Экспрессия микроРНК в опухолевой ткани у больных раком гортани

Автор: Никитина Екатерина Геннадьевна, Черемисина Ольга Владимировна, Бычков Вячеслав Алексеевич, Кульбакин Денис Евгеньевич, Чойнзонов Евгений Лхамацыренович, Стегний Владимир Николаевич, Литвяков Николай Васильевич
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 2 (68), 2015 года.
Бесплатный доступ
Изучен аберрантный паттерн экспрессии микроРНК-18а, -21, -155, -200а, -200с, -205, -221, -494 в опухолевой ткани плоскоклеточной карциномы гортани относительно прилежащей нормальной ткани у 46 пациентов. Установлено, что клинико-морфологические параметры не оказывают влияния на экспрессию анализируемых микроРНК. Показана гиперэкспрессия онкогенных микроРНК-21, -155, -205 и гипоэкспрессия онкосупрессорной микроРНК-200а в опухолевой ткани относительно прилежащей неизмененной ткани. Представленные результаты позволяют предполагать существенную роль этих микроРНК в канцерогенезе опухолей гортани.
Рак гортани, микрорнк, экспрессия microrna
Короткий адрес: https://sciup.org/14056526
IDR: 14056526 | УДК: 616.22-006.6,
Expression microRNA in larynx cancer
MicroRNA are nonprotein-coding small ribonucleic acids controlling gene expression at post-transcriptional level. MicroRNA technically could controll 2/3 of human genome. At the present study abnormal expression of microRNA-18а, -21, -155, -200а, -200с, -205, -221, -494 in malignant tissue of squamous cell larynx carcinoma of 46 patients were studied. There was no statistically significant correlation between microRNA expression and pathological characteristics. Our data showed that oncogenic microRNA-21, -155, -205 were overexpressed as well as oncosypression microRNA-200a were underexpressed in larynx cancer compared to adjacent normal epithelium.
Текст научной статьи Экспрессия микроРНК в опухолевой ткани у больных раком гортани
МикроРНК постранскрипционно регулируют генную экспрессию, связываясь с 3’-UTR мРНК генов-мишеней по принципу комплементарности, ингибируя или полностью инактивируя мРНК [3, 31]. Благодаря такому механизму микроРНК участвуют в регуляции многих клеточных процессов, включая развитие, дифференцировку, апоптоз, пролиферацию, ответ на стрессовое воздействие, метаболизм и секрецию инсулина [2].
В опухоли часто наблюдается аберрантная экспрессия микроРНК, которая свидетельствует о важной роли этих молекул в процессе канцерогенеза [12]. Аберрантная экспрессия микроРНК может быть следствием различных причин, таких как делеции, амплификации хромосомных локусов микроРНК, мутации или нарушение регуляции транскрипционных факторов, регулирующих экспрессию микроРНК. Влияние на микроРНК могут оказывать и эпигенетические механизмы [17, 23]. Экспрессия микроРНК отличается значительной тканевой специфичностью, и аберрантный паттерн экспрессии микроРНК различается в зависимости от тканевого происхождения опухоли. Описание этого аберрантного паттерна важно с точки зрения понимания механизмов канцерогенеза, поиска новых мишеней для таргетной терапии и маркеров для диагностики, прогноза и мониторинга заболевания. Одной из малоизученных локализаций в этом отношении являются плоскоклеточные карциномы гортани, которые по частоте заболеваемости занимают второе место в структуре опухолей головы и шеи
[8]. Среди микроРНК, для которых было проведено исследование аберрантного паттерна экспрессии в опухоли гортани, были выбраны те молекулы, относительно которых в литературе представлены данные, доказывающие их значительную роль в канцерогенезе при других локализациях. Это микроРНК-21, -18а, -200а, -200с, -205, -221, -494. Было показано, что они играют одну из ключевых ролей в регуляции процессов клеточного цикла, апоптоза, роста, инвазии, пролиферации клеток при опухолях репродуктивных органов у женщин (рак молочной железы, рак яичников), глиобластомах и многих других локализациях [9, 19, 20, 24].
