Эколого-гомологические ряды разных масштабов

Автор: Розенберг Г.С.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общие проблемы экологии
Статья в выпуске: 2 т.2, 2000 года.
Бесплатный доступ
Рассмотрены различные варианты параллельной изменчивости (гомологичности) экологических объектов разной природы. Показана принципиальная возможность использования закона гомологических рядов на уровне популяций, сообществ и теоретических конструкций современной экологии.
Короткий адрес: https://sciup.org/148197588
IDR: 148197588
Ecology-homologous series of different scales
The various variants of parallel variability (homologous) of ecological objects of a different nature are considered. The basic opportunity of use of the low of homologous series at a level populations, communities and theoretical designs of modern ecology is shown.
Текст научной статьи Эколого-гомологические ряды разных масштабов
Институт экологии Волжского бассейна РАН, г. Тольятти
Рассмотрены различные варианты параллельной изменчивости (гомологичности) экологических объектов разной природы. Показана принципиальная возможность использования закона гомологических рядов на уровне популяций, сообществ и теоретических конструкций современной экологии.
Понятие "гомология" (сходство, единство или близость объектов) достаточно широко используется в естествознании - неорганической химии (химические элементы главной подгруппы каждого вертикального столбца Периодической системы Д.И. Менделеева), органической химии (вещества, сходные по химическим свойствам - системы предельных и непредельных углеводородов), сравнительной анатомии (органы, имеющие общее происхождение), при изучении изменчивости признаков культурных растений (закон гомологических рядов наследственной изменчивости Н.И. Вавилова), в молекулярной биологии и генетике - гомологии на уровне триплетов (состоят из четырех нуклеотидов), генов (семейства генов, несущих информацию о структуре родственных белков, имеют очень высокое сходство последовательностей триплетов), хромосом (одинаковый набор и порядок генов) и генома (по геномному критерию родом считается группа близкородственных видов, имеющих специфический первичный геном или полиплоидный геном, состоящий из двух или более копий этого специфического первичного генома).
Гомологические ряды видов
Примеров такого рода рядов имеется предостаточно. Сошлемся лишь на классическую работу Н.И. Вавилова [1, с.49-50]: "Н.В. Мо-розова-Водяницкая в статье "Гомологические ряды как основа классификации рода Pediastrum Meyen" (1925) подтвердила на водорослях применимость закона гомологических рядов и сделала попытку применения его к систематике рода Pediastrum... В.М. Шимкевич (1906) для пантопод дал периодическую систему классификации, которая облегчила ориентировку в разнообразии форм и дала возможность предсказать существование еще неизвестных форм. Д.Н. Соболев (1913) установил параллельные ряды у ископаемых головоногих (гониотитов), которых он назвал в своей более поздней работе (1924) "изоморфными", так как в основе этих параллелизмов лежит изоморфия живого вещества, т.е. сходство в строении этого вещества. В.А. Догель (1923), классифицируя инфузорий, расположил их в параллельные ряды. Г.Г. Виттенберг, применив закон гомологических рядов, построил стройную периодическую систему трематод (Cyclocoelidae) П.В. Терентьев нашел параллельные ряды у амфибий, Добжанский у божьих коровок (Coccinellidae), Б.Н. Шванвич - у Rhopalocera".
"Обобщая сказанное, можно назвать следующие общие свойства объектов, формирующих гомологические ряды. Члены одного гомологического ряда:
-
- состоят из более простых однотипных элементов;
-
- имеют общий план строения;
-
- различаются между собой по составу, количеству или взаимному расположению этих элементов" [2, с.427].
