Элементы фрактальной теории видовой структуры гидробиоценозов

Автор: Гелашвили Д.Б., Иудин Д.И., Розенберг Г.С., Якимов В.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1 т.8, 2006 года.
Бесплатный доступ
В статье в тезисной форме представлены основные положения фрактальной теории видовой структуры сообществ (в первую очередь, видового богатства и видового разнообразия).
Короткий адрес: https://sciup.org/148197824
IDR: 148197824 | УДК: 547.2+51.001.572
Elements of fractal theories of the specific structure of hydrobiocenoses
Fundamentals of the fractal theories of the specific structure of communities (in the first place species resources and species diversity) are presented in the article.
Текст научной статьи Элементы фрактальной теории видовой структуры гидробиоценозов
1 Нижегородский государственный университет им.Н.И. Лобачевского,г. . Новгород 2 Институт экологии Волжского бассейна РАН,г.
В статье в тезисной форме представлены основные положения фрактальной теории видовой структу-ры сообществ (в первую очередь, видового богатства и видового разнообразия).
Физические условия и химический состав водной среды обуславливают специфические комбинации лимитирующих абиотических факторов, воздействующих на становление, развитие и функционирование гидробиоце-нозов . Гидробиоценозы являются сильно не-равновесными системами, связанными с пе-реносом интенсивных потоков вещества и энергии, и обладают имманентной структур-ной универсальностью, которую в настоящее время можно интерпретировать с позиции теории фракталов [1, 2, 4, 5, 8, 10]. Потоки энергии и вещества, проходящие через откры-тые системы, обеспечивают возникновение в них эффектов самоорганизации - образо-вание макроскопических диссипативных структур . Эти структуры очень часто демон-стрируют в широком диапазоне параметров пространственно-временной скейлинг -свойство самоподобия, или масштабной ин-вариантности - один из фундаментальных видов симметрий физического мира, играю-щих формообразующую роль во Вселенной. Пространственно-временной скейлинг ха-рактеризуется сильными, спадающими по сте-пенному закону, , которые ти-пичны для критических явлений . Поэтому ка-чественно динамика подобных систем про-изводит впечатление самоорганизованного и самонастраивающегося критического режима. Явления такого рода были объединены не-давно общим наименованием систем с само-организованной критичностью (self-organized criticality [17]). Самоорганизованная критич- ность соединяет в себе два направления со-временной физической парадигмы - самоор-ганизацию и критические явления - и актуа-лизирует новый подход к анализу сложного поведения нелинейных распределенных си-стем . Этот новый подход связывает динами-ку критических флуктуаций с появлением фракталов в конфигурационном пространстве нелинейной распределенной системы при кинетическом фазовом переходе.
Все это в полной мере относится к био-экологическим системам. Процесспроникно-вения теории фракталов в экологию можно условно разделить на три этапа. Первый из них был связан с необходимостью описания пространственной сложности тех или иных биотопов, в частности горных массивов, реч-ных систем, почвы, коралловых рифов и т. . На втором этапе пришел черед описания фрактального распределения отдельных ви-ДОВ. Наконец, на третьем этапе встал вопрос о самоподобии внутренней структуры самих сообществ.
В настоящей работе в тезисном виде пред-ставлены основные положения фрактальной теории видовой структуры сообществ (на примере гидробиоценозов) разрабатываемой в настоящее время авторами . В первом при-ближении ограничим рассмотрение видовой структуры анализом видового богатства (чис-ла видов), и относительными численностя-ми составляющих сообщество видов,т. . ви-довым разнообразием .
Видовое богатство
При анализе видового богатства ключевое значение имеет как его зависимость от пло-щади (species-area relationship, SAR), так и ха-рактер поведения кривой накопления видов в зависимости от объема выборки. История анализа SAR, имеющего, в основном, отно-шение к наземным местообитаниям и под-робно рассмотренная в нашей предыдущей работе [5], насчитывает уже около полутора столетий . Постановка вопроса о фрактальной структуре биотического сообщества является вполне логичной и в контексте дискуссии о видовом насыщении сообществ [3, 6 и др.]. Именно степенной закон, не предполагаю-щий насыщения функции f ( x ) , учитывает вклад редких видов, или видов с «единичной численностью» [20], не противоречит «пра-вилу экологической неаддитивности» Леви-ча [9] и, наконец, дает возможность приме-нить фрактальный формализм для характери-стики видовой структуры сообщества. Сле-дует отметить интересные перспективы, свя-занные с применением фрактального подхо-да к анализу таксономического разнообразия [13], а также «островных» сообществ, где в качестве «острова» могут выступать геогра-фические объекты, организмы или их части (лист - фитофаги - паразиты), системы орга-низмов, например, консорции, паразитарные системы, микробиоценозы семей обществен-пых насекомых и т. .
