai |
Китай |
||
|
ssp . sylvestris (Lam.) Janchen |
176 |
Италия |
||
|
(сурепица лесная дикая) |
218 |
Nabo silvestre |
Перу |
|
Установлено, что средняя по коллекции длина и ширина листовой пластинки в полевых условиях в Китае и при выращивании в Пушкин-132
ском филиале ВИР различались несущественно, тогда как в теплице размеры листовой пластинки оказались значительно меньше (табл. 2). В то же время в условиях Китая размеры листовой пластинки были существенно больше у китайской капусты, ноздреватой и пурпурной капусты (все культуры южно-китайского происхождения), а также у корнеплодной репы; незначительно больше — у южно-китайской розеточной капусты и сурепицы. Размеры листовой пластинки у пекинской капусты и листовой репы, а также длина пластинки у японской капусты оказались больше при выращивании в условиях Ленинградской области. Очевидно, это вызвано, очень высокими летними температурами в Южном Китае (35 ° С и выше), лимитирующими быстрый рост вегетативных органов у некоторых культур. У пекинской капусты размеры листовой пластинки достоверно превышали средние по популяции: в полевых тестах в Китае, Пушкине и в условиях теплицы длина составляла соответственно 38,22±1,61; 41,82±1,03 и 31,87±0,71 см, ширина — 19,33±0,80; 25,02±0,78 и 16,02±0,45 см. Размер листа у листовой репы в полевых условиях также превышал средние значения по коллекции, в теплице — не отличался от среднего для вида. Листовая пластинка у культурной сурепицы была значительно меньше (на 3040 % независимо от условий выращивания), чем в среднем по популяции, а дикая сурепица и розеточная капуста выделялись наименьшими размерами. Листовые пластинки у пекинской, китайской, ноздреватой капусты, репы, культурной сурепицы в полевых условиях были на 5-12 см длиннее и шире, чем в теплице, у розеточной и японской капусты, а также дикой сурепицы в тех же условиях — практически не различались по размеру.
-
2. Средние размеры листа у культур вида Brassica rapa L. в контрастных условиях
-
3. Образцы вида Brassica rapa L., выделившиеся по размеру продуктовых органов в контрастных условиях
Образец, номер по каталогу ВИР
Длина пластинки, см полевой опыт П,
К I П теплица
Ширина пластинки, см полевой опыт П,
К I П теплица
Длина черешка, см полевой опыт П,
К I П теплица
Kiriba Santo, к-210
Chosen, к-207 Среднее по культуре
Тай-на, к-46
Nicanme Jukijiro
Taisai, к-214
Среднее по культуре
Goseki Late, к-242
Bansei Mana, к-372 Среднее по культуре
П е ки н с к ая к апу ст а (бесчерешковая культура) 55,0±2,6 45,9±3,3 36,3±4,2 22,0±1,7 22,6±1,5 14,8±1,2 50,6±2,8 42,1±2,1 37,9±2,7 25,6±2,4 20,2±1,9 18,1±2,6 38,2±1,6 41,8±1,0 31,8±0,7 19,3±0,8 25,0±0,8 16,0±0,4
Китайская капуста
24,2±2,3 22,8±2,7 16±3,1 19,0±2,6 18,2±2,2 13,8±2,8 13±1,8 22,8±3,0
26,5±2,8 23,3±2,5 12,9±2,8 21,5±1,4 18,1±1,8 10,3±1,6 25,4±2,4 19,5±2,7
26,1±2,5 21,3±1,1 16,1±1,2 19,2±1,3 16,3±1,1 11,2±0,6 16,0±1,8 15,4±1,7
Листовая репа
40,3±3,7 40,5±2,9 25,9±3,2 21,5±2,8 23,1±2,0 9,5±1,9 16,6±2,3 11,5±2,2
50,4±3,2 45,2±3,6 22,6±2,6 36,8±3,4 26,9±2,6 15,4±1,4 9,1±2,1 9,8±2,1
27,6±2,1 33,8±2,6 21,8±1,0 18,7±1,2 23,5±1,3 12,4±0,7 14,8±1,8 12,7±1,7
16,7±2,5
17,1±2,0
12,3±1,3
9,6±1,6
8,4±1,2
11,1±0,9
П р и м еч а ни е. К — Южный Китай, г. Гуанчжоу (провинция Гуандун), П — Пушкинский филиал
Всероссийского института генетических ресурсов растений им. Н.И. Вавилова — ВИР, г. Санкт-Петербург. Полевые опыты выполняли в Китае в 2010 году, в Пушкине в 1997-2016 годах, эксперименты в условиях теплицы — в 2013-2016 годах. Латинские обозначения культур см. в таблице 1. Результаты получены при анализе 96 образцов из стержневой коллекции ВИР. Приведены средние ( Х ) и ошибки средних (±S x ).