Целью исследования явилось изучение аберрантного паттерна экспрессии микроРНК -18а, -21, -155, -200а, -200с, -205, -221, -494 в опухолевой ткани плоскоклеточной карциномы гортани относительно нормальной прилежащей ткани и связи этого показателя с клинико-морфологическими характеристиками опухолевого процесса.
Материал и методы
В исследование включены 46 больных раком гортани (табл. 1), мужского пола, в возрасте от 45 до 64 лет (в среднем – 56 ± 10 лет), проходивших обследование и лечение в Томском НИИ онкологии. Работа проведена с соблюдением принципов добровольности и конфиденциальности в соответствии с «Основами законодательства РФ об охране здоровья граждан» (Указ президента РФ 39 от 24.12.93 № 2288), получено разрешение
таблица 1
Клинико-морфологические характеристики пациентов
В работе был использован парный операцион-ный/биопсийный материал (опухолевая и прилежащая нормальная ткань) от пациентов с первичным диагнозом плоскоклеточного рака гортани (РГ), не получавших на момент исследования специального лечения. Биопсийный материал из опухолевой и неизмененной ткани получали при видеоларингоскопии. Диагноз у всех пациентов был морфологически верифицирован. Биологический материал помещали в пробирки типа Eppendorf с консервирующим раствором RNAlater (Sigma, USA), пробирки выдерживали сутки при 4°С и потом хранили при –80°С до выделения РНК. Тотальная фракция РНК из ткани была выделена набором реагентов mirVana™ (Ambion, USA). Количество РНК и чистоту выделения оценивали на спектрофотометре NanoDrop 2000 (Thermo-Scientific, USA) (концентрация РНК варьировала от 30 до 800 нг/ мкл, А260/280=1,90). Значение RIN (RNA Integrity Number) колебалось от 8,0 до 9,4 (оценивали на приборе TapeStation 2200 Agilent Technologies, USA). Мультиплексная обратно транскриптазная ПЦР (ОТ-ПЦР) проводилась в объеме 20 мкл, в состав которого входили 100–500 нг РНК-матрицы, 0,5 нM микроРНК-специфичного праймера особой шпилевидной конструкции (hsa-miR-18a-5p, hsa-miR-21-5p, hsa-miR-155-5p, hsa-miR-200a-3p, hsa-miR-200c-3p, hsa-miR-205-5p, hsa-miR-221-3p, hsa-miR-494-3p) [18], 1 ед. ингибитора РНКаз, 50 ед. MMLV ревертазы («Сибэнзим», Новосибирск), 1x MMLV буфера и 1 мM каждого dNTP. Протокол реакции: 38 циклов при 16°C в течение 20 сек, при 42°C в течение 20 сек, при 50°C в течение 1 сек, с заключительным этапом прогрева смеси при 85°C в течение 1 мин. При каждой постановке были включены отрицательные контроли, не содержащие РНК и/или несущие в составе геномную ДНК. Все реакции ПЦР в режиме реального времени проведены в триплете согласно протоколу [18]. В качестве гена-рефери выбрана микроРНК-103 [26], калибровка выполнена относительно прилежащей к опухоли нормальной ткани у каждого пациента, уровень экспрессии микроРНК рассчитан согласно методу Pfaffl [27]. В качестве результата использовано логарифмически трансформированное по основанию е нормализованное значение уровня экспрессии (ln(fold+const)).
Корреляционный анализ, а также обработка данных с применением t-критерия Уэлша и Стьюдента проводились в программе Statistica 8.0. Для всех статистических подсчетов применен критерий множественности сравнения Бенджамини – Хох-берга (FDR – false discovery rate). Все значения р, удовлетворяющие условию FDR ≤ 5 %, принимались как статистически значимые.
