Эколого-географические особенности и перспективы использования Salicornia perennans в качестве масличной и энергонасыщенной культуры

Автор: Нестеров Виктор Николаевич, Розенцвет Ольга Анатольевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-2 т.19, 2017 года.
Бесплатный доступ
Исследовано влияние условий произрастания (эдафические, климатические, биотические факторы) на физиолого-биохимические характеристики растений Salicornia perennans, произрастающих в бассейне Средней и Нижней Волги. Установлено, что данный вид образует монодоминантные ценозы на солончаках, своеобразные пояса по берегам соленых озер, сменяющиеся в направлении градиента увлажнения и засоления. Растения имели суккулентный облик, который связан с оводненностью органов. Выявлено, что возрастание оводненности листьев растений (на 17%), связано с постепенным усилением аридности климата и достаточно сильно сказывается на процессах синтеза и распада биополимеров, таких как липиды и белки. Показана потенциальная возможность использования видов рода Salicornia (на примере S. perennans) в качестве масличной и/или энергонасыщенной культуры.
Засоленные почвы, аридизация климата, надземная и подземная биомасса, липиды, белки, абиотические факторы среды
Короткий адрес: https://sciup.org/148205367
IDR: 148205367 | УДК: 575.857+581.1
Ecological and geographical features and prospects of using of Salicornia perennans as oil and energy-sorted crops
The influence of growth conditions (soil, climatic, biotic factors) on physiological and biochemical characteristics of plants of Salicornia perennans, native to Middle Volga and the Lower Volga basin. It was found that this species forms monodominant cenoses on saline soils, the peculiar zone on the shores of the salt lakes, changing in the direction of the gradient of moisture and salinity. Plants were succulent appearance. It was found that the increase in water content of plant leaves (17%), due to the gradual strengthening of climate aridity and strong enough impact on the processes of synthesis and disintegration of biopolymers, such as proteins and lipids. It demonstrated the potential use of species of the genus Salicornia (on example of S. perennans) as oilseed and/or energy saturated culture.
Текст научной статьи Эколого-географические особенности и перспективы использования Salicornia perennans в качестве масличной и энергонасыщенной культуры
ослабленной конкуренцией, хотя и на непродолжительное время. Эти два признака – большое количество семян в годичной биомассе и способность произрастать на условно непригодных для культурных растений землях указывают на потенциальную возможность их использования в качестве с/х растений. Галофитное растениеводство, использующее для орошения соленые воды, может стать важным источником производства масличных и энергонасыщенных культур в аридных районах России и Средней Азии [4]. Известен целый ряд растений данного рода, семена которых содержат высокие концентрации полиненасыщенных жирных кислот [5, 6]. Так, один из видов Salicornia bigelovii дает 1,7 кг биомассы и 0,2 кг масличных семян на квадратный метр, что превышает урожайность сои и других масличных культур [4]. Следовательно, галофиты, приспособленные к жизнедеятельности в условиях засоления, с одной стороны являются потенциально перспективными культурными растениями, а с другой – видами, позволяющими вернуть в хозяйственный оборот нарушенные земли, снизить техногенное воздействие на окружающую среду, уменьшить засоление почвы [7, 8]. Истинные галофиты (виды родов Salicornia L., Suaeda Forssk. ex. и др.) даже нуждаются в засоленной почве, в то время как культурные гликофиты переносят лишь слабое засоление.