|
Длина, см |
Ширина, см |
|||||
|
Культура |
полевой опыт |
П, |
полевой опыт |
П, |
||
|
К |
П |
теплица |
К |
П |
теплица |
|
|
Ли |
стовая пластинка |
|||||
|
Пекинская капуста |
38,22±1,61 |
41,82±1,03 |
31,87±0,71 |
19,33±0,80 |
25,02±0,78 |
16,02±0,45 |
|
Китайская капуста |
26,10±2,58 |
21,36±1,08 |
16,13±1,16 |
19,20±1,30 |
16,33±1,06 |
11,18±0,58 |
|
Розеточная капуста |
14,50±0,50 |
12,30±2,00 |
13,93±1,11 |
11,50±0,50 |
10,45±1,05 |
10,05±1,03 |
|
Ноздреватая и пурпурная |
||||||
|
капуста |
24,70±1,86 |
21,50±1,98 |
16,73±0,42 |
22,30±2,61 |
18,60±0,38 |
11,17±0,42 |
|
Японская капуста |
18,00±3,06 |
24,63±2,20 |
24,10±7,03 |
9,67±2,40 |
9,50±3,10 |
8,37±1,19 |
|
Репа |
24,78±3,14 |
19,12±1,39 |
19,10±0,83 |
19,00±1,78 |
12,26±0,91 |
15,95±0,95 |
|
Листовая репа |
27,58±2,09 |
33,84±2,63 |
21,79±1,01 |
18,75±1,22 |
23,51±1,32 |
12,45±0,67 |
|
Сурепица |
19,04±1,13 |
16,67±0,52 |
12,75±0,48 |
13,72±0,64 |
12,96±0,47 |
10,38±0,40 |
|
Сурепица дикая |
13,50±0,50 |
11,50±1,00 |
14,03±2,01 |
11,50±0,50 |
9,50±0,50 |
11,85±1,86 |
|
Среднее по коллекции |
25,75±1,09 |
26,34±1,20 |
20,63±0,85 |
16,92±0,53 |
17,61±0,67 |
12,85±0,35 |
|
НСР 05 |
3,31 |
3,62 |
2,55 |
1,60 |
2,04 |
1,05 |
|
Черешок |
||||||
|
Китайская капуста |
16,00±1,79 |
15,41±1,74 |
12,35±1,35 |
3,33±1,39 |
3,39±0,23 |
2,06±0,09 |
|
Розеточная капуста |
9,00±2,01 |
10,50±0,10 |
12,28±1,65 |
2,20±0,25 |
1,95±0,35 |
1,51±0,09 |
|
Ноздреватая и пурпурная |
||||||
|
капуста |
21,00±1,53 |
13,87±1,25 |
15,73±1,73 |
3,67±0,34 |
3,40±0,15 |
1,64±0,08 |
|
Японская капуста |
16,30±2,17 |
14,43±0,66 |
15,76±3,06 |
0,97±0,03 |
1,10±0,15 |
1,03±0,09 |
|
Репа |
21,25±1,36 |
23,39±1,04 |
30,90±2,54 |
2,00±0,29 |
1,63±0,15 |
1,44±0,09 |
|
Листовая репа |
14,80±1,84 |
12,68±1,66 |
11,11±0,95 |
2,90±0,28 |
3,81±0,32 |
1,97±0,14 |
|
Сурепица |
16,20±0,85 |
15,63±0,58 |
14,71±0,80 |
1,10±0,07 |
1,16±0,10 |
0,71±0,05 |
|
Сурепица дикая |
14,00±3,01 |
10,50±0,50 |
15,15±3,16 |
0,90±0,10 |
0,75±0,25 |
0,64±0,13 |
|
Среднее по коллекции |
16,53±0,64 |
16,07±0,65 |
16,33±1,37 |
1,98±0,14 |
2,04±0,13 |
1,29±0,07 |
|
НСР 05 |
1,93 |
1,95 |
4,11 |
0,43 |
0,41 |
0,22 |
|
П р и м е ч а н и е. К — Южный Китай, |
г. Гуанчжоу |
(провинция |
Гуандун), П — Пушкинский филиал |
|||
Всероссийского института генетических ресурсов растений им. Н.И. Вавилова — ВИР, г. Санкт-Петербург. Полевые опыты выполняли в Китае в 2010 году, в Пушкине в 1997-2016 годах, эксперименты в условиях теплицы — в 2013-2016 годах. Латинские обозначения культур см. в таблице 1. Результаты получены при анализе 96 образцов из стержневой коллекции ВИР. Приведены средние ( Х ) и ошибки средних (±S x ).
Средняя по популяции длина черешка при всех условиях выращивания была стабильной, ширина черешка в условиях юга Китая и Ленинградской области разнились незначительно, но в теплице оказалась суще- ственно меньше (см. табл. 2). У пурпурной и ноздреватой капусты, японской капусты, листовой репы и дикой сурепицы длина черешка в условиях юга Китая была значительно больше, чем в Ленинградской области, у культурной сурепицы — незначительно больше, у репы и розеточной капусты — меньше. В теплице у китайской капусты, листовой репы и сурепицы черешок был короче, чем в поле, у остальных культур — длиннее (у репы — в 2 раза длиннее среднего по популяции). Все образцы пекинской капусты имели цельные сидячие листья без черешка. Длинный, относительной широкий черешок был характерен для корнеплодной репы, ноздреватой и японской капусты (на 15-30 % длиннее среднего значения по коллекции), короткий — для розеточной капусты. У образцов длина черешка в полевых условиях и в теплице в основном различалась незначительно (на 1-4 см), за исключением японской капусты и корнеплодной репы, у которых черешок в теплице оказался на 10-15 см длиннее, чем в полевых условиях. Самый широкий черешок обнаружили у образцов китайской и ноздреватой капусты и листовой репы, тонкий — у японской капусты и сурепицы. В полевых тестах черешок был достоверно шире у всех культур, за исключением японской капусты и дикой сурепицы.