Специфика пространственного распреде-ления сообщества в водной среде заставляет обратиться к закономерности связи темпов накопления видов с увеличением объема выборки с позиции альфа-разнообразия Уит-текера [14]. Маргалеф [12], предложив сте-пенную зависимость числа видов ( S ) от сум-марной численности -
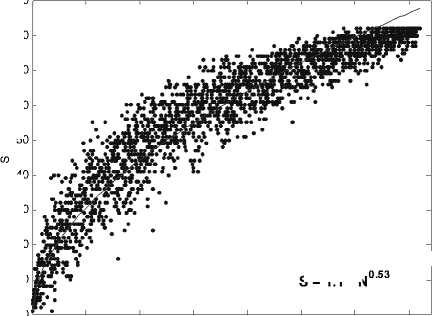
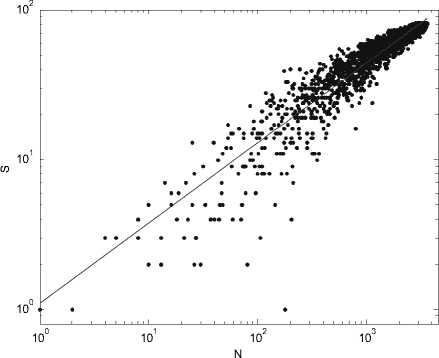
(N)S Nk, (1) тем самым, по-видимому, -нул фрактальную природу этой связи. Степей-ная зависимость в ортогональных координа-тах натуральных значений SиN легко преоб-разуется в линейную в билогарифмических координатах lnS k lnN , где k = lnS/lnN, 0 k 1 имеет смысл индек-са видового разнообразия Маргалефа. Выра-жение (1) показывает, сообщества (в данном случае в виде его ком-понента - видового богатства) инвариантна относительно преобразования его численно-сти . Величину k можно трактовать как соот-ветствующую фрактальную размерность: -ло элементов видовой структуры - число видов сообщества - меняется по степенному закону с показателем k при увеличении раз-меров системы,т. . с ростом численности сообщества. Таким образом, адекватным математическим образом накопления вида-вого богатства при росте выборочного уси-лия, являются монофракталы : , характеризующиеся единственной фрак-тальной размерностью.
Использование степенного закона для ана-лиза зависимости между видовым богат-ством выборки и ее размером начало полу-чать распространение относительно недав-но . Наличие фрактальной структуры для гид-робиоценозов разномасштабных водных объектов было показано в наших работах [8, 16]. Кроме того, наклоны степенной зави-симости (то есть фактически индекс Марга-лефа k ) часто обсуждаются в контексте влия-ния размера тела животных на структуру со-ответствующих сообществ [11, 18, 21].
На рис . 1 приведена экспериментальная зависимость числа видов макрозообентоса от его численности, полученная для ряда город-ских озер в Нижнем Новгороде . Видно, что точность определения показателя быстро увеличивается с ростом размера выборки. Необходимо подчеркнуть, что в отличие от регулярного математического фрактала для реального природного фрактала существует некоторый минимальный масштаб длины min , такой, что на расстояниях l » lmin его основ-
.
Кроме того, на достаточно больших масшта-бах длин l > lmax, где l max -
, самоподобия также нарушается . Поэтому свойства природных фракталов рассматрива-ются лишь на масштабах l, удовлетворяющих соотношению min max. В рассматри-
S = 1.1 * N
1000 1500 2000 2500 3000 3500
N
.1. .() ()
ваемом контексте фрактальности видового богатства сообщества эквивалентом масшта-ба длины является суммарная численность ( N ), что позволяет сформулировать следующее обобщение: фрактальная, структура -о-го объекта проявляется в асимптотике -
(N). - рой критической N « Nшп его осиовное свойство - самоподобие - нарушается.