Известно, что солеустойчивость галофитов обусловлена необходимостью адаптации растений к осмотическому, токсическому и окислительному воздействию [9]. Замедленный обмен веществ галофитов в сравнении с гликофитами, позволяет растениям выживать в условиях засоления почвы, так как освобождает множество ресурсов (структурных блоков и энергии), необходимых для реализации защитной программы [10]. Экологическая оценка галофитов, особенности их физиолого-биохимических признаков при развитии на различных почвенных субстратах, отбор ценных кормовых, пищевых, лекарственных, масличных видов является актуальной задачей в условиях продолжающейся аридизации климата и антропогенной деградации земель [11]. Хотя галофиль-ных культурных растений нет, идет целенаправленный поиск использования нетрадиционных путей ведения с/хозяйства с участием галофитной растительности [5, 12]. Засоленные почвы на территории России составляют порядка 53 997 тыс. га (3,3% почвенного покрова страны или 5,0% площади равнинных территорий), а в южных регионах достигают 30–40% от площади с/хозяй-ственных угодий [13]. Приведенные факты свидетельствуют о необходимости последовательного изучения жизнедеятельности галофитов, особенно в естественной среде, выявления важнейших характеристик, обеспечивающих успешность произрастания на засоленных территориях и хозяйственно-полезные свойства.
Цель работы – исследовать влияние условий произрастания (эдафические, климатические, биотические факторы) на физиолого-биохимические характеристики Salicornia perennans . В задачу работы входило также показать потенциальную возможность использования видов рода Salicornia (на примере S. perennans ) в качестве масличной и/или энергонасыщенной культуры.
МАТЕРИАЛ И МЕТОДИКА
Объект исследования – Salicornia perennаns Willd. относится к сем. Chenopodiaceae. Растительный материал отбирали во второй декаде сентября 2013 и 2014 г., т.е. в конце вегетационного периода, когда однолетние растения полностью завершают свой рост и образуют семена. Станции отбора проб расположены на левобережной части Среднего и Нижнего Поволжья в Самарской, Саратовской, Волгорадской и Астраханской областях. Координаты станций отбора проб, нормы среднемесячных температур в период вегетации представлены в табл. 1.
В пределах одного фитоценоза размер закладываемой площадки для отбора проб растений и почвы составлял 10–15 м2. Следует уточнить, что у S. perennаns также как и у других видов рода редуцированные листья и стебель выпол- няют функцию листьев. Растения S. perennаns всегда прямостоячие, с членистыми, супротивными, голыми ветвями, осенью красного цвета и с семенами. Для биохимических анализов использовалась средняя часть таких «листьев» из 15–20 типичных растений, исключая самые мелкие и самые крупные экземпляры, произрастающие на площадке. Из объединенной биомассы листьев составляли три-пять независимых биологических проб (2–4 г сырой массы), которые потом и использовались для биохимических анализов. Экстракцию и анализ липидов в растительном материале проводили методами, описанными ранее [14]. Водорастворимый белок (ВБ) анализировали по методу Брэдфорд [15]. Суммарное содержание липидов рассчитывали как сумму проанализированных отдельно нейтральных липидов (НЛ), глико- (ГЛ) и фосфолипидов (ФЛ). Оводненность тканей рассчитывали после определения сырого и сухого веса как отношение содержания воды к сухому весу, в %. Для определения массы корней, стеблей и листьев отбирали 10–20 экземпляров растений типичных для выбранного фитоценоза.
Галофитные растительные сообщества исследовались в рамках системы классификации Браун–Бланке.
Агрохимический анализ почвы осуществлен в ФГБУ «Станция Агорохимической службы «Самарская», г. Самара.
Данные в таблицах и рисунках представлены как средние арифметические со стандартной ошибкой. Для выявления зависимостей использовали расчет коэффициента корреляции Спирмена. Все статистические расчеты выполнены с использованием программ Statistica 6.0 for Windows и Microsoft Excel 2007.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В литературе отмечено, что растительные сообщества с участием S. perennаns могут произрастать как на увлажненных береговых полосах различных соленых водоемов, так и на недавно освободившихся участках после отступления соленой воды, часто на периодически затапливаемых засоленных территориях [16]. Встречается S. perennans и в сильнозасоленных понижениях рельефа, но обязательно с высоким уровнем влажности почвы.
В наших исследованиях все выбранные экспериментальные площадки относятся к береговым участкам озер Б. Морец (№2), Булухта (№3), Эльтон (№4), Баскунчак (№5). Участок близ села Августовка (№1) представляет собой солончак, образовавшийся из-за близкого залегания грунтовых вод к поверхности.