Поверхность листовой пластинки у образцов коллекции различалась в зависимости от ботанической принадлежности генотипа — от гладкой до складчатой или слабо-, средне- и сильноморщинистой, характер опушения варьировал (от отсутствия до сильной степени проявления), окраска пластинки изменялась от светло- до темно-зеленой.
В стержневой коллекции в качестве генетических источников мы выделили образцы, у которых размеры продуктовых органов были достоверно выше среднепопуляционных показателей (табл. 3). При этом у ряда образцов в контрастных условиях испытания длина и ширина листовых пластинок и черешков стабильно превышала средние значения для культуры, что свидетельствует о высокой адаптивной способности. Незначительную изменчивость по этим признакам отмечали у образцов пекинской капусты Kiriba Santo, Chosen, китайской капусты Тай-на, Nicanme Jukijiro Taisai, местной китайской репы (к-163), формирующей товарный корнеплод и листовую розетку высокого салатного качества. Настоящее исследование подтвердило, что для выращивания в теплице более подходят листовые и полукочанные сорта пекинской капусты в отличие от типично кочанных сортов, о чем сообщалось ранее (6); адаптивные свойства коллекционных образцов других культур вида при сравнительных испытаниях в теплице и в полевых условиях нами оценены впервые.
Отсутствие опушения и яркая окраска повышают потребительские качества листовых культур B. rapa . У выделившихся образцов опушение отсутствовало или проявлялось очень слабо, а окраска листовой пластинки варьировала от ярко-светло-зеленой до ярко-темно-зеленой.
Биохимические исследования показали (табл. 4), что содержание сухого вещества в среднем по стержневой коллекции вида B. rapa в полевых условиях составляло 7,77±0,25 % и достоверно превышало аналогичный показатель в теплице — 5,36±0,15 %, причем у всех культур, кроме пекинской капусты, превышение было значительным — на 30-140 % (см. табл. 4). Содержание сухого вещества достоверно различалось у разных культур вида и колебалось от 5,65 % у пекинской капусты до 11,24 % у листовой репы в полевых опытах и от 4,44 % у китайской капусты до 6,62 % у репы в теплице. Таким образом, амплитуда изменчивости по содержанию сухого вещества в полевых условиях была значительно больше, чем в теплице.
-
4. Содержание (Х±Sх) питательных и биологически активных веществ у культур вида Brassica rapa L. при разных условиях произрастания
Культура
Сухое вещество, %
Белок, %
Аскорбиновая кислота, мг/100 г
Хлорофилл, мг/100 г
Каротиноиды, мг/100 г
β -Каротин, мг/100 г
а
b
Капуста:
пекинская
5,65±1,47
В п о л е в ы х у с л о в и я х (2006-2016 годы)
23,73±3,86 50,45±14,82 26,21±12,04 11,72±5,94
10,30±4,28
1,82±0,85
китайская
9,99±1,91
24,85±2,73
65,21±13,36
64,51±11,96 29,03±9,87
17,60±3,62
4,12±0,67
розеточная
9,41±1,98
27,80±3,24
58,03±13,69
98,87±9,06
55,46±6,79
18,74±2,05
6,34±0,63
ноздреватая и пурпурная
8,36±3,29
29,09±2,00
46,24±5,93
74,71±4,91
37,69±6,83
16,18±2,58
4,56±0,19
японская
9,17±0,10
23,49±3,01
46,24±1,36
86,47±6,86
40,73±5,54
21,43±3,50
5,39±0,26
Репа: корнеплодная, листья
8,81±1,62
24,66±3,82
71,91±10,29
78,26±25,09 43,16±20,32
15,56±4,75
4,89±1,53
листовая
11,24±3,67
21,35±1,28
77,52±10,26
98,61±4,12
47,15±3,83
20,71±10,1
6,24±0,83
Сурепица
7,45±1,34
27,95±3,13
69,74±21,15
81,83±11,69 47,29±14,08
12,99±4,30
5,05±0,89
Сурепица дикая
10,64±0,73
17,99±1,04
68,68±4,80
75,25±0,64
41,09±1,09
10,77±1,02
4,55±0,03
Среднее
по коллекции
7,77±0,25
24,87±0,39
58,70±1,65
59,06±3,27
30,63±2,09
14,06±0,55
3,77±0,19
НСР 05
0,77
1,17
4,95
9,83
6,29
1,64
0,56
Капуста:
пекинская
4,84±0,08
31,06±0,48
В теплиц
35,00±1,44
е (2014 год)
48,15±2,54
18,23±1,32
23,07±0,76
2,97±0,16
китайская
4,44±0,13
24,12±0,98
30,00±4,21
73,30±4,49
36,03±2,88
27,85±3,78
4,46±0,30
розеточная
4,72±0,28
23,65±0,90
19,00±3,01
63,22±0,78
33,34±0,33
20,46±0,56
4,09±0,01
ноздреватая и пурпурная
5,40±0,18
20,46±0,58
19,00±1,15
64,11±5,09
36,77±3,63
16,93±3,38
3,78±0,31
японская
5,44±0,12
27,23±1,51
38,00±5,20
87,60±6,67
37,41±3,47
36,39±2,42
5,45±0,38
Репа:
корнеплодная, листья
6,62±0,17