Видовое разнообразие
Мы видим, таким образом, что показатель Маргалефа (k) тоответствует фрактальной размерности (D) три описании регулярных фракталов . Однако сама по себе фрактальная размерность не дает исчерпывающего количе-ственного представления о структуре само -подобного объекта (мнтжества с одинаковой фрактальной размерностью могут иметь раз -личную структуру). В р тмках такого описания роли отдельных видов в формировании структуры сообщества нивелированы: зависимость фиксирует лишь факт наличия того или иного вида в выборке, итнорируя его представленность и степень доминирования. В реальности мы имеем дело с принципиаль-но неоднородным распределением особей выборки по видам реестра. Нтиболее общее описание внутреннего устройства самопо- добных объектов позволяет дать теория муль-тифракталов, характеризуемых бесконечной иерархией размерностей [15]. Основные понятия , лежащие в основе того, что теперь принято называть мультифракталами, были в в едены Мандельбротом [10] в начале 70-х годов прошлого века.
Нами разработаны и обоснованы алгорит-мы анализа и интерпретации мультифрак -тальных спектров как обобщенного геомет -рического образа видовой структуры гидро -биоценозов, отражающего его основные компоненты : видовое богатство и видовое разнообразие [5, 8]. В эксперименте исследователь имеет дело с относительными частота -ми распределения особей по видам. Пусть набор {pi } характеризует относительные частоты распределения особей по видам: р = N/N , чде N - число особей i -го вида, N - размер пробной выборки, a i пробегает значения от единицы до полного числа видов S(N) , обнаруженных в пробе. Вектор p = (p1 ps) называется вектором относительной значи-мости видов. Сумма компонент этого вектора дает нам численность или размер выбор -ки, а количество ненулевых компонент дает число обнаруженных видов. Очевидно, что
Z м А^
Введем моменты распределения особей по видам и выясним характер их асимптотичес- кого поведения при увеличении численное-ти N:
n
Mq =z piq N т ( q ), (2)
i 1
где - ОО < q < оо - называется порядком момен-та, а показатель г( q ) характеризует скорость изменения соответствующего момента при увеличении размера выборки. Второе равен-ство в выражение (2) является обобщением формулы Маргалефа и, очевидно, совпадает с последней при q = 0 . Обобщенной размер-ностью Dq ( обобщенной размерностью Реньи ) распределения является убывающая функция q , вводимая определением [15]:
Dq
lim <
N
1 q
ln Mq ln N
£ ( q )
1 q ,
На практике величины (3) можно оценить, используя несколько различающихся значений N , по более простой формуле
ln M Dq q 1 q ln N
.
Для q=0 :
D k ln S ( N )
0 ln N
.
Можно показать, что для q = 1
i pi ln pi H
D 1 i 1 , где H есть ни
1 lnN lnN что иное как информационный индекс видо-вого разнообразия Шеннона. Для q=2
lnC 1
D2 - 1 или N 2, где C - индекс lnNC доминирования,а1/C 1 /у iS 1 pi 2
видового разнообразия Симпсона. Для q = ln ц
1/2: D1/2 ‘ ИЛИ и = N D12, где м — извес- lnN тный в экологической литературе индекс
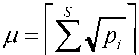
Заметим, i1
что для доли редких видов h 1- р) S [7], легко можно получить : h 1 N D 1 ' 2 D 0 . Нео -
жиданным может показаться отсутствие клас-сических аналогов индексов разнообразия для размерностей Реньи с отрицательными q . Вместе с тем значение таких индексов пред-ставляется чрезвычайно важным при выяв-лении роли редких видов . Рассмотрим, на-пример, обобщенную размерность Реньи для
-
1 ln M 1
q = -1 : D 1 , где наибольший вклад
-
2 ln N
в значение величины момента M-1, представ-ляющего собой сумму обратных частот, n1
именно редкие виды: M1-2 . Исполь-i 1 pi зуя соотношение (4) можно получить весь спектр обобщенных размерностей Dq q - .
достигается лишь в случае равно представ-ленности видов.
Таким образом, обобщенные размерности практически не зависят от численности со-общества и являются своеобразными струк-турными инвариантами однотипных систем отличающихся друг от друга размерами .В отличие от них, часто используемые индек-сы Симпсона ( C , 1/C ) и Шеннона ( H ) зависят и от численности, :
1/ C ND 2 SD 2 / D 0 и
H D 1 ln N D 1 ln S/ D 0 .
