Экспериментальная оценка температурного оптимума у молоди пресноводных видов рыб методом термопреферендума

Автор: Голованов Владимир Константинович
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Экология
Статья в выпуске: 3-4, 2013 года.
Бесплатный доступ
В статье описывается метод термопрефендума, позволяющий определять значение температурного оптимума у молоди рыб; анализируются данные по 21 виду пресноводных рыб - значения окончательно избираемых температур у сеголеток и годовиков -в летний сезон года.
Молодь рыб, термоизбирание, избираемая температура, окончательно избираемая температура, метод, эксперимент, термоградиентные условия
Короткий адрес: https://sciup.org/14720042
IDR: 14720042 | УДК: 57.088.1:591.128.3:597
Experimental estimation of а temperature optimum in the juvenile of a freshwater fish species by method thermopreferendum
The description of a method allowing to define meaning of a temperature optimum for the juvenile fish is given. The thermopreferendum method is based on creating of experimental thermogradient conditions to fishes, in which they spontaneously choose a zone of a ecological-physiological optimum. The data on 21 species of freshwater fishes -meaning of final preferred temperatures for fingerlings and yearlings in a summer season of year are analyzed. The received results are urgent and are claimed both in theoretical and in the practical plan.
Текст научной статьи Экспериментальная оценка температурного оптимума у молоди пресноводных видов рыб методом термопреферендума
Поведение является важной формой жизнедеятельности животных, обеспечивающей им возможность быстро реагировать на изменения среды. В процессе прохождения сезонных и жизненных циклов гидробионты постоянно сталкиваются с градиентом абиотических и биотических факторов водной среды. Сезонные термоклины в глубоководных водоемах, весенний прогрев и осеннее охлаждение воды, аномальное повышение температуры в летний сезон года, сброс подогретых вод ГРЭС, АЭС и крупных про- мышленных предприятий – только краткий перечень существующих термоградиентных условий. В естественных или экспериментальных условиях при наличии градиента температуры рыбы обычно не остаются в тех условиях, в которых они обитали или содержались в лаборатории предварительно. Наблюдения показали, что особи разных видов рыб проявляют в таком случае реакцию, которую зарубежные исследователи назвали «thermopreferendum» [17; 18; 24], а отечественные – «избиранием температуры» [7].
Исследования проводились в 1974– 2012 гг. на 21 виде пресноводных рыб, относящихся к 9 семействам: Acipenseridae (сибирский осетр Acipenser baerii ), Salmonidae (радужная форель Parasalmo ( Oncorhynchus ) mykiss ), Coregonidae (пелядь Coregonus peled ), Esocidae (обыкновенная щука Esox lucius ), Cyprinidae (синец Abramis ballerus ; лещ Abramis brama ; уклейка Alburnus alburnus ; густера Blicca bjo-erkna ; серебряный карась Carassius auratus gibelio ; золотой, или обыкновенный, карась Carassius carassius , сазан или обыкновенный карп Cyprinus carpio , пескарь Gobio gobio ; язь Leuciscus idus ; обыкновенный гольян Phoxinus phoxinus ; плотва Rutilus rutilus ), Cobitidae (вьюн Misgurnus fossilis ), Lotidae (налим Lota lota ), Percidae (обыкновенный ерш Gymnocephalus cernuus ; речной окунь Perca fluviatilis ; обыкновенный судак Sander lucioperca ), Odontobutidae (головеш-ка-ротан Perccottus glenii ).
Большинство исследованных видов широко распространено в водоемах Северо-Запада России и отличается по образу жизни и характеру питания. Рыб отлавливали в Рыбинском водохранилище, его притоках и близлежащих водоемах или выращивали на экспериментальной прудовой базе «Суно-га» ИБВВ РАН. Отлов проводили мальковыми волокушами (25–50 м).