25,32±0,67
15,00±0,63
77,26±4,03
50,29±3,46
14,65±1,08
4,70±0,26
листовая
4,46±0,23
27,36±0,98
37,00±4,54
57,62±5,23
27,85±6,78
25,19±2,81
3,56±0,30
Сурепица
6,08±0,14
27,69±0,55
20,00±0,53
65,70±1,45
46,11±2,37
11,79±0,56
4,02±0,07
Сурепица дикая
5,80±0,28
25,32±2,98
21,00±0,71
78,24±9,88
56,08±12,01
15,28±4,28
4,73±0,49
Среднее
по коллекции
5,36±0,10
27,50±0,40
27,03±1,16
63,24±1,71
35,15±1,72
19,77±0,94
3,88±0,10
НСР 05
0,29
1,2
3,47
5,12
5,16
2,81
0,31
П р и м е ч а н и е. Содержание белка указано в расчете на сухую, биологически активных веществ – на
сырую массу. Испытания проводились в Пушкинском филиале Всероссийского института генетических ресурсов растений им. Н.И. Вавилова (ВИР, г. Санкт-Петербург). Латинские обозначения культур см. в таблице 1. Результаты получены при анализе 96 образцов из стержневой коллекции ВИР.
По белку мы не выявили подобных закономерностей: в полевых опытах его содержание было значительно ниже среднего по коллекции у дикой сурепицы и значительно выше — у ноздреватой и пурпурной капусты (см. табл. 4), а в теплице у ноздреватой капусты, наоборот, отмечали 135
наименьший показатель при наибольшем у пекинской капусты. В среднем по коллекции содержание белка повышалось в теплице, однако размах изменчивости признака (11 %) сохранялся независимо от условий произрастания. У китайской капусты, репы и сурепицы содержание белка при сравнительных испытаниях было практически стабильным, у пекинской и японской капусты, листовой репы и сурепицы — оказалось выше в теплице, у розеточной, ноздреватой и пурпурной капусты — в полевых условиях. В отношении последней группы растений причина, возможно, связана с их относительно медленным ростом или с характерным проявлением анализируемого признака, общим для указанных ботанически близких культур.
Культуры вида B. rapa — важные источники аскорбиновой кислоты, каротиноидов, хлорофиллов. В наших исследованиях содержание аскорбиновой кислоты у образцов B. rapa варьировало достаточно широко: минимальные значения соответствовали средним для белокочанной капусты, максимальные превосходили их в несколько раз. В среднем по коллекции показатели были в 2 раза выше в полевых условиях (58,7±1,65 против 27,03±1,16 мг/100 г), и эту закономерность отмечали для всех культур, за исключением японской капусты, у которой разница оказалась не такой значительной. Напротив, содержание аскорбиновой кислоты у розеточной и ноздреватой капусты, корнеплодной репы и сурепицы при выращивании в поле было выше в 3 раза и больше, чем в теплице. Наибольшее накопление аскорбиновой кислоты в поле наблюдали у китайской капусты, репы и сурепицы, в теплице — у пекинской и японской капусты и листовой репы.
В среднем по коллекции каротиноиды достоверно активнее накапливались в условиях теплицы (19,77±0,94 против и 14,06±0,55 мг/100 г, НСР05 соответственно 2,81 и 1,64). Это особенно важно в зимний и весенний периоды, когда пища бедна витаминами. Самым богатым источником каротиноидов были тепличные растения японской капусты, затем следовала китайская капуста, дикая сурепица и листовая репа. В полевых условиях по этому признаку выделялись образцы японской, китайской, розеточной капусты и листовой репы. У пекинской капусты и дикой сурепицы превышение количества каротиноидов в теплице было более чем 2-кратным относительно регистрируемого в поле. У розеточной, ноздреватой и пурпурной капусты, сурепицы и репы количество каротиноидов оказалось относительно стабильным. Примерно 20 % фракции каротиноидов составляют каротины. Собственно каротин на 80-90 % представлен β -формой. В среднем содержание β -каротина мало различалось (в теплице и в поле — соответственно 3,88±0,1 и 3,77±0,19 мг/100 г). Его накопление было стабильным у китайской и японской капусты, репы, дикой сурепицы, у пекинской капусты оно было достоверно выше в теплице (2,97±0,16 против и 1,82±0,85 мг/100 г, НСР05 соответственно 0,56 и 0,31), у розеточной, ноздреватой и пурпурной капусты, листовой репы — в поле, а максимальным накоплением характеризовалась японская, китайская и розеточная капуста.