Следовательно, сами по себе они не мо-гут охарактеризовать разнообразия всего со-общества в целом и требуют дополнительного нормирования. Так, например, нормировав индекс Шеннона на его максимальное значе-ние для данного числа видов, получим ин-деке выравненности Пиелу [19], который уже не зависит от числа видов и инвариантен :
H D 1
E .
ln S D 0
В случае с индексом видового разнообра-зия Симпсона 1/C , нормирование его лога-рифма на логарифм числа видов дает новый индекс разнообразия у , который также явля-:
ln (1/ C L D 2 D 2
а =------=E .
ln S D 1 D 0
Обобщенную размерность D2 называют в теории фракталов корреляционной размерно-стью , поэтому индекс разнообразияу можно назвать корреляционным инвариантом. За-метим, что каноническая форма индекса вы-
равненности Симпсона
1 CS
инвариантом
:
1 CS
ND 2 S
N D 2 k . Непосред-
ственной проверкой легко убедиться в том, что каждый из всего бесконечного множества инвариантов вида Dq / D 0 представляет со -бой индекс выравненное™ и меняется в пре-делах от нуля (для хемостата) до единицы (в случае равнопредставленности видов). Заме-ТИМ, что индекс Маргалефа k (впрочем, каки все обобщенные размерности Dq) достигает единицы только при S = N . Показано, что известные в экологической литературе индек-сы разнообразия (Симпсона, Шеннона, Мар -галефа, Животовского), следует рассматри-вать как частные случаи обобщенных размер-ностей Реньи , лежащих в основе построения
мультифрактального спектра. Обоснована возможность рассмотрения биологического сообщества как мультифрактального объекта и характеристики его спектром размерностей . При таком подходе сообщество рассматрива-стоя как множество, состоящее из отдельных фрактальных подмножеств, которые можно интерпретировать как совокупности особей, относящихся к видам со сходной представ-ленностью. Для таких подмножеств можно вычислить фрактальную размерность Dq , ко -торая и характеризует видовое разнообразие в соответствующей группе видов . Математи-ческий аппарат фрактального формализма позволяет получить весь спектр обобщенных размерностей Dq q в интервале от
- . q
Характерный вид зависимости Dq(q) пред-ставлен на рис. 2 для четырех отборов проб макрозообентоса в вегетационном периоде 2000 г. , что функция Dq(q) является не
q возрастающей: D 1 D0 D1 D2 ... Ра- венство достигается лишь в случае равно представленности видов.
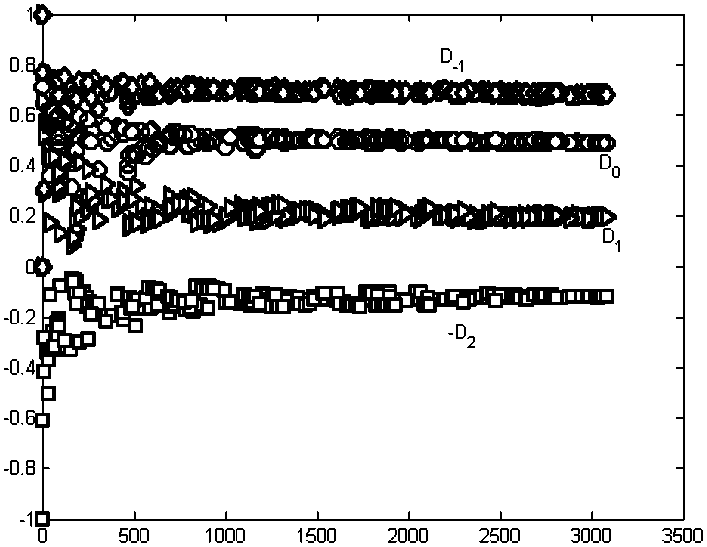
Рис. 2. Вычисление значений обобщенных размерностей Реньи по асимптотическому поведению моментов распределения особей макрозообентоса городских озер по видам при увеличении численности N пробных выборок. По оси абсцисс отложена численность выборок N , по оси ординат - значения моментов.
