Электрофизиологические параметры потенциалов действия клеток типа скрытого водителя ритма сердца кролика и мыши при действии лидокаина

Автор: Лебедева Е.А.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 3 (15), 2013 года.
Бесплатный доступ
Эффекты лидокаина, блокатора Na+-каналов, на клетки, работающие в режиме скрытого водителя ритма, синусно-предсердного (СП) узла кролика и мыши, были исследованы с помощью стандартной микроэлектродной техники. Эксперименты показали, что лидокаин дозозависимо замедлял скорость фазы быстрой деполяризации (dV/dtmax) потенциала действия у кролика и мыши. Полученные результаты свидетельствуют о значительном вкладе Na+-каналов в формирование фазы быстрой деполяризации клеток СП области.
Синусно-предсердный узел, лидокаин, ионные каналы, кролик, мышь
Короткий адрес: https://sciup.org/14992625
IDR: 14992625 | УДК: 612.17
Electrophysiological parameters of action potentials of latent pacemaker cells of rabbit and mouse heart under the influence of lidocaine
The effects of lidocaine on the latent pacemaker cells of sinoatrial (SP) node of rabbit and mouse were studied using standard microelectrode technique. Experiments have shown that lidocaine dose-dependently slows down the AP fast depolarization phase (dV/dtmax) in rabbit and mouse. The results obtained testifyto considerable contribution of Na+-channels in formation of cells fast depolari-zation phase of sinoatrial area.
Текст научной статьи Электрофизиологические параметры потенциалов действия клеток типа скрытого водителя ритма сердца кролика и мыши при действии лидокаина
Синусно-предсердный (СП) узел представляет собой группу клеток, способных самостоятельно генерировать импульсы без участия нервной системы, что обеспечивает одно из главных свойств сердца – автоматию. В отличие от потенциала действия (ПД) клеток рабочего миокарда, клетки СП узла имеют фазу медленной диастолической деполяризации (МДД), которая при достижении порога и переходит в ПД. Несмотря на продолжительное изучение, механизмы, лежащие в основе генерации пейсме-керной активности, до сих пор не установлены.
В частности, до конца не ясна роль Na+-ка-налов. Долгое время считалось, что ионы натрия не участвуют в фазе быстрой деполяризации и значение Na+-тока в пейсмекерной активности клеток СП узла незначительно [1, 2]. Недавние исследования показали, что, кроме сердечной изоформы Na v 1.5 Na+-каналов, в СП узле также присутствует и нейрональная изоформа Na v 1.1, которая располагается равномерно по всему СП узлу и имеет более высокую чувствительность к блокаторам, чем сердечная изоформа [3, 4]. Кроме того, мутация гена Scn5a, кодирующего Na+-каналы, была найдена у многих пациентов с синдромом слабости СП узла [5].
Лидокаин – один из блокаторов потенциалзависимых Na+-каналов, антиаритмик I класса, широко применяемый в кардиологической практике и в качестве локального анестетика. Однако механизм его действия на клетки синусно-предсердной области до сих пор до конца не ясен. Эксперименты на изолированных предсердиях крыс показали, что лидокаин слабо снижает частоту генерации спонтанного ритма [6]. Но работы, проведенные на изолированных миоцитах СП узла кролика [7] и изолированном СП узле мыши [8], демонстрируют более сильное влияние лидокаина на пейсмекерную активность клеток этой области.
Таким образом, цель нашей работы заключалась в исследовании Na+-тока, участвующего в формировании фазы быстрой деполяризации, а также динамики изменения других электрофизиологических параметров ПД при различных концентрациях лидокаина у кролика и мыши.
Материал и методы
Опыты проводили на кроликах породы Шиншилла (возраст 6–7 месяцев, масса тела 3,0–3,5 кг) и белых мышах (возраст 8–16 недель, масса тела
30±5 г). Животных наркотизировали введением зо-летила (кролики, 15 мг/кг массы тела) и в парах эфира (мыши). Экспериментальный протокол был одобрен комиссией по биологической этике Института физиологии Коми НЦ УрО РАН. Сердце извлекали и помещали в растворе Тироде следующего состава (мМ/л): 140 NaCl; 10 NaHCO 3 ; 5,4 KCl; 1,8 CaCl 2 ; 1 MgSO 4 ; 0,33 Na 2 HPO 4 , 10 глюкоза; 5 HЕРЕS (pH 7,4). Удаляли желудочки и предсердия. В результате препарат имел размер 3х3 мм и включал область синусно-предсердного узла, фрагмент правого предсердия с поперечным гребешком ( crista terminalis ) и сегменты верхней и нижней полых вен ( superior и inferior vena cava ). Препарат помещали в проточную, аэрируемую, термостатируемую камеру, заполненную раствором Тироде при температуре 31±1°С. Эффекты лидокаина регистрировали на протяжении 15 мин экспозиции.