Результаты исследования
На первом этапе было изучено возможное влияние на экспрессию микроРНК таких параметров, как стадия распространенности процесса, статус лимфогенного метастазирования и степень дифференцировки опухоли путем сравнительной оценки показателей уровня экспрессии микроРНК в подгруппах. Ввиду малочисленности выборки были объединены пациенты с I–II и III–IV стадиями опухолевого процесса. Сравнение этих групп не выявило значимых различий в уровне экспрессии анализируемых микроРНК. Не обнаружено влияния лимфогенного метастазирования и степени дифференцировки опухолевой ткани на экспрессию анализируемых микроРНК (табл. 2). Хотя для микроРНК-200с показано снижение уровня экспрессии у пациентов без метастазирования в сравнении с подгруппой лиц с лимфогенными метастазами, для микроРНК-18а наблюдается тенденция к повышению уровня экспрессии от низкой к высокой степени дифференцировки опухоли. Однако с учетом поправки FDR данные значения р не являются статистически значимыми (табл. 2). Таким образом, клинико-морфологические параметры не оказывали статистически значимого влияния на экспрессию микроРНК в опухолевой ткани гортани. Это позволило анализировать аберрантный паттерн экспрессии микроРНК в общей группе.
Установлено, что для некоторых микроРНК выявлен аберрантный уровень экспрессии в опухолевой ткани относительно прилежащей нормальной ткани пациентов. Показана статистически значимая гиперэкспрессия микроРНК-21, -155 и -205 (р=0,00005, р=0,00008 и р=0,00085 соответственно). Частота встречаемости случаев с гиперэкспрессией по этим микроРНК в 2,3–4,8
таблица 2
Уровень экспрессии микрорнК в опухоли относительно прилежащей нормальной ткани пациентов в зависимости от клинико-морфологических параметров
|
Параметры |
МикроРНК |
-18а |
-21 |
-155 |
-200а |
-200с |
-205 |
-221 |
-494 |
|
Распространенность первичной опухоли |
Т |
0,20 ± 0,21 |
0,64 ± 0,27 |
0,67 ± 0,33 |
-1,05 ± 0,30 |
-0,22 ± 0,19 |
0,50 ± 0,15 |
-0,70 ± 0,32 |
-0,29 ± 0,39 |
|
Т |
0,26 ± 0,15 |
1,27 ± 0,33 |
0,84 ± 0,17 |
-0,69 ± 0,36 |
-0,01 ± 0,30 |
0,81 ± 0,39 |
0,06 ± 0,31 |
0,09 ± 0,33 |
|
|
р |
0,15 |
0,66 |
0,45 |
0,56 |
0,46 |
0,09 |
0,46 |
0,83 |
|
|
FDR |
58,3 |
75,2 |
119,7 |
74,2 |
74,1 |
74,2 |
91,3 |
82,9 |
|
|
Лимфогенное метастазирование |
N0 |
0,16 ± 0,14 |
0,85 ± 0,24 |
0,72 ± 0,21 |
-0,82 ± 0,28 |
-0,35 ± 0,20 |
0,63 ± 0,25 |
-0,27 ± 0,24 |
-0,11 ± 0,32 |
|
N 1-2 |
0,26 ± 0,23 |
1,82 ± 0,57 |
1,04 ± 0,37 |
-0,82 ± 0,50 |
0,87 ± 0,46 |
0,53 ± 0,49 |
-0,35 ± 0,76 |
0,04 ± 0,25 |
|
|
р |
0,696 |
0,123 |
0,449 |
0,993 |
0,018 |
0,858 |
0,923 |
0,717 |
|
|
FDR |
139,2 |
49,2 |
119,8 |
99,3 |
14,5 |
114,5 |
105,5 |
114,8 |
|
|
Степень дифференцировки опухоли |
Низкая |
0,29 ± 0,18 |
0,87 ± 0,68 |
0,61 ± 0,39 |
-0,60 ± 0,51 |
-0,65 ± 0,62 |
0,37 ± 0,61 |
0,15 ± 0,11 |
-1,14 ± 1,05 |
|
Умеренная |
0,31 ± 0,15 |
0,97 ± 0,27 |
0,74 ± 0,22 |
-0,91 ± 0,31 |
-0,1 ± 0,221 |
0,69 ± 0,28 |
-0,40 ± 0,31 |
0,07 ± 0,28 |
|
|
Высокая |
0,68 ± 0,57 |
0,34 ± 0,68 |
0,62 ± 0,70 |
-0,94 ± 0,70 |
0,19 ± 0,20 |
0,47 ± 0,38 |
-0,38 ± 0,30 |
-0,06 ± 0,53 |
|
|
р1 |
0,01 |
0,89 |
0,77 |
0,6 |
0,41 |
0,63 |
0,1 |
0,27 |
|
|
FDR, % |
12 |
89,4 |
88,5 |
95,9 |
82,8 |
84,6 |
39,7 |
72,9 |
|
|
р2 |
0,14 |
0,59 |
0,99 |
0,7 |
0,23 |
0,9 |
0,14 |
0,39 |
|
|
FDR, % |
57,6 |
94,8 |
99,2 |
93,7 |
61,9 |
102,5 |
112,8 |
77,3 |
|
|
р3 |
0,53 |
0,39 |
0,87 |
0,97 |
0,32 |
0,64 |
0,97 |
0,83 |
|
|
FDR, % |
141,8 |
156,9 |
115,9 |
97,3 |
257 |
127,7 |
110,7 |
132,4 |
Примечание: р – уровень значимости рассчитан по t-критерию Уэлша; р1 – «низкий» vs «умеренный», р2 – «низкий» vs «высокий», р3 – «умеренный» vs «высокий»; FDR – поправка Бенджамини–Хохберга (FDR – false discovery rate).