Анализ почвы показал, что степень ее засоления высокая и составляет – 2–8 % от сухой
Таблица 1 . Место и координаты станций отбора проб растений и почвы, нормы средней месячной температуры за период вегетации (данные Гидрометцентра России)
|
№ станции |
Координаты |
Н°рмы средней месячн°й температуры, t ( ° C) |
|||||
|
Апрель |
Май |
Июнь |
Июль |
Август |
Сентябрь |
||
|
1 |
52 ° 15'с.ш„ 50 ° 42в.д. |
6,4 |
14,9 |
18,5 |
20,6 |
18,6 |
12,9 |
|
2 |
51 ° 28'с.ш., 50 ° 01'в.д. |
7,4 |
15,4 |
19,4 |
21,4 |
19,6 |
13,7 |
|
3 |
49 ° 22'с.ш., 46 ° 06в.д. |
9,6 |
17,0 |
21,1 |
23,5 |
22,0 |
16,1 |
|
4 |
49 ° 07' с.ш., 46 ° 50'в.д. |
9,6 |
17,0 |
21,1 |
23,5 |
22,0 |
16,1 |
|
5 |
48 ° 12'с.ш., 46 ° 49'в.д. |
11,1 |
18,3 |
22,8 |
25,2 |
23,3 |
17,4 |
Примечание: №1 – Село Августовка Самарской обл., солончаки, №2 – Оз. Большой Морец, рядом с п. Солянка Саратовской обл., №3 – Оз. Булухта Волгоградской обл., №4 – Оз. Эльтон, устье реки Б. Сморогда, Волгоградская обл., №5 – Оз. Баскунчак, рядом с с. Малый баскунчак, Астраханская обл.
массы (табл. 2) [13]. На станциях №3 и 5 уровень засоления был выше в сравнении с другими участками. Наиболее засоленные участки исследуемой территории, где произрастали растения, отличались и наибольшей увлажненностью почвы – 19–38 %. pH исследованных почв был 7,6–8,2 – щелочной. По содержанию основных макроэлементов выделялась станция №1, где было отмечено самое высокое содержание N, P и K в почве. Отношение С/N, характеризует обеспечение азотом гумуса. Эта величина составляла 11,6 и 13,6 на станциях №1 и №2, соответственно, несмотря на то, что на последней содержание азота было существенно ниже. На остальных участках отношение С/N было в два и более раз выше, что свидетельствует об обе-дненности почв азотом.
Ценотическое окружение исследуемых мест произрастания S. perennans варьировало, хотя практически повсеместно он является на засоленных и увлажненных участках почвы доминирующим видом. Средняя высота трав составляла 25–45 см. Проективное покрытие S. perennans составляло 15–65 %, при общем проективном покрытии трав 25–65% (табл. 3).