Гомологические ряды популяций
Интересный вариант периодической системы группового поведения был предложен в 1978 г. Ю.К.Рощевским (цит. по [3]), который представлен на следующей схеме:
Таблица1. Вариант периодической системы группового поведения популяций
|
Уровень |
Конструктивный уровень группы |
||||
|
индивидуальности |
Ð |
À |
R |
O |
I |
|
рассудочный ( r ) |
Pr |
Ar |
Rr |
Or |
|
|
самообучаеымй ( d ) |
Pd |
Ad |
Rd |
Od |
|
|
инстинктивный ( i ) |
Pi |
Ai |
Ri |
Oi |
|
|
кинезный ( k ) |
Pk |
Ak |
Rk |
Ok |
Ik |
|
нулевой ( 0 ) |
Io |
||||
Ю.К.Рощевский различает пять уровней поведенческой индивидуальности:
-
• нулевой (0) - усложнение химической организации, приведшее к возникновению на Земле простейших организмов;
-
• кинезный (k) - конструкция моновидовых групп основана на врожденных внутриклеточных механизмах; это самый простой уровень поведенческой индивидуальности;
-
• инстинктивный (i) - безусловно рефлекторное поведение особей в группах с использованием сигнальных индивидуальных приспособлений; поведение таких животных "приводится" в соответствие с изменяющимися факторами среды путем естественного отбора;
-
• самообучаемый (d) - связан с высшей формой сигнальных приспособлений; животные самообучающегося уровня обладают перцептивной психикой;
-
• рассудочный (r) - высший уровень поведенческой индивидуальности; этого уровня достиг только человек.
Обратим внимание на то, что эти пять уровней поведенческой индивидуальности совпадают с пятью принципами усложняющегося поведения систем [3,4]. В конструкции моновидовых групп Ю.К. Рощевский также различает пять уровней по степени усиления целостности:
-
• протоинтегративный (Р, от греч. protos - первый) - целостность системы определяется только близостью составляющих ее особей (механическая целостность); пример -скопление муравьев-фуражиров возле только что налитого сиропа;
-
• эквипотенциальный (А, от лат. aequus -равный) - целостность определяется поведенческим группированием, все особи такой системы равноценны (миграционные стаи
саранчи, шеренги муравьев-воинов, летящий за маткой рой пчел - примеры Ai, прохождение строем солдат - пример Ar; Ad - подражание в прыжках в воду - "волна бегства" -прудовой лягушки Rana esculenta);
-
• возвратно-дифференциальный (R, от лат. refero - нести назад) - группировка особей осуществляется по социальным функциям; поведение разных функциональных групп различно и взаимозаменяемо (Ri - поведение муравьев в муравейнике, Rd - пожалуй, самый совершенный способ группового поведения животных);
-
• облигатно-дифференцированный (О, от лат. obligatus - обязательный) - каждая социальная общность индивидуумов выполняет только один комплекс поведенческих реакций, определяющих конструкцию группы, и не может выполнять никакой другой (вариант Od - семья, Oi - полиморфизм пчел, муравьев, термитов);
-
• организменный (I, от лат. individuus - неделимый) - элементы системы (особи) перестают функционировать как самостоятельные организмы, "...высокая интегрированность системы входит в такое сильное противоречие с индивидуальностью ее элементов, что полностью блокирует всякую возможность проявления у них каких либо поведенческих реакций" (цит. по [3, с.201]).
Гомологические ряды сообществ
Гомологические ряды сообществ [2, 5] -синтаксоны как близких, так и отдаленных типов растительных сообществ, характеризуются параллельными рядами изменчивости флористического состава.
"Свойства, присущие объектам, составляющим различные типы гомологических рядов, прослеживаются также и в раститель- ности и отражающих ее разнообразие синтаксонах (единица систематики растительных сообществ. - Ремарка наша). В синтаксоно-мии под общим планом строения мы будем понимать участие во флористическом составе близких синтаксонов одних и тех же групп диагностических видов, индицирующих определенный набор факторов среды. Группы диагностических видов в данном случае выступают в качестве однотипных, повторяющихся в разных синтаксонах и более простых по отношению ко всему флористическому составу синтаксонов элементов. Если в двух или более синтаксонах имеется не одна, а несколько повторяющихся групп видов, то изменчивость таких синтаксонов может быть представлена в виде рядов с параллельно меняющимся флористическим составом, которые мы будем называть гомологическими рядами изменчивости растительных сообществ" [2, с.427].