таблица 3
Уровень экспрессии микрорнК в опухоли относительно прилежащей нормальной ткани
|
МикроРНК |
Уровень экспрессии |
р |
FDR |
Число случаев с гиперэкспрессией |
Число случаев с гипоэкспрессией |
|
-18а (n=45) |
0,23 ± 0,83 |
0,071 |
11,39 |
27 (61 %) |
18 (39 %) |
|
-21 (n=46) |
0,98 ± 1,49 |
0,00005 |
0,04 |
38 (83 %) |
8 (17 %) |
|
-155 (n=46) |
0,76 ± 1,19 |
0,00008 |
0,03 |
34 (74 %) |
12 (26 %) |
|
-200а (n=46) |
-0,85 ± 1,61 |
0,0008 |
0,22 |
14 (30 %) |
32 (70 %) |
|
-200с (n=46) |
-0,11 ± 1,24 |
0,569 |
65,05 |
21 (45 %) |
25 (54 %) |
|
-205 (n=44) |
0,66 ± 1,43 |
0,003 |
0,75 |
37 (81 %) |
9 (19 %) |
|
-221 (n=43) |
-0,31 ± 1,48 |
0,174 |
23,24 |
25 (54 %) |
21 (46 %) |
|
-494 (n=43) |
-0,08 ± 1,64 |
0,762 |
76,21 |
22 (43 %) |
26 (56 %) |
Примечание: р – уровень значимости рассчитан по t-критерию Стьюдента (t-for single means); FDR – поправка Бенджамини–Хохберга.
раза выше, чем гипоэкспрессии. Кроме повышенной экспрессии, для микроРНК-200а отмечается гипоэкспрессия (р=0,0008), и частота случаев с гипоэкспрессией микроРНК-200а в 4,3 раза выше, чем частота гиперэкспрессии (табл. 3).
Для микроРНК, как и для многих молекулярных регуляторов клетки и организма в целом, характерно образование сложной системы взаимодействий друг с другом [14, 32]. На основе корреляционного анализа была показана такая сеть взаимодействий для изученных микроРНК (рис. 1). Выявлена положительная корреляция для микроРНК-21 и -155 (r=0,31, р=0,00001), микроРНК-200с и -18а (r=0,38, р=0,001), 200с и -205 (r=0,34, р=0,025), -200а и -221 (r=0,35, р=0,021) (рис. 1), а также наблюдается тенденция к положительной корреляции между микроРНК-21 и -200с (r=0,31, р=0,036), однако значение р не удовлетворяет 5 % поправке FDR.