После отлова в течение 0,5–2,0 ч рыб доставляли в лабораторию и помещали в аквариумы объемом от 60 до 300 л с от-9
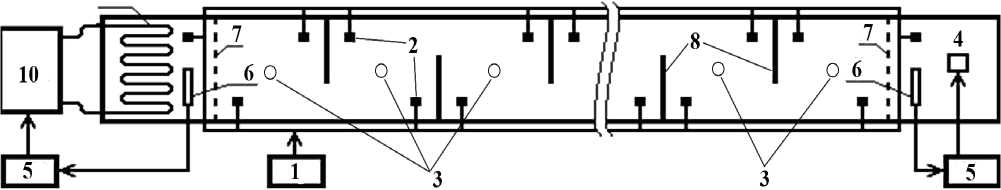
1 – компрессор, 2 – распылитель, 3 – термометр (электронный датчик), 4 – нагреватель, 5 – терморегулятор, 6 – датчик температуры, 7 – сетка-ограждение, 8 – неполные перегородки между отсеками, 9 – теплообменник, 10 – холодильный агрегат
стоянной водопроводной водой, а также регулируемой температурой и аэрацией. Всех рыб акклимировали в течение 7–14 дней к температуре, близкой к средним значениям летнего сезона (18–22 °С) и содержали в условиях естественного фотопериода при периодической смене воды. В период акклимации и во время опытов рыб кормили живым кормом (дафния, зоопланктон, олигохеты, личинки хи-рономид), рыбным фаршем, сухим кормом (дафния, рыбный комбикорм) или молодью рыб (щука).
Динамика выбора температур в градиентных условиях у разных видов рыб отличается и характеризуется в течение первых часов наблюдения разнонаправленностью. Температура, которую рыбы выбирают в течение часов или нескольких суток после предоставления термоградиентных условий, принято называть избираемыми температурами (ИТ) [3; 4; 17–19; 24].
При определении ИТ и окончательно избираемой температуры (ОИТ) использовался метод «конечного термопреферендума» [7; 11; 17; 18; 21], при котором рыбам предоставлялась возможность свободного выбора температуры в условиях термоградиента.
Относительно простая техника создания температурного градиента в горизонтальных емкостях описана в научной литературе [11]. Общая схема экспериментальной установки для определения ИТ и ОИТ приведена на рисунке.
Рисунок. Схема экспериментальной установки для определения избираемой и окончательно избираемой температуры у молоди рыб
Градиент создавали путем поддержания контрастных значений температуры на противоположных концах лотка с помощью устройств автоматического подогрева и ох- лаждения воды. Низкую температуру в холодном отсеке – с помощью холодильных агрегатов ВСЭ-1.5 и Aspera UJ9232E, высокую температуру в теплом отсеке – с по- мощью нагревательного элемента. Диапазон температуры, создаваемый в градиентной установке, составлял 15–20 °С.
Для получения равномерного горизонтального градиента температуры, уменьшения конвекционных токов и устранения вертикального градиента лотки были разделены на 10–12 отсеков неполными перегородками, образующими зигзагообразный лабиринт. Для предотвращения вертикального градиента температуры в каждом отсеке устанавливали по 2 распылителя с подачей воздуха от мощного компрессора. За счет донного расположения распылителей происходило вертикальное перемешивание воды. Это позволяло создавать плавный горизонтальный градиент температуры, составлявший 0,04– 0,13 °C/см. Общая длина всех рабочих камер в установках различного типа (для личинок, сеголетков и годовиков, неполовозрелых и взрослых особей) составляла 2,4 и 4,2 м. Температуру измеряли с помощью ртутных термометров, электротермометров, а также дистанционных датчиков температуры, расположенных в каждой камере установки. Для регистрации распределения рыб также использовали видеокамеры Panasonic SDR-H250 и Canon Legria FS306 с непрерывным или периодическим включением. Продолжительность опытов составляла 10–14 суток.