Регистрацию внутриклеточных ПД проводили с помощью стандартной микроэлектродной техники. Обработку результатов выполняли с помощью программ Microsoft Office Excel и PowerGraph Professional версия 3.3 (Россия). Данные приведены как среднее арифметическое ± стандартное отклонение (М±σ). Достоверность различий определяли по U-критерию Манна-Уитни. Различия считали достоверными при 0,01< р <0,05.
Результаты исследований
Общая характеристика ПД клеток скрытого водителя ритма кролика и мыши. Синуснопредсердная область состоит из клеток, электрофизиологические параметры которых различаются в зависимости от места регистрации. В наших экспериментах ПД клеток были зарегистрированы вдоль артерии СП узла, ниже поперечного гребешка (рис. 1). Частота генерирования ПД у спонтанно сокращающихся препаратов была равна 108±22 имп / мин (n=6) и 316±40 имп/мин (n=20) у кролика и мыши соответственно. В контрольном растворе Тироде скорость фазы быстрой деполяризации у пейсмекерных клеток в этой области у кролика составила 9±3 В∕с и 16±5 В∕с у мыши. Место регистра- ции ПД и высокая скорость фазы быстрой деполяризации позволяют отнести эти клетки к клеткам, работающим в режиме скрытого водителя ритма.
Эффекты лидокаина на фазу быстрой деполяризации (фаза 0) ПД у кролика и мыши. Дозозависимый эффект лидокаина, блокатора Na+-каналов, исследован в диапазоне от 5 до 1000 мкМ (рис. 2). Установлено, что пороговая концентрация лидокаина для клеток, работающих в режиме скрытого водителя ритма, составила 100 мкМ (n=4) для кролика и 12 мкМ (n=4) для мыши. У обоих видов животных происходило замедление скорости нарастания фазы быстрой деполяризации (dV/dt max ) на 24% ( р <0,05) и 35% ( р <0,05) соответственно. При этом частота генерации ПД уменьшалась незначительно (у мыши на 13% ( р <0,05), у кролика не было отмечено достоверных изменений).
При дальнейшем повышении концентрации лидокаина происходило дозозависимое снижение dV/dt max у обоих видов животных (рис. 2). Анализ дозозависимой кривой показал, что концентрация лидокаина, при которой происходит снижение максимальной скорости деполяризации ПД в фазу 0 на 50% (ЕС 50 ) у клеток синусно-предсердного узла кролика, составила ≈230 мкМ, у мыши – ≈ 20 мкМ. При высоких концентрациях блокатора (1000 мкМ) у кролика наблюдается монотонное замедление dV/dt max , снижение амплитуды ПД, смещение максимального диастолического потенциала (E макс ) в сторону деполяризации и блок генерации ПД на 7-й мин экспозиции (n=5). У клеток СП узла мыши блок спонтанной активности происходил в течение 3 мин (n=3) после добавления лидокаина (1000 мкМ). При удалении блокатора из перфузирующего раствора спонтанная активность препаратов возобновлялась через 5–10 мин.