Хлорофилл играет значительную роль в диетическом питании: употребление в пищу зеленых листьев повышает количество гемоглобина и эритроцитов в крови. В среднем по коллекции содержание хлорофиллов a и b было несущественно выше в теплице (сумма 63,24±1,71 и 35,15±1,72 против суммы 59,06±3,27 и 30,63±2,09 мг/100 г). Практически стабильное содержание хлорофиллов (особенно хлорофилла а) при обоих способах выращивания отмечали у японской капусты, репы и дикой сурепицы, повышение показателя в теплице — у пекинской и китайской капусты, в поле — у розеточной и ноздреватой капусты, листовой репы и сурепицы. При недостатке свежих салатных овощей в зимне-весенний период в теплицах 136
выращивают главным образом быстро растущие культуры (пекинскую и китайскую капусту), ценность которых, таким образом, повышается благодаря способности накапливать зеленые пигменты в этих условиях. В качестве тепличных культур можно также рекомендовать розеточную, ноздреватую, японскую капусту и листовую репу, значительно превосходящие пекинскую капусту по содержанию хлорофиллов (их максимальное количество отмечали у розеточной и японской капусты и листовой репы).
Проведенный корреляционный анализ биохимических признаков показал, что количество сухого вещества достоверно и однонаправленно связано с содержанием хлорофиллов, каротинов и β -каротина, независимо от условий культивирования (табл. 5). Взаимозависимость между содержанием сухого вещества и аскорбиновой кислоты, а также каротиноидов была существенной, но разнонаправленной (прямая — в полевых условиях, обратная — в теплице). В полевых условиях выявили значимую обратную связь между содержанием сухого вещества и белка, в условиях теплицы — между содержанием аскорбиновой кислоты и хлорофилла b (обратная зависимость), каротиноидов, белка (прямая зависимость). Высокодостоверной была взаимозависимость между количеством хлорофиллов a и b, а также их связь с накоплением каротиноидов и каротинов (особенно β -каротина). Существенную корреляцию отмечали между содержанием каротинов и β -каротина. Таким образом, образцы, сочетающие высокое содержание хлорофиллов и каротинов, можно получить достаточно легко, тогда как селекция на одновременно высокое содержание белка и аскорбиновой кислоты будет вызывать затруднения.
5. Коэффициенты корреляции между биохимическими признаками качества у культур вида Brassica rapa L. при разных условиях произрастания
|
Показатель Показатель |
СВ |
АК |
Ха |
Хb |
Кд |
Ка |
β -К |
Б |
|
В п о л е в ы х у с л о в и я х (2006-2016 годы) |
||||||||
|
Сухое вещество (СВ) |
0,51* |
0,59* |
0,36 |
0,55* |
0,67* |
0,59* |
- 0,49* |
|
|
Аскорбиновая кислота (АК) |
0,29 |
0,26 |
- 0,02 |
0,33 |
0,30 |
- 0,39 |
||
|
Хлорофилл а (Ха) |
0,93** |
0,59* |
0,49* |
0,99** |
0,16 |
|||
|
Хлорофилл b (Хb) |
0,32 |
0,37 |
0,92** |
0,28 |
||||
|
Каротиноиды (Кд) |
0,49* |
0,63* |
0,09 |
|||||
|
Каротины (Ка) |
0,49* |
0,01 |
||||||
|
β -Каротин ( β -К) |
0,16 |
|||||||
|
Белок (Б) |
||||||||
|
В т е п л и ц е (2014 год) |
||||||||
|
Сухое вещество (СВ) |
- 0,59* |
0,31 |
0,58* |
- 0,53* |
0,71** |
0,36 |
- 0,02 |
|
|
Аскорбиновая кислота (АК) |
- 0,11 |
- 0,59* |
0,83** |
- 0,42 |
- 0,09 |
0,58* |
||
|
Хлорофилл а (Ха) |
0,74** |
0,21 |
0,67* |
0,95** |
- 0,31 |
|||
|
Хлорофилл b (Хb) |
- 0,48* |
0,87** |
0,64* |
- 0,37 |
||||
|
Каротиноиды (Кд) |
- 0,36 |
0,29 |
0,22 |
|||||
|
Каротины (Ка) |
0,63* |
- 0,18 |
||||||
|
β -Каротин ( β -К) |
- 0,25 |
|||||||
Белок (Б)
П р и м е ч а н и е. Испытания образцов из стержневой коллекции ВИР проводились в Пушкинском филиале Всероссийского института генетических ресурсов растений им. Н.И. Вавилова (ВИР, г. Санкт-Петербург). *, ** Значимые (p < 0,05) и высокозначимые (p < 0,001) коэффициенты корреляции.
На основании полученных данных мы определили новые генетические источники изучаемых биохимических признаков при выращивании в полевых условиях и в теплице с содержанием сухого вещества соответственно 5,44-7,03 и 4,20-5,40 % (выше среднего значения для культуры), белка — 12,61-24,66 и 24,12-33,23 %, аскорбиновой кислоты — 32,56-46,46 и 30,00-61,47 мг/100 г, хлорофилла a — 33,35-110,64 и 53,82-95,99 мг/100 г, хлорофилла b — 12,20-53,80 и 17,89-44,78 мг/100 г, β -каротина — 1,80-6,75 и 3,00-6,04 мг/100 г. Это образцы полукочанной пекинской капусты Сяо-бай-коу и Дунганская, китайской капусты Майская и Ching Pang Yu Tsai, вы-137
делившийся особенно высоким содержанием хлорофиллов и каротина.