Наиболее крупные по содержанию сух. массы растения, включая корень, стебель и лист встречались на станции №4 (в районе оз. Эль-
Таблица 2 . Некоторые физико-химические характеристики почвы на станциях отбора проб
|
Почвенные характеристики |
Содержание некоторых макроэлементов |
||||||||
|
№ станции |
Влажность, % |
Соленость, % (сух. остаток) |
pH |
N, % |
C, % |
H, % |
C/N |
P, % |
K, % |
|
1 |
19,00 |
3,00 |
7,60 |
0,53 |
6,15 |
1,53 |
11,60 |
0,0089 |
0,07 |
|
2 |
4,40 |
1,60 |
8,20 |
0,07 |
0,95 |
0,17 |
13,57 |
0,0023 |
0,03 |
|
3 |
29,40 |
5,90 |
7,90 |
0,04 |
1,68 |
1,80 |
42,00 |
0,0005 |
0,02 |
|
4 |
23,80 |
1,60 |
8,00 |
0,05 |
1,28 |
0,40 |
25,60 |
0,003 |
0,03 |
|
5 |
37,90 |
7,90 |
8,10 |
0,08 |
2,47 |
1,57 |
30,87 |
0,0006 |
0,04 |
Таблица 3 . Характеристики сообществ в местах отбора проб. Проективное покрытие видов, %
|
№ станции |
1 |
2 |
3 |
4 |
5 |
|
Размер площадки, м2 |
15 |
15 |
12 |
10 |
10 |
|
Общ. проект. покр., % |
61 |
25 |
36 |
65 |
32 |
|
Средн. выс. трав, см |
45 |
25 |
35 |
45 |
35 |
|
Salicornia perennans |
55 |
20 |
35 |
65 |
15 |
|
Suaeda salsa |
5 |
1 |
1 |
||
|
Puccinellia distans |
+ |
3 |
|||
|
Limonium caspium |
3 |
||||
|
Tamarix ramosissima |
5 |
||||
|
Phragmites australis |
+ |
5 |
Примечание: Встречались в одном геоботаническом описании: 1 Artemisia paucifl ora , Suaeda acuminata (+); Limonium gmelinii, Petrosimonia triandra, Puccinellia gigantea (1); Halimione verrucifera (3)
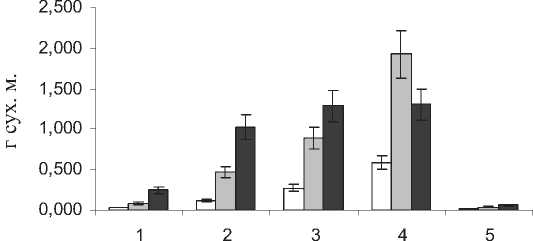
тон)– в среднем 3,8 г. сух. м., а наиболее мелкие – на ст. №1 и 5 – 0,2 и 0,4 г. сух м., соответственно (рис. 1).
Как показали наши результаты содержание влаги в исследуемых растениях различалось в зависимости от места произрастания и составляла от 14 до 90 % (рис. 2). Последнее значение, как правило, характерно для суккулентных эуга-
□ Корень □ Стебель □ Лист
№ станции отбора проб
Рис . 1 . Содержание сух. м. в органах S. perennans
□ Корень □ Стебель □ Лист
100 п
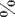
1 2 3 4 5
№ станции отбора проб
Рис . 2 . Оводненность органов S. perennans
лофитов, т.к. благодаря запасу жидкости происходит разбавление солей внутри растительного организма [17].
Наименее оводненные корни отмечены у растений станции №5 (14%), а наименее оводненные листья - у растений на станции №1 (68%).
Для того, чтобы проверить существует ли связь между семенной продуктивностью растений и их фитомассой были измерены 10 типичных растений на станции №4, где встречались наиболее крупные представители вида S. perennans . Масса растений варьировала от 10 до 75 г., а содержание семян - от 700 до 4600 шт на одно растение (рис. 3). Для растений с массой 10-30 г. количество семян составляло 700-1600 шт, а для растений с массой 45-75 г. - 1400-4600 шт. В целом установлено, что, чем крупнее было растение, тем большее количество семян оно содержало.
Далее была проанализирована фитомасса листьев с семенами с разных станций отбора проб на компонентный состав липидов и белков, которые характеризуют признаки маслич-ности и энергонасыщенности культуры.
Метаболизм азотсодержащих соединений, в первую очередь белков, играет важную роль в общем обмене веществ и в значительной мере определяет продуктивность растений. От уровня синтеза белка в клетке зависит интенсивность роста вегетативных органов [18]. Содержание водорастворимой фракции белка, которое обыч- но составляет более 50% от общего содержания белка, в надземной части S. perennans равнялось 0,5-0,7 мг/г сыр. м. в зависимости от места произрастания.