Разработанные нами установки предназначены в основном для работы с непроточной водой, однако в них предусмотрена и система эрлифт-самоочистки. Данная система создавала небольшой проток воды 1–2 л/ч и периодически включалась на 1–2 суток во время опыта либо в промежутках между опытами. Все установки размещались либо в отдельном помещении, либо в аквариальной части лаборатории, где ограждались матерчатыми загородками для предотвращения влияния различных внешних раздражителей.
Группу рыб (10 экз.) помещали в отсек установки с температурой воды, равной температуре акклимации. Распределение рыб, а также избираемая ими температура на начальном этапе выбора фиксировали 8–10 раз в светлое время суток с интервалом в 1,0–1,5 ч. За величину избираемой температуры принимали температуру в отсеке, в котором находилась каждая особь в момент снятия показаний. Данные за каждые сутки опыта суммировали и делили на число наблюдений (для 10 рыб число наблюдений за сутки составляло от 80 до 100), получая средние значения ИТ. Если в течение трех суток средние значения ИТ достоверно не различались, эту температуру принимали за значение ОИТ, характеризующее зону стабильного выбора [5]. Рыб кормили 1–2 раза в сутки, размещался корм в один или несколько отсеков, в которых на момент наблюдения находились рыбы.
В общей сложности было исследовано 612 экз. сеголеток и двухлеток 19 видов рыб, кроме того, 48 экз. трехлеток и четырехлеток пескаря и обыкновенного гольяна соответственно. Все эксперименты выполнялись в двух повторностях. Данные по ОИТ фиксировались в виде средних значений. В связи с тем, что методические разработки А. М. Свирского и В. Г. Терещенко [8], а также анализ данных [4] показали, что ошибка определения ОИТ у группы особей в горизонтальных термоградиентных установках с учетом всех методических погрешностей составляет ±1 °С, различия показателей, превышающие 1 °С, считались достоверными.
В кратковременных (2–12 ч) опытах на примере молоди карповых и окуневых видов нами было показано, что температурный выбор начинается после посадки рыб в экспериментальную установку и предоставления особям термоградиентных условий. Непродолжительный (около 0,5–1,0 ч) период ознакомления с новой обстановкой, в которую попадали рыбы, сменялся активными поисковыми движениями в более теплые и холодные отсеки градиентной установки. При летней температуре аккли-мации (18–20 °С) преобладала тенденция выбора более высоких температурных зон. Так, сеголетки леща, густеры, карпа, плотвы и речного окуня уже через час концентрировались в диапазоне t 21–23 °С, а затем особи разных видов избирали еще более высокую температуру – от 25 до 29 °С. Отмечено, что выбор температуры группой рыб происходил быстрее по сравнению с одиночными особями. Результаты этих опытов подтвердили данные о процессе начального термоизбирания, полученные ранее другими авторами [7; 13; 15]. Такая поведенческая реакция была известна для многих животных – от простейших до позвоночных [7].
В начальный период исследования поведения и распределения рыб в термоградиентных условиях опыты проводились непродолжительное время (минуты, часы). Эксперименты с большей продолжительностью были единичны [22]. Тем не менее и за это время рыбы явно обнаруживали тенденцию к передвижению в более теплые, а иногда и в холодные зоны градиента. Отмечено также, что эта реакция, меняющая распределение рыб в градиенте фактора среды, самопроизвольна и представляет собой терморегуляционное поведение («thermoregula-tory behavior» в интерпретации иностранных исследователей). Было высказано предположение, что эта зона, видимо, представляет собой область эколого-физиологического оптимума роста и питания пойкилотермных и гомойотермных животных [7].
В естественных условиях возникающие градиенты температуры могут быть или кратковременными, или продолжительными. Представляло интерес выяснить, как поведут себя рыбы в экспериментальных термоградиентных условиях в течение более длительных промежутков времени – нескольких дней, одной или двух недель. С этой целью на молоди рыб проводились опыты продолжительностью 10–14 суток. Такие данные, полученные в горизонтальных термоградиентных установках, сходных с нашими, были единичными [16; 22].