Лидокаин представляет собой селективный блокатор Na+-каналов. На нейронах крысы показано, что лидокаин (1000 мкМ) полностью блокирует Na+-каналы и генерацию ПД [9]. Однако ряд исследователей свидетельствуют, что кроме Na+-каналов местный анестетик действует на несколько типов Са2+- и К+-каналов [9]. В нейронах концентрация ли-
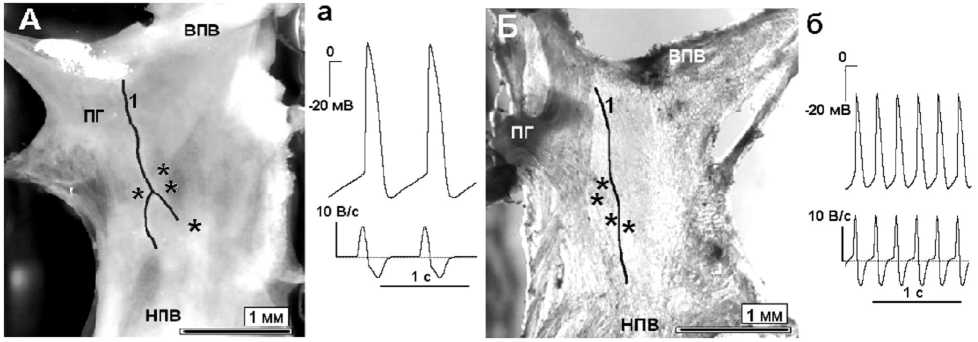
Рис. 1. Препараты синусно-предсердной области кролика (А) и мыши (Б) и потенциалы действия с первой производной (а, б). Примечание: ВПВ – верхняя полая вена; НПВ – нижняя полая вена; ПГ – поперечный гребешок; 1 – артерия синусно-предсердного узла; * – зона регистрации ПД типа скры-
того водителя ритма.
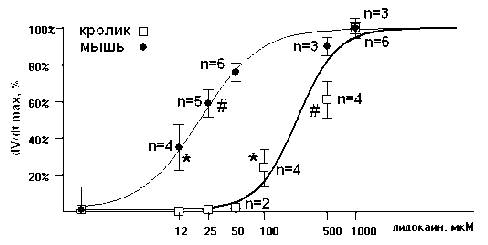
Рис. 2. Дозозависимый эффект лидокаина на скорость нарастания фазы быстрой деполяризации (dV/dt max ) ПД у кролика и мыши.
* – достоверность различий по сравнению с контролем ( р <0,05); # – достоверность различий по сравнению с * ( р <0,05).
докаина, которая блокирует Ca2+-каналы на 50% (ЕС 50 ), составляет ≈ 2,8 мМ [9]. Сродство К+-каналов к действию лидокаина зависит от типа самого канала и примерно в 10 – 80 раз ниже, чем для Nа+-канала [10]. Это в несколько раз больше, чем требуется для полной блокады Na+-каналов.
Таким образом, остановка генерации ПД в клетках СП узла кролика и мыши связана с полным блоком Na+-каналов. Однако стоит отметить, что чувствительность пейсмекерных клеток к низким концентрациям блокатора у этих животных различна. Возможно, что различие в чувствительности клеток к действию лидокаина (≈ в 10 раз) связано с разным соотношением сердечной (Na v 1.5) и нейрональной (Na v 1.1) изоформ Na+-каналов, бо́льшим вкладом быстрого компонента Na+-тока в генерацию пейсмекерной активности в СП узле мыши или в особенностях в строении ионных каналов (Na v 1.5 и Na v 1.1) на молекулярном уровне у этих животных.
Эффекты лидокаина на фазу медленной диастолической деполяризации (фаза 4) ПД у кролика и мыши. В низких концентрациях (100 мкМ для кролика и 12 мкМ для мыши) лидокаин не влиял на длительность и скорость фазы МДД. Повышение концентрации блокатора (500 мкМ для кролика, n=4; 25 мкМ для мыши, n=5) приводило к замедлению скорости фазы МДД (V 4 ) у мыши на 21% ( р <0,05). В клетках СП узла кролика V 4 достоверно не изменялась, но E макс смещался в сторону деполяризации (на 21 %). В результате частота генерации ПД снижалась на 23 и 27% у кролика и мыши соответственно (рис. 3).
В экспериментах на изолированных клетках СП узла кролика Rocchetti и соавт. [7] показали, что лидокаин может блокировать каналы HCN, по которым идет ток, активируемый гиперполяризацией (If), способствующий фазе МДД.
Наши результаты также демонстрируют изменение фазы МДД при действии лидокаина, однако механизм действия блокатора у исследуемых видов животных различен. В СП узле кролика заключительная часть МДД не изменялась, но за счет смещения E макс в сторону деполяризации существенно удлинялась фаза конечной реполяризации и незначительно снижалась спонтанная деполяризация. В СП узле мыши МДД существенно растягивалась в последней трети, а фаза конечной реполяризации не изменялась. Если эти изменения вызваны блоком HCN-каналов, тогда можно предположить, что свойства тока, активируемого гиперполяризацией (I f ), у этих видов животных различны и генерирование фазы МДД имеет свою специфичность.