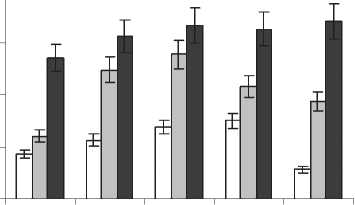
Анализ липидов надземной части растений показал, что содержание ГЛ, основных мембранных липидов хлоропластов, составило 0,41,0, ФЛ, которые составляют основу внешних мембран клеток и их органелл, - 0,5-1,8, НЛ, несущих в основном запасную и энергетическую функцию - 0,8-2,0 мг/г сыр. м. (рис. 4). Наибольшее количество структурных компонентов мембран - ГЛ и ФЛ отмечено на станции № 2, энергетических НЛ - на станции № 1.
В составе липидов было идентифицировано 18 компонентов: структурные мембранные липиды - Мгдг, Дгдг, Схдг, Фх, Фэ, Фг, Ст; запасные липиды - Таг, Даг, Вск, Эф, Уг, Сп; минорные компоненты и продукты метаболизма липидов - Фи, Фс, Дфг, Кис, Фк.
Анализ состава липидов показал высокое относительное содержание Дгдг (42-47%) по сравнению с Мгдг (28-34%), что вероятно объясняется тем, что растения пребывали в фазе плодоношения в конце вегетационного периода, так как в зеленой массе молодых растений, как правило, преобладает Мгдг (табл. 4). Состав ФЛ отличался высоким относительным содержанием обычно минорного Фи - 12-19%, однако наибольшее количество отмечено для Фх - 50-61% и Фэ - 14-21%.
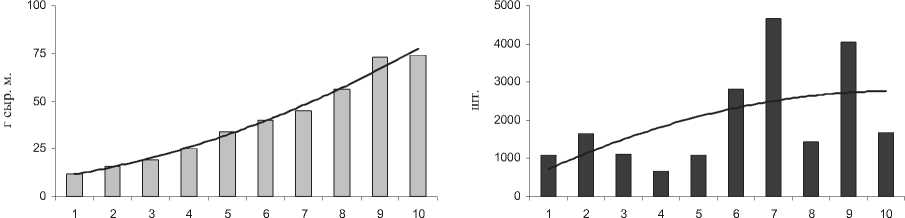
Рис. 3 . Масса 10 типичных растений S. perennans , г. сыр.м. (слева) и число их семян, шт. (справа).
Станция № 4
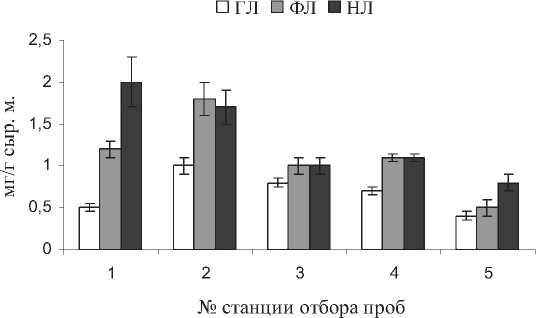
Рис. 4 . Содержание ГЛ, ФЛ, НЛ в листьях S. perennans
Таблица 4 . Состав липидов листьев S. perennans , % от суммы ГЛ, ФЛ и НЛ
|
Липиды |
№ станции отбора проб |
||||
|
1 |
2 |
3 |
4 |
5 |
|
|
Мгдг |
28,5 |
34,0 |
32,3 |
28,7 |
27,0 |
|
Дгдг |
46,5 |
45,5 |
42,5 |
46,8 |
46,0 |
|
Схдг |
25,0 |
20,5 |
25,2 |
24,5 |
27,0 |
|
Фх |
50,2 |
50,4 |
55,3 |
58,4 |
61,1 |
|
Фэ |
21,2 |
20,1 |
17,1 |
14,3 |
16,7 |
|
Фг |
9,9 |
13,3 |
12,5 |
13,2 |
9,1 |
|
Фк |
Сл. |
Сл. |
Сл. |
Сл. |
Сл. |
|
Фи |
18,7 |
16,1 |
15,2 |
12,1 |
11,6 |
|
Дфг |
Сл. |
0,1 |
Сл. |
Сл. |
Сл. |
|
Фс |
Сл. |
Сл. |
Сл. |
2,0 |
1,6 |
|
Ув |
4,9 |
14,7 |
13,3 |
10,4 |
20,4 |
|
Эф |
6,8 |
7,1 |
8,7 |
3,9 |
14,6 |
|
Вск |
0,4 |
1,1 |
2,2 |
1,4 |
1,8 |
|
Таг |
47,0 |
29,1 |
24,4 |
36,8 |
16,4 |
|
Неизвестные |
10,4 |
6,0 |
7,9 |
11,1 |
4,9 |
|
Кис |
10,4 |
10,5 |
8,6 |
5,9 |
9,7 |
|
Сп |
9,5 |