В качестве примера (табл.2) рассмотрим гомологические ряды изменчивости у трех ассоциаций сфагновых болот (две первые -из Западной Европы, последняя - из северозападных районов России; [2, с.429-430]) класса Scheuchzerio-Caricetea, порядка Sheuchzerietalia Nordh. 1936, союзов Rhynchosporion albae Koch 1926 и Caricion lasiocarpae Van. Bergh. in Lebr. et al. 1949. Каж дая из трех ассоциаций имеет сходный набор субассоциаций, выделяемых по доминированию сфагновых мхов и образующих параллельные ряды изменчивости.
Представленный результат достаточно нагляден, и здесь прокомментируем только одно свойство гомологических рядов - прогностическую роль, т.е. способность предсказывать существование новых, еще не описанных типов растительных сообществ. Выявив в классификационной системе в одной из ассоциаций полный ряд изменчивости, следует ожидать, что и другие близкие к ней ассоциации будут иметь такие же ряды. Так, в сообществах олигомезотрофных болот (см. табл.2) ассоциации Caricetum limosae, описанных в Центральной и Северо-Западной Европе, отсутствуют субассоциации (в таблице они отмечены пунктирным овалом) sphagnetosum maji и shp. baltici , а в ассоциации Sphagno-Rhynchosporetum - субассоциация sph. cuspidati. "Возникает вопрос: связано отсутствие этих субассоциаций с какими-либо экологическими или фитосоциологичес-кими ограничениями, или объясняется неполнотой данных? Поиск сообществ для заполнения пустующих мест в данных рядах показал, что такие синтаксоны были описаны на северо-западе России... и на Южном Урале. Основанием для прогноза служит
Таблица 2. Гомологические ряды изменчивости у трех ассоциаций сфагновых болот
Ассоциация Число описаний Номер синтаксона
Диагностические виды
ассоциаций и субассоциаций
Rhynchospora alba Carex limosa
Carex lasiocarpa Sphagnum pulchrum Sphagnum lindbergii Sphagnum fallax Sphagnum majus Sphagnum cuspidatum Sphagnum balticum Sphagnum papillosum Sphagnum subsecundum Sphagnum angustifolium
Sphagno-Rhynchosporetum
11 110 76 20 92
2 3 4 5 6
66 8
Caricetum limosae
Caricetum lasiocarpae
VV V VVVV
I IV I
I I
V
V
V
I
I
I
II
II
153 9
146 10
338 11
110 12
II I II II II II
V V V IV V V
I III
I III
I
I
I III III
I I II
I
III I
I
I II
I I
II
I V
I
I
I V
II V I I
I
II I
I
I V
I III
I II I
I
II
I
I
V
II I I
I
I
I
II
II
I I II I
VVVVVV
I
II
V
II
V I
III
II
I V
V
I
V
V
III
V
III
I
V
V
Примечание. Субассоциации: колонки 1,8 - sphagnetosum pulchri; 2, 9 - sph. lindbergii, 3, 10 - sph. fallacis; 4, 15 - sph. maji; 5, 17 - sph. baltici; 6, 12, 18 - sph. papillosi; 7, 13 - sph. subsecundi; 11, 16 - sph. cuspidati; 19 - sph. angustifolii. Римские цифры - постоянство видов в синтаксонах: I -1-20%, II - 21-40%, III -41-60%, IV- 61-80% и V- 81-100%. Штриховка - общее проективное покрытие: без штриховки - до 25%, далее - 26-50%, 51-75% и более 75%.
предположение, что близкие синтаксоны подчинены одному закону преобразования" [2, с.434].