В опытах продолжительностью 10–14 суток на молоди леща, синца, серебряного и золотого карася, карпа, плотвы, окуня, ерша, сибирского осетра, пеляди и радужной форели было выявлено, что процесс термоизбирания делится на два этапа – переходный (ИТ) и стабильного выбора (ОИТ). На первом этапе значения избираемой температуры значительно варьировались, на втором этапе происходила стабилизация температурного выбора. Наиболее быстрый (уже на 2-е – 3-и сутки опыта) выход в зону ОИТ был зафиксирован у серебряного и золотого карасей, а также у головешки-ротана.
Характерно, что опыты короткой продолжительности, особенно при использо- вании набора значений акклимационной температуры и методов графической интерполяции, также позволяют рассчитывать предполагаемые зоны ОИТ с ошибкой, не превышающей 1 °C [24]. Парадигма «конечного преферендума» Ф. Е. Дж. Фрая, сформулированная канадским исследователем намного раньше многочисленных экспериментов по поведенческой терморегуляции рыб в 1960–80 гг., заключается в двойном (бипартитном) определении термина «конечный термопреферендум» [17; 18]. Согласно одному из них, ОИТ – температура, при которой температуры акклимации и избираемые равны; согласно другому, ОИТ – температура, к которой гидробионты «притягиваются» (реакция «gravitation») или которой они достигают в градиенте действия фактора независимо от их температуры акклимации (термального прошлого).
Большинство исследователей в качестве критерия зоны ОИТ выбирают значения среднесуточной ИТ за одни, двое или трое суток, в течение которых разница этих значений у рыб в процессе опыта статистически недостоверна. Такой методический подход считается верным для всех типов градиентных установок, линейных горизонтальных, вертикальных, круговых, а также установок последнего поколения двухкамерных ихти-отронов, или шаттл-боксов [21; 23]. В наших последующих длительных опытах для определения зоны ОИТ был принят 3-суточный критерий стабильного избирания температуры как у молоди, так и у взрослых особей рыб.
Данные по термоизбиранию рыб в длительных экспериментах продолжительностью 10–14 суток были получены ранее лишь для некоторых видов рыб, обитающих в пресных водах США и Канады [14; 15; 22]. Для рыб из пресных водоемов России, а также региона Верхней Волги, такие результаты отсутствовали. Для характеристики видовых особенностей распределения и поведения рыб в искусственных (экспериментальных) термоградиентных условиях представлялось целесообразным исследование термоизби-рания у молоди разных видов рыб, относящихся к различным систематическим и экологическим группам. Такие эксперименты были проведены нами на молоди (в от- дельных случаях – на более взрослых особях) 21 вида рыб из 9 семейств (таблица). Время проведения опытов – июнь-август, температура акклимации в условиях лаборатории – 18±2 °С, для пеляди, радужной форели и налима – 15±2 °С.