14,7 |
16,7 |
15,3 |
14,6 |
|
Ст |
7,8 |
11,3 |
13,9 |
10,2 |
12,3 |
|
Даг |
2,8 |
5,5 |
4,3 |
5,0 |
5,3 |
Примечание: среднее квадратичное отклонение составляет не более 10 %. Сл. - следовое количество
Одной из важнейших функций НЛ является запасание энергии. Эту роль выполняют Таг, которые служат формой хранения углерода и представляют собой важные вместилища питательных веществ. Эпидермис листьев, как правило, покрыт Вск, слой которых защищает их от потери влаги, механических и химических повреждений, патогенов и пр. Стерины способствуют стабилизации мембран, влияя на их текучесть и даже способны защищать зеленые части растений от перегрева. Было установлено, что содержание Таг и Вск взаимосвязано с содержанием в почве P (r=0,90, p =0,04 и r= -0,88, p =0,04), а содержание Ст взаимосвязано с отношением С/N в почве (r=0,87, p =0,04).
В составе ЖК были идентифицированы пальмитиновая (18-24%), стеариновая (2-3%), олеиновая (7-14%), линолевая (31-42%) и линоленовая (17-22%) кислоты (рис. 5). Как видно подавляющее количество ЖК составляют ненасыщенные кислоты, среди которых доминирует линолевая кислота. Как известно, именно данная ЖК содержится в семенах и маслах подсолнечника, льна, кукурузы, сои, облепихи и др. [19,20].
Исследование растений S. perennans, произрастающих на пяти независимых площадках, показало, что в целом данный вид образует монодоминантные ценозы на солончаках или своеобразные пояса по берегам соленых озер, сменяющиеся в направлении градиента увлаж-
□ С<16:0 □ С 16:0 ПС 18:0 ■ С 18:1 ■ С 18:2 И С 18:3 И С>20:0 В X
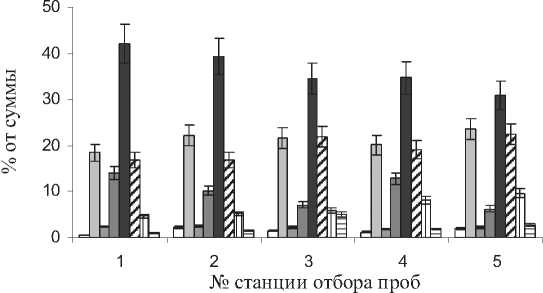
Рис . 5 . Состав ЖК в листьях S. perennans . Х - неизвестные ЖК
нения и засоления. Растения имели суккулент-ный облик, который непосредственно связан с оводненностью органов, в частности листьев,. Было установлено, что оводненность листьев связана с отношением главных макроэлементов в почве - С/N (r=0,90; p= 0,04), и, в основном, S. perennans произрастает на влажных почвах относительно бедных азотом. Следует подчеркнуть, что оводненность листа не была связана с накоплением ни сухой, ни сырой массы растений, но, что интересно, возрастала в направлении расположения выбранных станций от севера к югу - от 68 до 85%.