Обсуждение
Полученные нами результаты по гиперэкспрессии микроРНК-21, -155 и -205 в опухоли гортани согласуются с данными литературы. J. Ren et al. [29] и P. Cao et al. [5] выявили гиперэкспрессию микроРНК-21 при раке гортани. Результаты по микроРНК-155 при злокачественных опухолях головы и шеи частично соотносятся с данными литературы, имеются отдельные работы, в которых показаны как гиперэкспрессия [6, 16], так и отсутствие значимых изменений ее экспрессии в опухолевой ткани [5]. В целом данные относительно экспрессии микроРНК-205 при раке гортани противоречивы. P. Cao et al. [5] выявили значимую гиперэкспрессию микроРНК-205, однако L. Tian et al. [30] в исследованиях in vitro (культура Hep-2 клеток) и in vivo (n=30) обнаружили гипоэкспрессию этой микроРНК при раке гортани, что авторы
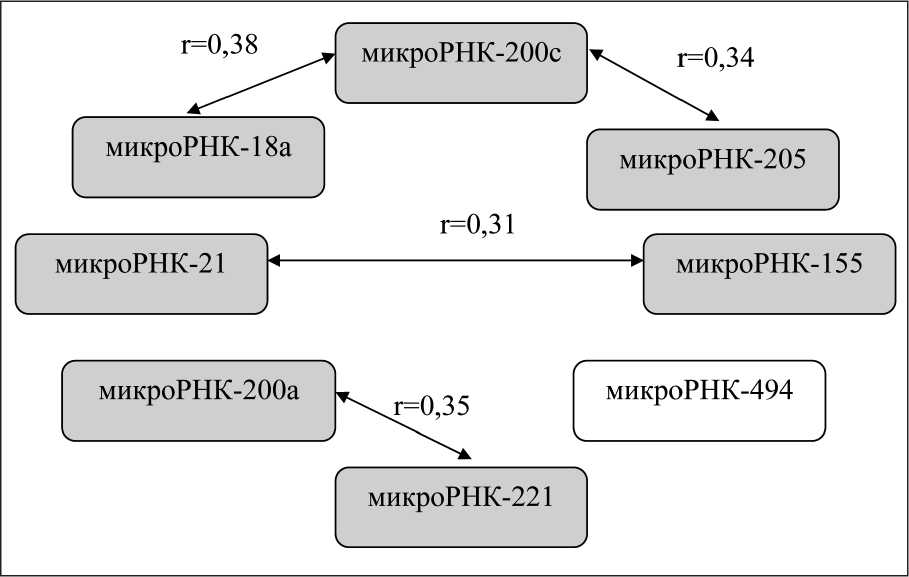
Рис. 1. Корреляционные связи между показателями экспрессии изучаемых микроРНК в опухолевой ткани у больных РГ
объясняют возможными влияниями выбора гистотипов опухоли, ее подтипов, микроокружения и регуляции генов-мишеней. Результаты исследования и данные литературы позволяют предполагать, что микроРНК-21, -155 и -205 играют онкогенную роль при опухолях гортани, и повышение их экспрессии связано с регуляцией различных процессов, задействованных в канцерогенезе.
Онкогенную функцию этих микроРНК подтверждает список генов-мишеней, экспрессию которых регулируют данные микроРНК. В настоящее время для микроРНК-21 валидированы следующие мишени: PDCD4 (programmed cell death protein 4), RECK (reversion inducing cysteine-rich protein with kazal motifs), maspin (mammary serine protease inhibitor), NFIB (nuclear factor I), TPM1 (Tropomyosin 1), SPRY2 (Sprouty2), PTEN (phosphatase and tensin homologue), Tap63 (transformation-related protein 63), лиганд Fas, Cdc25a (cyclin-dependent kinases 25), HNRPK (Heterogeneous nuclear ribonucleoprotein K). Все перечисленные гены-мишени микроРНК-21 принимают непосредственное участие в трансформации клеток [15, 20].
МикроРНК-155 принимает участие в регуляции нескольких ключевых процессов канцерогенеза через ингибирование генов-мишеней, задействованных в TGF-β/Smad-индуцированном эпителиально-мезенхимальном переходе, гликолизе, JNK2/STAT3 пути, NF-kB и AKT путях. Представлены убедительные доказательства того, что мишенями этой микроРНК являются: FOXO3a (Forkhead box O3), C/EBPβ (CCAAT-enhancer-binding proteins), которая, в свою очередь, активирует экспрессию hexokinase 2, STAT3 (Signal Transducer and Activator of Transcription), действие которого проявляется через репрессию гена SOCS1 (suppressor of cytokine signaling 1), CDC73 (Cell division cycle 73) и ряд других [7].