Заметим, что в синтаксономии с конца 80-х годов [6, с. 158] используется понятие "рефрен" (от фр. refrain - припев) - повторяющиеся синтаксоны-аналоги в параллельных экологических рядах (например, ряды по отношению к фактору засоления при разных режимах увлажнения). В этом контексте рефрены являются полным аналогом гомологических рядов в понимании А.И. Соломеща.
Зонально-ландшафтные гомологические ряды
Основными факторами географической зональности являются солнечная радиация и показатели количества влаги. Именно на этих параметрах основаны многочисленные индексы, оценивающие различия и сопоставляющие величины тепла и влаги в разных территориях. В настоящее время наибольшей популярностью пользуется "радиационный индекс сухости" М.И. Будыко, предложенный в 1948 г. и имеющий, по мнению многих географов, наиболее общий биогеографический смысл:
КВ = R / (L * г) , где R - годовой радиационный баланс; L -скрытая теплота испарения; r - годовая сумма осадков. Если КВ = 1, то это свидетельствует о соразмерности между количеством тепла и влаги, что характеризует наиболее благоприятные для биологических компонентов ландшафта условия. Значения КВ < 1 указывают на избыток влаги, КВ > 1 - на повышенную сухость.
Существует еще целый ряд гидротермических индексов, используемых при количественном анализе отдельных компонентов биосферы. Укажем только на один показатель, который использовал Д.А. Криволуцкий для анализа зонального распределения панцирных клещей, - " показатель благоприятствования условий " :
Кк
= R
( F + L) r
F
Е
m
где F - годовое количество опада, L - вес подстилки, Е - среднегодовая испаряемость.
Концепция периодической географичес кой зональности Григорьева-Будыко [7] - со сменой физико-географических поясов аналогичные ландшафтные зоны и их некоторые общие свойства периодически повторяются. В частности, наблюдается повторение индекса Будыко (КВ). Как подчеркивает Ю.И. Чернов [8, c.16], "...в разных климатических поясах наблюдается повторение радиационного индекса сухости. Это позволило А.А. Григорьеву и М.И. Будыко высказать идею "периодического закона географической зональности": в разных климатических поясах имеются зоны-аналоги с близкими величинами радиационного индекса сухости, сходные по структуре сообществ. Величина индекса около единицы соответствует как зоне широколиственных лесов в умеренном поясе, так и определенным территориям в субтропическом и экваториальном. Тундрам в тропических широтах соответствуют территории с резким преобладанием болот (индекс в пределах 0,2-0,4)".
Хорошей иллюстрацией этой концепции являются данные Д.А.Криволуцкого по соотношению численности панцирных клещей и индекса КК(цит. по: [8, с.157]), представленные на рис.1.
Гомологические ряды общеэкологического плана
Фактически, такого рода рядами пред-
Средняя численность
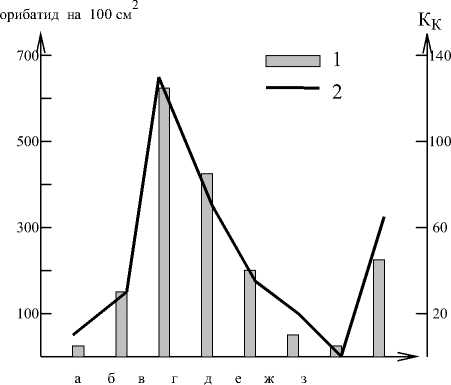
Рис. 1. Соотношение численности панцирных клещей (1) и "показателя благоприятствования условий" (2) в разных географических зонах: а - арктическая пустыня, б - тундра, в - тайга, г -широколиственный лес, д - луговая степь, е - сухая степь, ж - пустыня, з - субтропический лес [8]
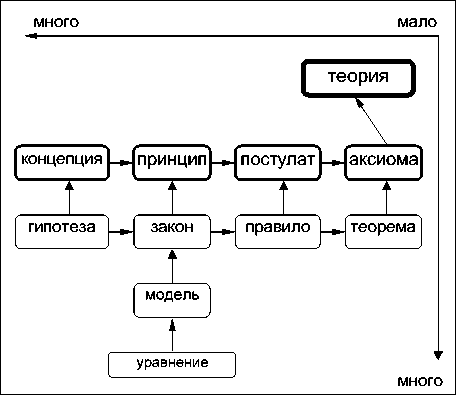
Рис. 2. Схема "соподчинения"основных теоретических конструкций (пояснения в тексте)
ставляются схемы "соподчинения" основных понятий (см. рис.2), которые призваны описать "ядро экологической теории" [3,9,10] или "центральное понятийное звено" [7, с.8].