Как известно, наличие или недостаток овод-ненности органов достаточно сильно сказывается на биохимических параметрах, в частности на процессах синтеза и распада биополимеров, таких как липиды и белки, которые и составляют биохимическую основу масличных и энергонасыщенных культур. Очень тесно оводненность листьев коррелировала с относительным содержанием линоленовой кислоты (r=0,97, p =0,01) и длинноцепочечных ЖК (r=0,90, p =0,04), отношением ГЛ/ФЛ (r=0,97 p =0,04) в листьях. Однако оводненность имела отрицательную взаимосвязь с содержанием ФЛ (r= -0,89, p =0,04), НЛ (r= -0,88, p =0,04) и с относительным содержанием олеиновой ЖК (r= -0,90, p =0,04).
Установлено также, что на общее содержание ФЛ отрицательно влияло повышение концентрации в почве соли (r= -0,82 p=0,05), а на содержание НЛ - почвенной влаги (r= -0,89, p=0,04). Отношение ГЛ/ФЛ имело положительную зависимость с отношением С/N в почве (r=0,91 p=0,04). Следовательно, содержание воды в листьях растений оказывало существенное влияние на структурную организацию клетки, в частности на состояние мембранного аппарата, а также на содержание запасных компаундов.
Есть большая вероятность того, что возрастание оводненности (на 17%) связано с постепенным усилением аридности климата (табл. 1). Так, разница среднемесячных температур между станциями № 1-5 за период Апрель-Сентябрь составляла 3-5оС. Экологическое значение суккулентности галофитов заключается в запасании влаги, разбавлении солей внутри растительного организма [17], что с увеличением температуры становится более необходимым. Интересно отметить географическую закономерность с увеличением аридности климата (станции №1 ^ №5) не только в изменении оводненности, но также в содержании основных биохимических компонентов, играющих решающую роль в метаболизме растений и представляющих ресурсную ценность для человека (рис. 6). Так, содержания СЛ в надземной части S. perennans увеличивалось в 1,4 раза по мере увеличения среднемесячной температуры воздуха и роста оводненности листьев. В то же время содержание суммы ненасыщенных жирных кислот и относительный вклад линолевой ЖК, концентрация ВБ и НЛ снижались в 1,2-2,0 раза. В изменении мембранного аппарата отмечено возрастание доли липидов, ответственных за структуру непластидных мембран (увеличение отношения ГЛ/ФЛ в 2 раза) и липидов, поддерживающих бислойную структуру мембран (увеличение отношения ФХ/ФЭ в 1,5 раза).
Таким образом, S. perennans с учетом глобальных тенденций изменения климата является не только перспективной масляничной и энергонасыщенной культурой, но и в зависимости от географического положения и воздей-
Оводненность листьев
Содержание НЛ
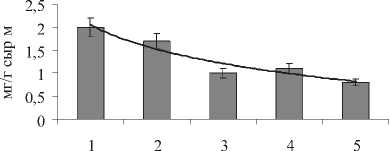
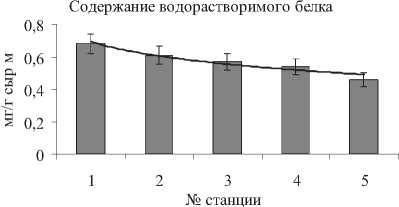
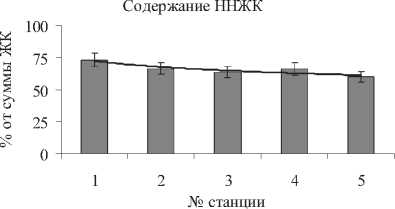
Рис . 6 . Закономерность изменения некоторых основных физиолого-биохимических характеристик надземной части S. perennans , связанная с ростом среднемесячных температур за вегетационный период и увеличением аридности климата (станции №1 ^ №5 или Север ^ Юг)
ствия локальных факторов среды, его ценопопу-ляции могут существенно (в 1,5–2 и более раз) отличаться по биомассе, содержанию липидов и соотношению их компонентов. Полученные данные могут стать основой для разработки технологий выращивания однолетних галофитов в качестве масляничных и энергонасыщенных культур в Южных регионах нашей страны.