Таблица
ОИТ у молоди различных видов рыб из региона Верхней Волги в сравнении с другими видами
|
Семейство |
Вид |
Возраст |
ОИТ, °С |
|
Сазан (карп) |
0+ – 1+ |
30,0 |
|
|
Карась золотой |
0+ – 1+ |
28,5 |
|
|
Карась серебряный |
0+ – 1+ |
27,5 |
|
|
Синец |
0+ – 1+ |
27,3 |
|
|
Уклейка |
0+ |
27,0 |
|
|
Карповые |
Лещ |
0+ – 1+ |
26,5 |
|
Густера |
0+ – 1+ |
26,0 |
|
|
Плотва |
0+ – 1+ |
26,0 |
|
|
Язь |
0+ – 1+ |
25,8 |
|
|
Пескарь |
2+ |
20,5 |
|
|
Гольян обыкновенный |
3+ |
16,8 |
|
|
Головешковые |
Головешка-ротан |
0+ – 1+ |
27,5 |
|
Речной окунь |
0+ – 1+ |
25,7 |
|
|
Окуневые |
Ерш |
0+ – 1+ |
24,8 |
|
Судак |
0+ – 1+ |
24,0 |
|
|
Вьюновые |
Вьюн |
1+ |
25,0 |
|
Щуковые |
Щука |
0+ |
24,3 |
|
Осетровые |
Сибирский осетр |
0+ |
22,5 |
|
Сиговые |
Пелядь |
0+ |
17,0 |
|
Лососевые |
Радужная форель |
0+ |
15,5 |
|
Налимовые |
Налим |
0+ |
15,0 |
Показательно, что в пределах только одного семейства карповых амплитуда значений ОИТ достигала 15,0 °С: от 30,0 °С у молоди карпа до 16,8 °С у более взрослых особей обыкновенного гольяна. Высоким уровнем ОИТ отличались сеголетки чужеродного вида-вселенца головешки-ротана. Следует отметить также высокие значения ОИТ у уклейки и язя. У представителей других семейств наблюдалось уменьшение значений ОИТ с 25,7 °С (речного окунь) до 15,0–15,5 °С (радужная форель и налим). В сравнении с речным окунем несколько меньшие значения ОИТ показаны у судака и ерша.
Значения ОИТ отличаются у тепло- и холодолюбивых видов рыб, а разница ОИТ у рыб внутри семейства может быть существенной или незначительной. Сеголетки щуки и окуня, начинающие хищничать в конце лета, имеют приблизительно такой же уро- вень ОИТ, как и сеголетки других видов рыб, питающихся зоопланктоном. Многие виды карповых выбирают очень близкие значения ОИТ, сходные с ОИТ окуневых видов и головешки-ротана. В то же время ОИТ у более взрослых обыкновенного гольяна и пескаря сходны с ОИТ сибирского осетра и холодолюбивых налима, радужной форели и пеляди.
Для видов, обитающих в регионе Верхней Волги или выращиваемых там же в рыбохозяйственных целях, характерны следующие показатели ОИТ. Так, в рамках семейства вьюновых двухлетки-трехлетки гольца Noemacheilus barbatulus , в отличие от вьюна, избирают сравнительно низкую температуру (ОИТ равен ~15,1) [2]. Сеголетки стерляди Acipenser ruthenus (L.) близки по уровню ОИТ к молоди сибирского осетра –23,6° С [9]. Другие два вида-вселенца, бычок-головач, Neogobius kessleri (Gunther)
и бычок-цуцик, Proterorhinus marmoratus (Pallas), почти не различались между собой по уровню ОИТ, равному ~22,0–23,0 °С [2].
Скорость выбора зоны ОИТ была наиболее быстрой (от 2 до 4 суток) у теплолюбивых карпа, серебряного и золотого карася, головешки-ротана, а также у холодолюбивых пеляди, радужной форели и налима. Прочие виды иногда достигали зоны ОИТ только на 6–9-е сутки. Принципиально поведение в градиенте температуры у молоди разных видов не различалось, хотя присутствовали и некоторые особенности. Многие карповые предпочитали «стайный» выбор температуры и держались только группой. Окунь, ерш и ротан часто затаивались в углах отсеков градиента, периодически меняя свое местоположение. Корм выедался практически сразу. У сеголеток радужной форели были отмечены элементы территориального поведения. В целом, распределение рыб по градиенту температуры в зоне ОИТ отражает их теплолюбивость и другие особенности экологии.