Моменты порядка: q = -1; О - q = 0; А - q = 1; D -q = 2
Таким образом, обобщенные размерности практически не зависят от численности со-общества и являются своеобразными струк-турными инвариантами однотипных систем отличающихся друг от друга размерами. Итак, обобщенные фрактальные размерно-сти, являющиеся инструментом мульти-фрактального анализа, отражают струк-турную гетерогенность сообщества, обус-ловленную различной представленностью, входящих в его состав видов.
Мультифрактальный формализм
Обсуждаемые выше обобщенные размер-ности Реньи не являются, строго говоря, фрактальными размерностями в общеприня-том понимании этого слова. Поэтому наряду с ними используется так называемая функция мультифрактального спектра f(a) .
Перейдем от переменных q и т( q ) к но-вым переменным с помощью преобразования :
a(q) = dd "Лq) dq .
f '(a(q )) = qa (q )+Д q)
Переменная a («индекс сингулярности») и неотрицательная функция f(a) («спектр син-гулярностей») дают представление о мульти-фрактальности структуры сообщества, полно-стью эквивалентное представлению через q и т( q ). На рис . 3 показана эволюция мульти-фрактальных спектров видового распределе-ния макрозообентоса городских озер Нижне-го Новгорода в период с мая по август 2000 . Экстремум спектра f max f(a(q 0)) = k со-впадает с индексом разнообразия Маргалефа и соответствует монофрактальной размерно-сти распределения. Правые ветви кривых на рис . 3 соответствуют отрицательным q . Они систематически короче левых. Такой вид спек-тров объясняется вкладом в сумму в выраже-НИИ (2) :
при больших отрицательных величинах по-рядка момента их вклад становится опреде-ляющим, подавляя влияние других элемен-товструктуры .
В случае равнопредставленности видов спектральная функция f(a) стягивается в точ-ку , f(a) = a .
Существует еще одна характерная точка d fa = 1 aH=a(q=1) кривой f(a): da f a 1, в ко -a aH торой прямая, проведенная через начало ко-ординат под углом 45° к положительному на-правлению оси a, касается кривой f(a). В этой точке значения индекса сингулярности и спектральной функции равны друг другу и совпадают с индексом D1 H /lnN:
f '(aH ) = aH ln N . На рис . 2и3 обсужда-емые точки отмечены треугольниками.
Индекс сингулярности a характеризует асимптотическое поведение подмножества видов с фрактальной размерностью f(a) при увеличении численности. , насколько быстро убывает удельное число представителей данного вида при увеличе-нии суммарной численности N: pi (N)х Nai.
Чем меньше ai, тем медленнее это убыва-ние и тем более представительный вид мы имеем . Размерность f(a) показывает, - го видов с данной сингулярностью ai содер-жится внутри всего сообщества. Иными ело-вами, f(a) есть размерность подмножества видов с сингулярностью a.
Таким образом, видовая структура биоти-ческих сообществ может быть количествен-но описана с помощью мультифрактального формализма. В свою очередь, график мульти-фрактального спектра (спектра сингулярно-сшей) есть геометрическое место точек, со -
-щепных фрактальных размерностей, вклю -чаюгцих, в качестве нормированных эквива-лентов, все известные показатели видовой структуры сообщества.
Мультифрактальный формализм представ-ляется серьезным инструментом при иссле-довании временнуй динамики видовой структуры биоценозов . Примером может слу-жить анализ зоопланктоценозов Чебоксарс-кого водохранилища. Характерными особен-
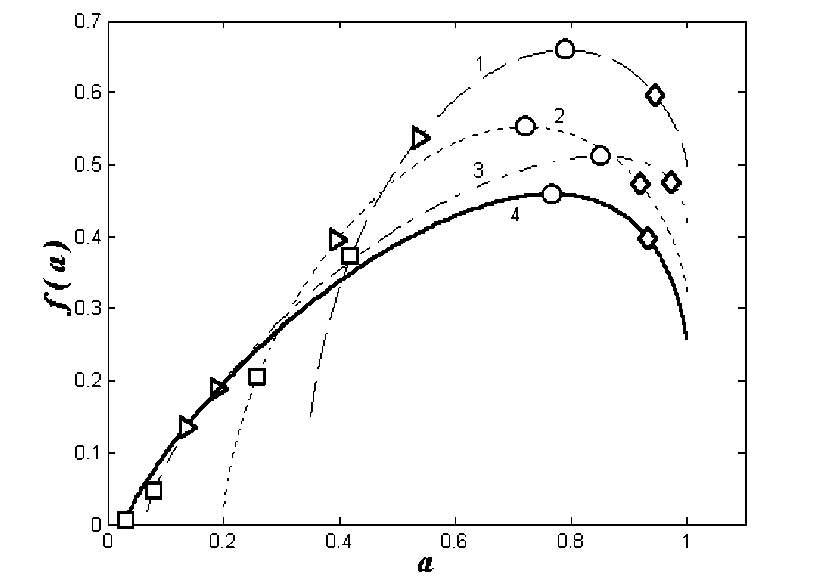
Рис. 3. Сезонная динамика мультифрактальных спектров видовой структуры сообщества макрозообентоса городских озер. По оси абсцисс: a - индекс сингулярности;
ординат: f(a) - спектр сингулярностей. :
1 - в мае, 2 - июне, 3 - июле, 4 - августе.