Список литературы Эколого-географические особенности и перспективы использования Salicornia perennans в качестве масличной и энергонасыщенной культуры
- Global biodiversity information facility. Free and Open Access to Biodiversity Data. http://www.gbif.org (дата обращения14.09.2017).
- Раменский Л.Г. Избранные работы. Проблемы и методы изучения растительного покрова. Л.: Наука, 1971. 334 с.
- Grime J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory//Am. Nat. 1977. V. 111. P. 1169-1194.
- Шамсутдинова Э.З., Старшинова О.А., Шамсутдинов З.Ш. Галофитное растениеводство: концепция, опыт, перспективы//Достижения науки и техники АПК. 2013. № 11. С. 36-39.
- Hameed A., Khan M.A. Halophytes: Biology and Economic Potentials//Karachi University Journal of Science. 2011.V. 39. P. 40-44.
- Qasim M., Gulzar S., Shinwari Z.K., Khan M.A. Traditional ethno-botanical uses of halophytes from Hub, Balochistan//Pak. J. Bot. 2010. V. 42. P. 1543-1551.
- Чибрик Т.С., Глазырина М.А. Биологическая рекультивация и мониторинг нарушенных промышленностью земель: Учеб. пособие. Екатеринбург: УрГУ, 2008. 193 с.
- Реджепбаев К. Опыт выращивания галофитов на засоленных землях. Ашхабад, 2009. 44 с.
- Lokhande V.H., Suprasanna P. Prospects of Halophytes in Understanding and Managing Abiotic Stress Tolerance. In: P. Ahmad and M.N.V. Prasad (eds.), Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change. 2012. Springer Science+Business Media, LLC. P. 29-57.
- Шихмурадов А.З. Биоресурсный потенциал и эколого-генетические аспекты устойчивости представителей рода Triticum L. к солевому стрессу. Дисс. д.б.н. Дербент, 2014. 275 с.
- Оценочный доклад об изменении климата… Том. 1 Изменение климата. М.: Росгидромет, 2008. 227 с.
- Шамсутдинов З.Ш., Савченко И.В., Шамсутдинов Н.З. Галофиты России, их экологическая оценка и использование. М.: ООО Эдель-М, 2001. 399 с.
- Шишов Л.Л., Панкова Е.И. Засоленные почвы России. М.: ИКЦ «Академкнига», 2006. 854 с.
- Rozentsvet O.A., Nesterov V.N., Bogdanova E.S. Membrane-forming lipids of wild halophytes growing under the conditions of Prieltonie of South Russia//Phytochemistry. 2014. V. 105. P. 37-42.
- Bradford M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding//Anal. Biochem. 1976.V. 72. P. 248-254.
- Рухленко И.А. Разнообразие высших синтаксонов внутриконтинентальной растительности сильно засоленных почв юга России и сопредельных территорий в рамках устойчивого развития и сохранения биоразнообразия//Поволжский экологический журнал. 2014. № 1. С. 31-37.
- Ogburn R.M., Edwards E.J. The Ecological Water-Use Strategies of Succulent Plants//Advances in Botanical Research/Eds. Kader J.-C., Delseny M. Netherlands: Elsevier, 2010. P. 180-215.
- Орлова Н.В., Кусакина М.Г., Сучкова Н.В. Зависимость содержания водорастворимых белков в органах галофитов от уровня засоления почвы//Вестн. Перм. ун-та. 2007. Вып. 5 (10). С. 31-34.
- Хасанов В.В., Рыжова Г.Л., Дычко К.А., Куряева Т.Т. Состав жирных кислот и стероидов растительных масел//Химия растительного сырья. 2006. № 3. С. 27-31.
- Кучеренко Л.А., Петибская В.С., Ефименко С.Г., Савельев А.А. Содержание и качество масла в семенах сортообразцов сои различного происхождения//Масличные культуры. 2008. Вып. 1. (138). С. 1-6.