Для молекулярно-генетических исследований отобрали 8 SSR (simple sequence repeats) маркеров, пригодных для последующего скрининга образцов мировой коллекции. Маркеры располагались в A03 (3 маркера), A05 (1 маркер), A06 (2 маркера) и A09 (2 маркера) хромосомах, охватывая группы сцепления, на которых, по данным выполненного нами QTL-анализа (1012, 18), размещаются локусы, контролирующие морфологические и био-
А
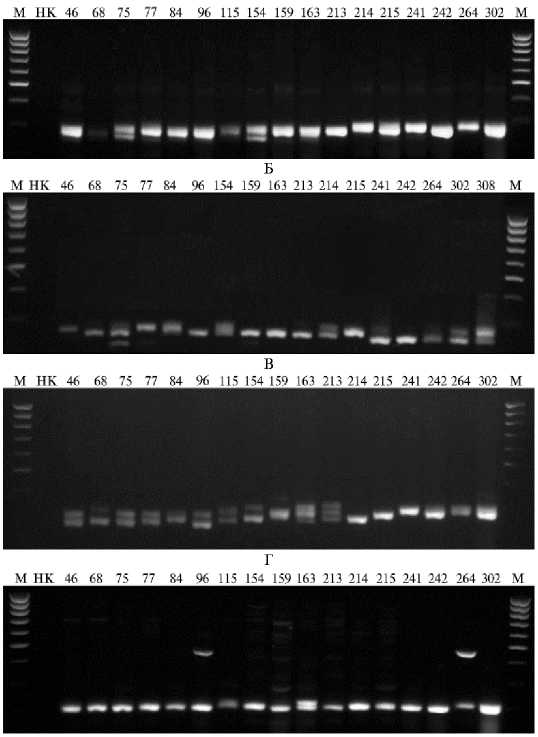
Результаты ПЦР-анализа образцов вида Brassica rapa L. SSR (simple sequence repeats) маркерами BRMS051 (А) , KS51082 (Б) , BRMS043 (В) и KS50200 (Г) : 46, 68, 75, 77, 84, 96, 115, 154, 159, 163, 213, 214, 215, 241, 242, 264, 302 и 308 — образцы (номера по каталогу ВИР), НК — отрицательный контроль, М — маркер молекулярной массы М16-ДНК-маркер 100 bp (10 фрагментов от 100 до 1000 п.н., «Сибэнзим», Россия). Коллекция Всероссийского института генетических ресурсов растений им. Н.И. Вавилова.
химические признаки качества вида. Неравномерность распределения маркеров по хромосомам косвенно свидетельствует о присутствии на них установленных нами геномных блоков ко-адаптированных генов, а в целом по геному — на существование ко-адаптированных блоков генов. Наличие таких блоков в геноме B. rapa подтверждается нашими проведенными ранее исследованиями, в результате которых были установлены генетические детерминанты устойчивости B. rapa к разным расам Xanthomo-nas campestris pv. cam-pestris, вызывающим у растений этого вида сосудистый бактериоз (18-20). Следует отметить эффективность отобранных нами мо- лекулярных маркеров, поскольку некоторые маркеры (например, BRMS043 и BRMS034) оказались эффективными молекулярно-генетическими дескри- пторами вида по устойчивости к сосудистому бактериозу, которые мы применили для выявления коллекционных образцов B. rapa, устойчивых к каждой расе X. campestris отдельно и к патогену в целом (18, 21, 22).
При оценке 8 молекулярными маркерами в выборке из 18 образцов B. rapa мы выявили 26 полиморфных SSR-фрагментов размером от 122 до
410 п.н. Маркер BRMS051 (рис., А) при ожидаемом размере ампликона 262 п.н. у 77 % образцов был сцеплен с признаком длины и ширины листовой пластинки, причем у 58 % ее ширина составляла 20 см и более. В 60 % случаев аллели, представленные фрагментами 262 п.н. и 280 п.н., ока- зались ассоциированы. Фрагмент (аллель) 280 п.н. проявлялся у всех линий со средней длиной листовой пластинки (до 20 см), у всех образцов розеточной капусты, у всех образцов и сортотипов китайской капусты и у 60 % стабильных гибридов, родительской формой которых выступала китайская капуста. Отметим, что эти сортотипы китайской капусты имели зеленую (но не светло- или темно-зеленую) окраску листовой пластинки. Фрагмент (аллель) 262 п.н. обнаружили у 75 % образцов листовой репы.
Маркер KS51082 (см. рис., Б) был сцеплен с QTL длины черешка. Образцы к-115, к-163 и к-214 с ожидаемым фрагментом 282 п.н. имели длинный и очень длинный черешок (22-40 см). Ампликон 282 п.н. выявили у всех образцов японской капусты и у 50 % образцов листовой репы.