Относительно микроРНК-205 отмечается ее дуалистическая роль в канцерогенезе. Онкосу- прессорную роль микроРНК-205 осуществляет путем подавления пролиферации при ЗНО, репрессируя экспрессию таких генов-мишеней, как ErbB3 (Receptor tyrosine-protein kinase erbB-3), E2F1(Transcription factor E2F1) и гены MAPK- и AR-сигнальных путей, PKCε (Protein kinase C epsilon type), ZEB1/2, ММР2 и ММР9, VEGF-A (Vascular endothelial growth factor). Имеются сообщения, свидетельствующие и об онкогенной роли микроРНК-205, которая участвует в малигнизации, прогрессии опухоли и развитии устойчивости к противоопухолевой терапии, репрессируя гены PTEN, CYR61 (Cysteine-rich angiogenic inducer 61) и CTGF (connective tissue growth factor), SHIP2 (enzyme phosphatidylinositol-3,4,5-trisphosphate 5-phosphatase), DHFR (Dihydrofolate reductase) [28].
Наши результаты относительно гипоэкспрессии микроРНК-200а при раке гортани согласуются и дополняют полученные ранее данные других авторов, которые показали онкосупрессорную роль этой микроРНК в канцерогенезе опухолей разных локализаций [13]. Для микроРНК-200а валидированы такие гены-мишени, как: ZEB1 и 2 (Zinc finger E-box-binding homeobox 1 и 2) [25], ERRFI-1 (ERBB receptor feedback inhibitor 1) [1], CTNNB1 (cadherin-associated protein) [22] и ряд других. Эти данные позволили заключить, что микроРНК-200а играет ключевую роль в эпителиально-мезенхимальном переходе и процессе приобретения клеткой эмбрионального фенотипа [4, 19].
Результаты корреляционного анализа могут свидетельствовать о наличии связи между этими микроРНК, в основе которой лежит их вовлеченность в процессы внутриклеточной регуляции общих генов-мишеней. Такие гены-мишени найдены для всех связанных пар. МикроРНК-21 и -155 объединяет в первую очередь их взаимодействие с сигнальным белком и активатором транскрипции STAT3 [21]. На микроРНК-18а и -200с оказывает влияние репрограммирующий фактор c-Myc [10]. МикроРНК-200с и -205 играют определенную роль в эпителиально-мезенхимальном переходе и процессе приобретения клеткой эмбрионального фенотипа посредством регуляции экспрессии генов ZEB1 и ZEB2 [13]. Пары микроРНК-221 и 200а могут быть связаны через ген р27Kip1, который играет существенную роль в регуляции перехода от G1 к S-фазе клеточного цикла [11].
Заключение
Таким образом, установлены гиперэкспрессия онкогенных микроРНК-21, -155, 205 и гипоэк- спрессия онкосупрессорной микроРНК-200а в опухолевой ткани гортани пациентов с первичным диагнозом рака гортани, что согласуется с данными литературы, полученными для этих микроРНК на примере других локализаций. Эти данные позволяют предположить значительную роль микроРНК21, -155, -200а и -205 в канцерогенезе опухолей гортани, что, вероятно, связано с их участием в регуляции многих генов-мишеней и клеточных процессов, задействованных в малигнизации.
Работа выполнена в рамках программы повышения конкурентоспособности Национального исследовательского Томского государственного университета.
Список литературы Экспрессия микроРНК в опухолевой ткани у больных раком гортани
- Adam L., Zhong M., Choi W., Qi W., Nicoloso M., Arora A., Calin G., Wang H., Siefker-Radtke A., McConkey D. miR-200 expression regulates epithelial-to-mesenchymal transition in bladder cancer cells and reverses resistance to epidermal growth factor receptor therapy//Clin. Cancer Res. 2009. Vol. 15 (16). P. 5060-5072 DOI: 10.1158/1078-0432.CCR-08-2245
- Ambros V. The functions of animal microRNAs//Nature. 2004. Vol. 431. (7006). P. 350-355.