Список литературы Элементы фрактальной теории видовой структуры гидробиоценозов
- Азовский А.И., Чертопруд M.B. Анализ пространственной организации сообществ и фрактальная структура литорального бентоса//Докл. АН. 1997. Т. 356, № 5.
- Азовский A.И., Чертопруд M.B. Масштабно-ориентированный подход к анализу пространственной структуры сообществ//Журн. общ. биол. 1998. Т. 59.
- Бигон М., Харпер Дж., Таунсенд К. Экология: особи, популяции, сообщества: В 2-х т. М: Мир. 1989. Т. 2.
- Гелашвили Д.Б., Иудин Д.И., Розенберг Г.С. Фрактальная структура перколяционного кластера и пространственное распределение доминантных видов//Докл. АН. 2006. Т. 408, №4.
- Гелашвили Д.Б., Иудин Д.И., Розенберг Г.С., Якимов В.Н., Шурганова Г.В. Степенной закон и принцип самоподобия в описании видовой структуры сообществ//Поволжский экол. журн. 2004. № 3.
- Джиллер П. Структура сообществ и экологическая ниша. М.: Мир, 1988.
- Животовский Л.А. Меры популяционной изменчивости комплекса количественных признаков//Журн. общ. биол. 1980. Т. 41, №2.
- Иудин Д.И, Гелашвили Д.Б., Розенберг Г.С. Мультифрактальный анализ структуры биотических сообществ//Докл. АН. 2003. Т. 3 89, №2.
- Левин А.Л. Структура экологических сообществ. М.: Изд-во МГУ, 1980.
- Мандельброт Б. Фрактальная геометрия природы. М.: Ин-т компьютерных исслед., 2002.
- Мазей Ю.А. О видовом богатстве сообщества как функции объема выборки//Материалы VIII Всерос. попул. семинара. Н. Новгород: Изд-во Нижегород. ун-та, 2005.
- Маргалеф Р. Облик биосферы. М.: Наука, 1992.
- Поздняков А.А. Значение правила Виллиса для таксономии//Журн. общ. биол. 2005. Т. 66. № 4.
- Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980.
- Федер Е. Фракталы. М.: Мир, 1991.
- Шурганова Г.В., Иудин Д.И., Гелашвили Д.Б., Якимов В.Н. Мультифрактальный анализ видового разнообразия зоопланктоценозов Чебоксарского водохранилища//Актуальные проблемы рационального использования биологических ресурсов водохранилищ. Рыбинск: Изд-во Рыбинский дом печати, 2005.
- Bak P., Tang C., WiesenfieldK. Self-organized criticality//Phys. Rev. A. 1988. V. 38.
- Hillebrand H., Watermann F., Karez R., Berninger U.G. Differences in species richness patterns between unicellular and multicellular organisms//Oecologia. 2001. V. 126.
- Pielou, E.C. Shannon's formula as measure of species diversity: its use and misuse//Amer. Nat. 1966.V. 100.
- Rosenzweig M.L. On continental steady states of species diversity//Ecology of Species and Communities/Eds. Cody M.L., Diamond J.M. Cambridge (Mass.): Harvard Univ. Press, 1975.
- Siemann E., Tilman D., Haarstad J. Insect species diversity, abundance and body size relationships//Nature. 1996. V. 380.