Заключение
Лидокаин дозозависимо снижал максимальную скорость нарастания потенциалов действия в фазу 0 (dV/dt max ) у клеток синусно-предсердной области сердца кролика и мыши.
При сопоставимых dV/dt max у обоих видов животных пейсмекерные клетки синусно-предсердного узла мыши демонстрировали более высокую чувствительность (в 10 раз) к блокатору Na+-кана-лов лидокаину, чем клетки синусно-предсердного узла кролика.
Остановка спонтанной активности у пейсме-керных клеток кролика и мыши в присутствии лидокаина свидетельствует о том, что наравне с Ca2+-каналами Na+-каналы играют важную роль в формировании фазы быстрой деполяризации клеток синусно-предсердной области.
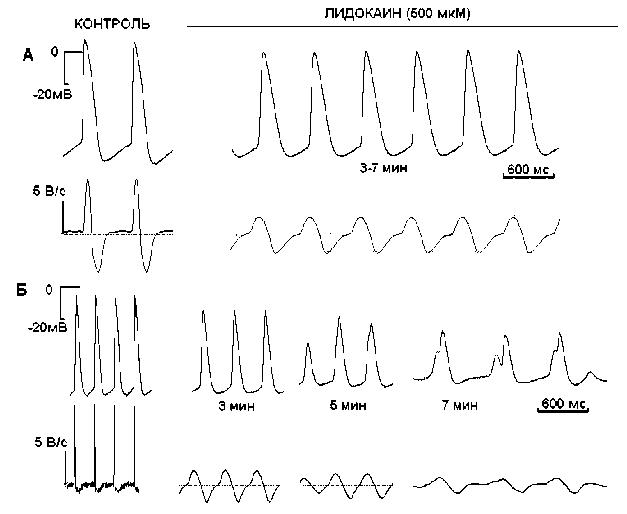
Рис. 3. Изменения конфигурации ПД и первой производной (dV/dt max ) кролика (А) и мыши (Б) при действии лидокаина (500 мкМ).
Список литературы Электрофизиологические параметры потенциалов действия клеток типа скрытого водителя ритма сердца кролика и мыши при действии лидокаина
- Baruscotti M., Robinson R.B. Electrophysiologyand pacemaker function of the developing sinoatrialnode//J. Physiol. Heart. Circ.Physiol. 2007. Vol. 293. P. Н2613-Н2623.
- Boyett M.R., Honjo H., Kodama I. The sinoatrialnode, a heterogeneous pacemakerstructure//Cardiovasc. Res. 2000. Vol. 47.P. 658-687.
- Lei M., Jones S.A., Liu T. et all. Requirementof neuronal-and cardiac-type sodium channelsfor murine sinoatrial node pacemaking//J.Physiol. 2004. Vol. 559. P. 835-848.
- Tellez J.O., Dobrzynski H., Greener I.D. et all.Differential expression of ion channel transcriptsin atrial muscle and sinoatrial node inrabbit//Circ. Res. 2006. Vol. 99. P. 1384-1393.
- Butters T.D, Aslanidi O.V., Inada S. et all.Mechanistic links between Na+ channel(SCN5A) mutations and impaired cardiacpacemaking in sick sinus syndrome//Circ.Res. 2010. Vol. 107. P. 126-137.
- Létienne R., Vié B., Le Grandet B. Pharmacologicalcharacterisation of sodium channels insinoatrial node pacemaking in the rat heart//Eur. J. Pharmacol. 2006. Vol. 530. P. 243-249.
- Rocchetti M., Armato A., Cavalieri B. et all.Lidocaine inhibition of the hyperpolarizationactivatedcurrent (If) in sinoatrial myocytes//J. Cardiovas. Pharmacol. 1999. Vol. 34. Р.434-439.
- Lebedeva E.А., Golovko V.А. An analysis of lidocaineand hyposodium solution effects ongeneration of pacemaker action potential inmouse sinoatrial node//Proc. XXX Ann.Meeting ISHR. Medimond. Bolonga. 2011.№628. P. 31-34.
- Gold M.S., Reichling D.B., Hampl K.F. et all.Lidocaine toxicity in primary afferent neuronsfrom the rat//J. Pharmacol. Exp. Ther. 1998Vol. 285. P. 413-421.
- Scholz A. Mechanisms of (local) anaesthetics onvoltage-gated sodium and other ion channels//Br. J. Anaesth. 2002. Vol. 89. P. 52-61.