Горизонтальные связи на этой схеме указывают направление возрастания "истинно сти" тех или иных положений теории, вертикальные - возрастание "важности", "главенства этих положений". Координатные оси указывают количественное соотношение различных понятий (очевидно, что частных уравнений будет значительно больше, чем основополагающих принципов, а гипотез -больше, чем теорем). Все теоретические конструкции современной экологии [7, 11] составляют фундамент "теоретической экологии" и объединены в 12 основных концепций современной экологии [3,12], которые без подробного комментария представлены в табл.2. Естественно, что эти концепции не охватывают все многообразие теоретических конструкций - особенно это касается разделов математической экологии, где число построенных и проанализированных моделей и разного рода уравнений огромно. С другой стороны, предложенные схемы (система концепций) помогают определить "слабые места" в строящемся здании теоретической экологии, что и позволяет рассматривать их в качестве своеобразных гомологических рядов.
Таблица 3. Концепции, принципы и другие теоретические конструкции современной экологии
|
Подраздел экологии |
Концепция |
Гипотеза |
Принцип |
Закон |
Постулат |
Правило |
Аксиома |
|
Факториальная экология |
Совокупного действия природных факторов |
3 |
1 |
3 |
1 |
3 |
|
|
Взаимодействие популяций |
Естественного отбора |
3 |
3 |
3 |
2 |
||
|
Демэкология (структура) |
Минимального размера популяции |
7 |
1 |
2 |
|||
|
Демэкология (динамика) |
Устойчивости популяций |
3 |
1 |
1 |
1 |
||
|
Экологические ниши |
Экологической ниши |
1 |
1 |
1 |
3 |
||
|
Экологическое Разнообразие |
Экологического разнообразия |
2 |
2 |
2 |
1 |
5 |
|
|
Экология сообществ (синэкология) |
Экосистемы |
3 |
2 |
1 |
2 |
||
|
Структура сообществ (общие закономерности) |
Континуума |
3 |
4 |
2 |
1 |
||
|
Структура сообществ (частные, пространственные закономерности) |
Периодической географической зональности |
2 |
1 |
2 |
|||
|
Динамика сообществ (сукцессии, климакс) |
Климакса |
4 |
1 |
3 |
1 |
1 |
|
|
Динамика сообществ (эволюция) |
Сетчатой эволюции сообществ |
3 |
1 |
3 |
1 |
Нетрудно видеть (табл.3), что число аксиом и теорем крайне мало (в этом просматривается прогностическая сила данной эколого-гомологический системы концепций современной экологии - можно видеть "слабые места" теоретических построений в экологии и, соответственно, направлять усилия на их преодоление). С другой стороны, представление всех известных теоретических конструкций по единой схеме способствует синтезу экологической теории.
Закончить статью хочется цитатой Н.И. Вавилова [1, c.54-55]: "Законгомологических рядов не есть прокрустово ложе, ограничивающее изменчивость; наоборот, он вскрывает и вскрыл практически огромные возможности изменчивости, констатируя лишь, что в целом, при сопоставлении выполненных систем, путем исчерпывающего изучения всех звеньев, составляющих вид (как показано, - и не только. - Ремарка наша), ряды изменчивости, характерные для видов, проявляют не беспорядочный процесс, а определенные правильности, вытекающие по существу из эволюционного развития".