С помощью маркера BRMS043 (см. рис., В) обнаружили образцы с ожидаемым размером ампликона 318 п.н., у которых в 70 % случаев поверхность листовой пластинки была гладкой и слабоморщинистой. Все образцы с парой фрагментов (аллелей) 285 и 305 п.н. имели гладкие или слабоморщинистые листья. Образцы к-115, к-154, к-163 и к-213, сочетающие фрагменты 285 и 318 п.н., выделялись по содержанию β -каротина (5,16,2 мг/100 г). При наличии ампликона 318 п.н. 71 % вариантов имели достаточно длинную или длинную листовую пластинку (26-44 см), но средней ширины (16-23 см), а у 75 % образцов с короткими листовыми пластинками (11-23 см) проявлялся фрагмент (аллель) 305 п.н. У всех образцов китайской капусты и гибрида китайской и пекинской капусты нашли аллель 285 п.н., у японской капусты в 60 % случаев проявился фрагмент 318 п.н. У половины образцов листовой репы обнаружили тот же аллель (318 п.н.), у 75 % — аллель, соответствующий фрагменту 305 п.н.
Использование молекулярного маркера Ol12-F02, сцепленного с QTL опушения и окраски листовой пластинки (7), выявило четыре фрагмента — 200, 185, 175 и 140 п.н. Молекулярный анализ тех же образцов маркером BRMS014 при наличии одновременно двух ампликонов 263 и 280 п.н. установил его сцепленность с QTL окраски (от светло-зеленой до зеленой) для большей части образцов (80 %) из тех, у которых эти ампликоны проявились. Ту же пару ампликонов обнаружили у 70 % образцов китайской капусты и ее гибридов.
Скрининг маркером KS50200 (см. рис., Г) выявил фрагменты 292, 280 и 260 п.н. Образцы, проявившие ампликон 260 п.н., в 75 % случаев имели темно-зеленую листовую пластинку шириной 15-20 см с гладким, иногда слабоволнистым краем. Ампликон 280 п.н. обнаружили у гибридов китайской и пекинской капусты, а также у 75 % листовых реп (у последних ширина пластинки — в среднем 20 см, окраска темно-зеленая, край от гладкого до слабоволнистого). У китайской капусты (75 % образцов со светло-зеленой и зеленой окраской и гладким краем листовой пластинки при ее ширине 15-21 см) проявился фрагмент, соответствующий аллелю 292 п.н. Фрагмент этого размера обнаружили у 66 % образцов японской репы с острозубчатой надрезанностью края листа.
Скрининг молекулярным маркером BRMS034, сцепленным, как сообщалось ранее при анализе картирующих популяций линий удвоенных гаплоидов (12), с признаком окраски листовой пластинки, показал наличие ампликонов 122 и 144 п.н. (у форм с ампликоном 144 п.н. она темнозеленая), однако у изученных образцов не проявилась достоверная взаимосвязь между выявленными ампликонами и признаком.
При скрининге BRMS042 оказалось, что ампликон с ожидаемым размером 380 п.н. имелся у всех изученных образцов. У двух из трех образцов ноздреватой капусты обнаружили ампликон 410 п.н., который также выявили у одного образца китайской капусты (к-46). У японской капу-139
сты и листовой репы присутствовали ампликоны размером 620-170 п.н., нехарактерные для остальных образцов изученной стержневой коллекции.
Таким образом, мы провели комплексную (морфологическую, биохимическую, молекулярно-генетическую) оценку признаков качества у форм Brassica rapa в эколого-географических испытаниях при разных способах выращивания. Отметим, что анализ геномов B. rapa молекулярно-генетическими маркерами, находящимися в неравновесном сцеплении с QTL таких признаков, как характер окраски листовой пластинки, содержание общего белка, β -каротина, каротиноидов, аскорбиновой кислоты, сухого вещества, никогда ранее никем не выполнялся. Эти исследования подтвердили возможность использовать найденные нами SSR-маркеры морфологических и биохимических признаков для скрининга коллекционного и селекционного материала. Кроме того, полученные оригинальные результаты позволяют перейти к практическому осуществлению ассоциативного картирования и идентификации конкретных генетических детерминант, определяющих проявление ряда хозяйственно ценных и экономически значимых признаков качества, на основе использования уникального материала B. rapa из коллекции Всероссийского института генетических ресурсов растений им. Н.И. Вавилова (ВИР).
Список литературы Эколого-генетическая оценка морфологических и биохимических признаков качества у образцов коллекции Brassica rapa L. ВИР
- Соловьева А.Е., Артемьева А.М. Биохимические исследования восточноазиатских листовых овощных растений рода Brassica L. Труды по прикладной ботанике, генетике и селекции, 1999, 157: 142-148.
- Соловьева А.Е., Артемьева А.М. Капустные растения рода Brassica L. (Характеристика образцов по основным биохимическим показателям качества). В кн.: Каталог мировой коллекции ВИР. СПб, 2004. Вып. 756.
- Соловьева А.Е., Артемьева А.М. Биологически активные вещества капустных растений рода Brassica L. Аграрная Россия, 2006, 6: 52-56.