- Bartel D.P. MicroRNAs: genomics, biogenesis, mechanism, and function//Cell. 2004. Vol. 116 (2). P. 281-297.
- Bracken C.P., Gregory P.A., Kolesnikoff N., Bert A.G., Wang J., Shannon M.F., Goodall G.J. A double-negative feedback loop between ZEB1-SIP1 and the microRNA-200 family regulates epithelial-mesenchymal transition//Cancer Res. 2008. Vol. 68, № 19. P. 7846-7854 DOI: 10.1158/0008-5472.CAN-08-1942
- Cao P., Zhou L., Zhang J., Zheng F., Wang H., Ma D., Tian J. Comprehensive expression profiling of microRNAs in laryngeal squamous cell carcinoma//Head Neck. 2013. Vol. 35 (5). P. 720-728 DOI: 10.1002/hed.23011
- Chang S.S., Jiang W.W., Smith I., Poeta L.M., Begum S., Glazer C., Shan S., Westra W., Sidransky D., Califano J.A. MicroRNA alterations in head and neck squamous cell carcinoma//Intern. J. Cancer. 2008. Vol. 123 (12). P. 2791-2797 DOI: 10.1002/ijc.23831
- Chen Z., Ma T., Huang C., Hu T., Li J. The Pivotal Role of microRNA-155 in the Control of Cancer//Journal of Cellular Physiology. 2014. Vol. 229 (5). P. 545-550 DOI: 10.1002/jcp.24492
- Chu E.A., Kim Y.J. Laryngeal cancer: diagnosis and preoperative work-up//Otolaryngol. Clin. North Am. 2008. Vol. 41 (4). P. 673-695 DOI: 10.1016/j.otc.2008.01.016
- Cochrane D.R., Howe E.N., Spoelstra N.S., Richer J.K. Loss of miR-200c: a marker of aggressiveness and chemoresistance in female reproductive cancers//J. Oncol. 2009. Vol. 2010: 821717 DOI: 10.1155/2010/821717
- Concepcion C.P., Bonetti C., Ventura A. The miR-17-92 family of microRNA clusters in development and disease//Cancer J. 2012. Vol. 18(3). P. 262-267 DOI: 10.1097/PPO.0b013e318258b60a
- Di Martino M.T., Gullà A., Cantafio M.E.G., Lionetti M., Leone E., Amodio N., Guzzi P.H., Foresta U., Conforti F., Cannataro M. In vitro and in vivo anti-tumor activity of miR-221/222 inhibitors in multiple myeloma//Oncotarget. 2013. Vol. 4 (2). P. 242-255.
- Esquela-Kerscher A., Slack F.J. Oncomirs -microRNAs with a role in cancer//Nature Rev. Cancer. 2006. Vol. 6 (4). P. 259-269.
- Feng X., Wang Z., Fillmore R., Xi Y. MiR-200, a new star miRNA in human cancer//Cancer lett. 2014. Vol. 344 (2). P. 166-173 DOI: 10.1016/j.canlet.2013.11.004
- Gurtan A.M., Sharp P.A. The role of miRNAs in regulating gene expression networks//J. Mol. Biol. 2013. Vol. 425 (19). P. 3582-3600 DOI: 10.1016/j.jmb.2013.03.007
- Hong L., Han Y., Zhang Y., Zhang H., Zhao Q., Wu K., Fan D. MicroRNA-21: a therapeutic target for reversing drug resistance in cancer//Expert Opin. Ther. Targets. 2013. Vol. 17 (9) P. 1073-1080 DOI: 10.1517/14728222.2013.819853
- Hui A.B., Lenarduzzi M., Krushel T., Waldron L., Pintilie M., Shi W., Perez-Ordonez B., Jurisica I., O’Sullivan B., Waldron J. Comprehensive MicroRNA profiling for head and neck squamous cell carcinomas//Clin. Cancer Res. 2010. Vol. 16 (4). P. 1129-1139 DOI: 10.1158/1078-0432.CCR-09-2166
- Iorio M.V., Croce C.M. MicroRNA dysregulation in cancer: diagnostics, monitoring and therapeutics. A comprehensive review//EMBO Mol. Med. 2012. Vol. 4 (3). P. 143-159 DOI: 10.1002/emmm.201100209