Таким образом, полученные данные представляют несомненный интерес для ихтиологов, экологов, гидробиологов, специалистов высшей школы и рыбного хозяйства. Прежде всего, значение ОИТ характеризует эколого-физиологический и, возможно, эволюционный оптимум [3; 4; 6]. Так,
М. Джоблинг на примере 49 видов рыб показал высокую корреляцию между температурой оптимального роста и значением ОИТ [20]. Такое же соответствие отмечено нами на примере 11 видов пресноводных бореальных видов рыб [4]. Высокие значения ОИТ у молоди карпа (28–30 °С), подтвержденные многими исследователями, не удивительны, учитывая его происхождение из районов Юго-Восточной Азии [4 ; 19]. Тем не менее выращивание молоди данного вида по экономическим и техническим причинам лучше проводить при более низкой температуре ~24 °С.
Полученные данные могут быть полезны для разработки нормативов тепловой нагрузки на водоемы, а также для прогноза распределения рыб в естественных водоемах и зонах сброса подогретых вод ГРЭС, АЭС и крупных промышленных предприятий. Они также должны учитываться при решении ряда вопросов аквакультуры и возможной оценке риска инвазий видов-вселенцев, что наблюдается в последнее время в регионах Верхней, Средней и Нижней Волги. Такие температурные критерии, как ИТ и ОИТ, наряду с другими данными, характеризующими температурные требования пресноводных рыб [1; 4; 6; 10], существенно расширяют наши представления о разных формах температурных адаптаций рыб.
Список литературы Экспериментальная оценка температурного оптимума у молоди пресноводных видов рыб методом термопреферендума
- Алабастер Дж. Критерии качества воды для пресноводных рыб/Дж. Алабастер, Р. Ллойд. -М.: Легкая и пищевая пром-ть, 1984. -384 с.
- Вербицкий В. Б. Окончательно избираемые температуры каспийских рыб-вселенцев (бычков головача и цуцика) в сравнении с двумя аборигенными видами (пескарем и гольцом)/В. Б. Вербицкий, И. В. Гибенко, А. М. Свирский//Поведение рыб: материалы докл. Междунар. конф. 1-4 нояб. 2005 г. Борок, Россия. -М.: АКВАРОС, 2005. -С. 58-62.
- Голованов В. К. Эколого-физиологические аспекты терморегуляционного поведения пресноводных рыб/В. К. Голованов//Поведение и распределение рыб: докл. 2-го Всерос. совещ. «Поведение рыб». -Борок, 1996. -С. 16-40.
- Голованов В. К. Эколого-физиологические закономерности распределения и поведения пресноводных рыб в термоградиентных условиях: автореф. дис.. д-ра биол. наук/В. К. Голованов. -М., 2012. -47 с.
- Голованов В. К. Сравнительный анализ окончательно избираемой и верхней летальной температуры у молоди некоторых видов пресноводных рыб/В. К. Голованов, А. К. Смирнов, Д. С. Капшай//Тр. Карел. НЦ РАН. (Сер. Эксперим. биология). -2012. -№ 2. -С. 70-75.
- Голованов В. К. Температурные требования рыб Рыбинского водохранилища и их реализация в естественных условиях/В. К. Голованов, А. М. Свирский, Е. И. Извеков//Современное состояние рыбных запасов Рыбинского водохранилища. -Ярославль: ЯрГТУ, 1997. -С. 92-123.
- Ивлев В. С. Экологический анализ распределения животных в градиентных температурных условиях/В. С. Ивлев, Х. А. Лейзерович//Тр. Мурман. морск. биол. ин-та. -М.; Л.: Изд-во АН СССР, 1960. -Вып. 1 (5). -С. 3-27.
- Свирский А. М. Точность определения температуры, избираемой рыбами в установке с горизонтальным термоградиентом/А. М. Свирский, В. Г. Терещенко//Биология внутренних вод: информац. бюл. -1992. -№ 92. -С. 85-88.
- Смирнов А. К. Избираемая температура молоди стерляди Acipenser ruthenus L./А. К. Смирнов//Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера: материалы XXVIII Междунар. конф., 5-8 окт. 2009 г. -Петрозаводск: КарНЦ РАН, 2009. -С. 511-514.