- Соловьева А.Е., Артемьева А.М. Качественная оценка некоторых восточноазиатских культурных типов вида Brassica rapa L. Аграрная Россия, 2006, 6: 56-60.
- Соловьева А.Е., Артемьева А.М. Особенности биохимического состава гибридов листовых овощных культур вида Brassica rapa L. Аграрная Россия, 2010, 3: 17-20.
- Артемьева А.М. Экологическая дифференциация капусты пекинской Brassica rapa ssp. pekinensis (Lour.) Olsson. В кн.: Генетические коллекции овощных растений. СПб, 2001. Т. 3: 148-166.
- Артемьева А.М. Доноры и источники для селекции листовых овощных культур вида Brassica rapa L. В кн.: Каталог мировой коллекции ВИР. СПб, 2004. Вып. 740.
- Lin K., Zhang N., Severing E.I., Nijveen H., Cheng F., Visser R.G.F., Wang X., de Ridder D., Bonnema G. Beyond genomic variation -comparison and functional annotation of three Brassica rapa genomes: a turnip, a rapid cycling and a Chinese cabbage. BMG Genomics, 2014, 15: 250 ( ) DOI: 10.1186/1471-2164-15-250
- Артемьева А.М., Соловьева А.Е., Чесноков Ю.В. Оценка морфологических и биохимических признаков качества у линий удвоенных гаплоидов Brassica rapa L. Вестник Российской академии сельскохозяйственных наук, 2014, 3: 38-41.
- Артемьева А.М., Соловьева А.Е., Кочерина Н.В., Чесноков Ю.В. QTL анализ биохимических признаков качества у Brassica rapa L. Овощи России, 2014, 1: 10-13.
- Артемьева А.М., Руднева Е.Н., Кочерина Н.В., Чесноков Ю.В. QTL анализ морфологических признаков качества у Brassica rapa L. Овощи России, 2014, 2: 14-17.
- Артемьева А.М., Соловьева А.Е., Кочерина Н.В., Беренсен Ф.А., Руднева Е.Н., Чесноков Ю.В. Картирование хромосомных локусов, определяющих проявление морфологических и биохимических признаков качества у культур вида Brassica rapa L. Физиология растений, 2016, 63(2): 275-289 ( ) DOI: 10.7868/S0015330316020044
- Specht C.E., Diederichsen A. Brassica. Mansfeld's Encyclopedia of agricultural and horticultural crops/P. Hanelt (ed.). Springer-Verlag, Berlin, 2001. V. 3: 1435-1465.
- Боос Г.В., Джохадзе Т.И., Артемьева А.М., Кривченко В.И., Симон А.М., Тимошенко З.В., Петровская Н.Н., Власова Э.А., Синельникова В.Н., Барашкова Э.А., Ивакин А.П., Ермаков А.И., Воскресенская В.В. Методические указания по изучению и поддержанию мировой коллекции капусты. Л., 1988.
- Ермаков А.И., Арасимович В.В., Иконникова М.И., Ярош Н.П., Луковникова Г.А. Методы биохимического исследования растений. Л., 1972.
- Дорохов Д.Б., Клоке Э. Быстрая и экономичная технология RAPD анализа растительных геномов. Генетика, 1997, 33: 358-365.
- Артемьева А.М., Чесноков Ю.В., Клоке Э. Генетическое разнообразие и внутривидовые филогенетические взаимоотношения культур вида Brassica rapa L. по результатам анализа микросателлитов. Информационный вестник ВОГиС, 2008, 12(4): 608-619.
- Артемьева А.М., Соловьева А.Е., Кочерина Н.В., Руднева Е.Н., Волкова А.И., Чесноков Ю.В. ДНК маркированные линии двойных гаплоидов Brassica rapa L. и идентифицированные QTL, контролирующие хозяйственно ценные признаки для использования в селекции листовых капустных культур. В кн.: Каталог мировой коллекции ВИР. СПб, 2012. Вып. 810.
- Артемьева А.М., Волкова А.И., Кочерина Н.В., Чесноков Ю.В. Молекулярно-генетическое картирование хромосомных локусов, определяющих устойчивость линий двойных гаплоидов Brassica rapa L. к сосудистому бактериозу. Известия Санкт-Петербургского государственного аграрного университета, 2012, 27: 73-77.
- Волкова А.И., Артемьева А.М., Чесноков Ю.В. Фитопатологическая оценка линий двойных гаплоидов вида Brassica rapa L. Мат. конф. молодых ученых и аспирантов «Актуальность наследия Н.И. Вавилова для развития биологических и сельскохозяйственных наук». СПб, 2012: 160-166.
- Волкова А.И., Артемьева А.М., Кочерина Н.В., Чесноков Ю.В. Идентификация молекулярных маркеров хромосомных локусов, детерминирующих устойчивость Brassica rapa L. к сосудистому бактериозу. Доклады ТСХА, 2013, 285(1): 219-222.
- Artemyeva A.M., Rudneva E.N., Volkova A.I., Kocherina N.V., Chesnokov Yu.V. Detection of chromosome loci determined morphological and black rot resistance traits in Brassica rapa L. Acta Horticulturae, 2013, 1005: 105-110 ( ) DOI: 10.17660/ActaHortic.2013.1005.8