- Iyevleva A.G., Kuligina E.S., Mitiushkina N.V., Togo A.V., Miki Y., Imyanitov E.N. High level of miR-21, miR-10b, and miR-31 expression in bilateral vs. unilateral breast carcinomas//Breast Cancer Res. Treat. 2012. Vol. 131 (3). P. 1049-1059 DOI: 10.1007/s10549-011-1845-z
- Korpal M., Lee E.S., Hu G., Kang Y. The miR-200 family inhibits epithelial-mesenchymal transition and cancer cell migration by direct targeting of E-cadherin transcriptional repressors ZEB1 and ZEB2//J. Biol. Chem. 2008. Vol. 283 (22). P. 14910-14914 DOI: 10.1074/jbc.C800074200
- Krichevsky A.M., Gabriely G. miR-21: a small multi-faceted RNA//J. Cell. Mol. Med. 2009. Vol. 13 (1). P. 39-53 DOI: 10.1111/j.1582-4934.2008.00556.x
- Löffler D., Brocke-Heidrich K., Pfeifer G., Stocsits C., Hackermüller J., Kretzschmar A.K., Burger R., Gramatzki M., Blumert C., Bauer K., Cvijic H., Ullmann A.K., Stadler P.F., Horn F. Interleukin-6 -dependent survival of multiple myeloma cells involves the Stat3-mediated induction of microRNA-21 through a highly conserved enhancer//Blood. 2007. Vol. 110 (4). P. 1330-1333.
- Mongroo P.S., Rustgi A.K. The role of the miR-200 family in epithelial-mesenchymal transition//Cancer Biol. Ther. 2010. Vol. 10 (3). P. 219-222.
- Nikitina E., Urazova L., Stegny V. MicroRNAs and human cancer//Exp. Oncol. 2012. Vol. 34 (1). P. 2-8.
- O’Donnell K.A., Wentzel E.A., Zeller K.I., Dang C.V., Mendell J.T. c-Myc-regulated microRNAs modulate E2F1 expression//Nature. 2005. Vol. 435 (7043). P. 839-843.
- Park S.-M., Gaur A.B., Lengyel E., Peter M.E. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2//Genes Dev. 2008. Vol. 22. (7). P.894-907 DOI: 10.1101/gad.1640608
- Peltier H.J., Latham G.J. Normalization of microRNA expression levels in quantitative RT-PCR assays: identification of suitable reference RNA targets in normal and cancerous human solid tissues//RNA. 2008. Vol. 14 (5). P. 844-852 DOI: 10.1261/rna.939908
- Pfaffl M.W. A new mathematical model for relative quantification in real-time RT-PCR//Nucleic Acids Res. 2001. Vol. 29 (9). P. e45-e45.
- Qin A.-Y., Zhang X.-W., Liu L., Yu J.-P., Li H., Wang S.-Z.E., Ren X.-B., Cao S. MiR-205 in cancer: An angel or a devil?//Eur. J. Cell Biol. 2013. Vol. 92. (2). P. 54-60.
- Ren J., Zhu D., Liu M., Sun Y., Tian L. Downregulation of miR-21 modulates Ras expression to promote apoptosis and suppress invasion of Laryngeal squamous cell carcinoma//Eur. J. Cancer. 2010. Vol. 46 (18). P. 3409-3416 DOI: 10.1016/j.ejca.2010.07.047
- Tian L., Zhang J., Ge J., Xiao H., Lu J., Fu S., Liu M., Sun Y. MicroRNA-205 suppresses proliferation and promotes apoptosis in laryngeal squamous cell carcinoma//Med. Oncol. 2014. Vol. 31 (1). P. 785 DOI: 10.1007/s12032-013-0785-3
- Zamore P.D., Haley B. Ribo-gnome: the big world of small RNAs//Science. 2005. Vol. 309 (5740). P. 1519-1524.
- Zhou J.-J., Zheng S., Sun L.-F., Zheng L. MicroRNA regulation network in colorectal cancer metastasis//World J. Biol. Chem. 2014. Vol. 5 (3). P. 301 DOI: 10.4331/wjbc.v5.i3.301