- Шмидт-Ниельсен К. Физиология животных: Приспособление и среда/К. Шмидт-Ниельсен. -М.: Мир, 1982. -Кн. 1. -416 с.
- Badenhuizen Т. Г. Temperature selected by Tilapia mossambica (Peters) in a test tank with a horisontal temperature gradient/T. R. Badenhuizen//Hydrobiologia. -1967. -Vol. 30, № 3-4. -P. 541-554.
- barans c. A. Temperatures selected seasonally by four fishes from Western Lake Erie/C. A. Barans, R. A. Tubb//J. Fish. Res. -1973. -Vol. 30, № 11. -P. 1697-1703.
- beitinger Т. L. Physiological and ecological correlates of preferred temperature in fish/T. L. Be-itinger, L. C. Fitzpatrick//Thermoregulation in ectotherms. Symp. Richmond, 1978. -Amer. Zool. -1979. -Vol. 19, № 1. -P. 319-329.
- cherry D. s. Biological monitoring. Part V: Preference and avoidance studies/D. S. Cherry, J. Jr. Cairns//Water Res. -1982. -Vol. 16, № 3. -P. 263-301.
- coutant c. c. Compilation of temperature preference data / C. C. Coutant // Temperature preference studies in environmental impact assessments: an overview with proce-dural recommendations. Proceed. Symp. and Panel Discuss. Northeast Fish and Wildlife Conf (Northeast Division, Amer. Fish. Soc.). Hershey. Pa. April 27, 1976 // J. Fish. Res. Board Can. — 1977. — Vol. 34, № 5. — P. 730—734.
- crawshaw L. J. Attainment of the final thermal preferendum in brown bullneads acclimated to different temperatures/L. J. Crawshaw//Comp. Biochem. Physiol. -1975. -Vol. 52A, № 1. -P. 171-173.
- Fry F. E. j. Effects of the environment on animal activity/F. E. J. Fry//Univ. Toronto Stud., Biol. Ser. -1947. -№ 55. -Publ. Ontario Fish. Res. Lab. -№ 68.-62 p.
- fry F. Е. j. The effect of environmental factors on the physiology of fish/F. E. J. Fry//Fish physiol. -Vol. VI. -N. Y. -1971. -P. 1-98.
- Golovanov V. K. The ecological and evolutionary aspects of thermoregulation behavior of fish/V. K. Golovanov//J. Ichthyology. -2006. -Vol. 46. -Suppl. 2. -P. 180-187.
- jobling M. Temperature tolerance and the final preferendum -rapid methods for the assessment of optimum growth temperature/M. Jobling//J. Fish. Biol. -1981. -Vol. 19, № 4. -P. 439-455.
- mccauley Г. w. Laboratory methods for determining temperature preference/R. W. McCauley//Temperature preference studies in environmental impact assessments: an overview with procedural recommendations. Proceed. Symp. and Panel Discuss. Northeast Fish and Wildlife Conf (Northeast Division, Amer. Fish. Soc.). Hershey. Pa. April 27. 1976/J. Fish. Res. Board Can. -1977. -Vol. 34, № 5. -P. 749-752.
- reutter j. M. Laboratory estimates of the seasonal final temperature preferenda of some Lake Erie fish/J. M. Reutter, C. E. Herdendorf//Proc. 17 th. Conf. Great Lakes Res. Hamilton. 1974. Part 1. Ann. Arbor., Mich. -1974. -P. 59-67.
- reynolds w. w. Temperature preference of four fish species in an electronic thermoregulatory shuttlebox/W. W. Reynolds, M. E. Casterlin//Progr. Fish-Cult. -1977. -Vol. 39, № 3. -P. 123-125.
- reynolds w. w. Behavioral thermoregulation and the «final preferendum» paradigm/W. W. Reynolds, M. E. Casterlin//Thermoregulation in ectotherms. Symp. Richmond. 1978/Amer. Zool. -1979. -Vol. 19, № 1. -P. 211-224.
