Эндогенная и временная изменчивость числа смоляных каналов в хвое деревьев сосны обыкновенной

Автор: Видякин Анатолий Иванович, Лебедев Алексей Геннадьевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 3-1 т.15, 2013 года.
Бесплатный доступ
Изучена эндогенная и временная изменчивость числа смоляных каналов и их аллометрических индексов в хвое деревьев сосны обыкновенной ( Pinus sylvestris L.). Установлены уровни изменчивости этих признаков. Показано, что аллометрические индексы по сравнению с числом смоляных каналов наиболее стабильны во времени.
Сосна обыкновенная, хвоя, смоляные каналы, аллометрические индексы, изменчивость
Короткий адрес: https://sciup.org/148201783
IDR: 148201783 | УДК: 574:574.2:582.475.4
Endogenous and temporal variability of the number of resin ducts in pine needles
Endogenous and temporal variability of the number of resin ducts and their allometric indices in pine (Pinus sylvestris L.) needles was studied. The levels of variability of these parameters are stated. It is shown that allometric indices are more stable in time, as compared with the number of resin ducts.
Текст научной статьи Эндогенная и временная изменчивость числа смоляных каналов в хвое деревьев сосны обыкновенной
Д альнейший прогресс в области микроэволю-ционного учения и эволюционно-генетической систематики древесных растений, а также разработка стратегии длительной эксплуатации и воспроизводства лесов возможны только при условии познания закономерностей формирования и организации популяционно-хорологической структуры вида.
За последние годы эта проблема наиболее успешно решается для сосны обыкновенной ( Pinus sylvestris L.) на основе разработанной системы методов выделения и географического анализа специфики изменчивости комплекса высокона-следуемых признаков генеративных органов [1]. Этими признаками-маркерами популяционноиерархической структуры вида являются фены и индексы шишек, семян, семенных крылышек, а также число семядолей [2, 3, 4, 5]. В результате феногеографических исследований, проведённых на востоке европейской части России, выявлены факторы и особенности формирования, организация популяционно-хорологической структуры P. sylvestris , а также составлены картосхемы расположения локальных популяций и их групп [1,6].
Основным объектом феногеографических исследований являются шишки, которые собираются со срубленных деревьев в насаждениях сосны, находящихся в точках пересечения субмеридиональных и широтных трансект, расположенных на расстоянии 100-150 км. При наличии урожая и действующих лесосек за осенне-зимний период (ноябрь-март) можно собрать шишки на большой части ареала вида. Однако, имеется ряд неблагоприятных факторов, значительно замедляющих выполнение этой работы. К ним относятся: 1) периодичность семеношения сосны, составляющая в зависимости от природной зоны 2-4 года; 2) непродолжительность периода возможной заготов-
ки шишек (около пяти месяцев); 3) отсутствие действующих лесосек в пунктах заготовки шишек.
Эти ограничения полностью устраняются при использовании в феногеографических исследованиях признаков-маркеров вегетативных органов P.sylvestris. Эти маркеры должны отличаться, прежде всего, высокой наследуемостью, простотой идентификации (качественные признаки) или количественной оценки.
Считается, что большинство признаков вегетативных органов сосны обыкновенной, по сравнению с генеративными, характеризуется более высокой эндогенной, временной, экологической изменчивостью и поэтому они обладают слабой информативностью в популяционных исследованиях и систематике [7, 8]. Одним из признаков этой группы, который наиболее часто и успешно применялся исследователями при изучении внутривидовой изменчивости, популяционной дифференциации и систематике P. sylvestris , является число смоляных каналов в хвое дерева [7, 8, 9, 10]. Однако, специальные исследования по изучению изменчивости числа смоляных каналов в кроне дерева, по годам формирования хвои, в результате которых возможна косвенная оценка степени наследуемости признака [1, 5], не проводились. В литературе нет также сведений о возможности индексной оценки признака. По аналогии с индексами генеративной сферы [1-3], относительные показатели числа смоляных каналов в феногеографических исследованиях вида могут оказаться эффективными.
На основании изложенного целью наших исследований является изучение эндогенной и временной изменчивости числа смоляных каналов в хвое деревьев сосны обыкновенной, оценка степени возможной информативности данного признака и вычисленных на его основе относительных показателей (индексов) в феногеографических исследованиях вида P. sylvestris .
МЕТОДИКА ИССЛЕДОВАНИЙ
В Быстрицком лесничестве Оричевского лесхоза Кировской области было отобрано 10 деревьев сосны обыкновенной естественного происхождения в возрасте 12 лет, растущих на открытой территории на расстоянии 30-35 м от материнского насаждения. В средней части центральных побегов 2009, 2010, 2011, 2012 годов каждого дерева собрано по 50 пар хвоинок. До начала анатомических исследований хвоя хранилась в этиловом спирте.
При изучении анатомо-морфологического строения хвои использовались методические рекомендации Л.Ф. Правдина [7] и С.А. Мамаева [8]. Для этого из каждой пары отбирали одну хвоинку. В средней части каждой хвоинки делали поперечный срез, на котором с помощью микроскопа МБС-1 определяли количество смоляных каналов с плоской и выпуклой стороны, их расположение (периферическое, паренхимное), измеряли ширину (ℓ) и толщину (h) хвоинки.
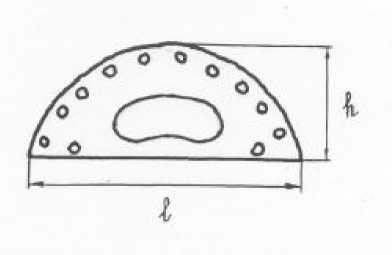
Рис. Схема измерений поперечного среза хвои сосны обыкновенной
Полученные данные по каждому количественному признаку обработаны статистически с помощью компьютерной программы Еxcel. В ре- зультате этого по четырём выборкам хвои (2009, 2010, 2011, 2012 гг.) каждого дерева были получены средние значения (М±m) и коэффициенты изменчивости(CV %): 1) общего числа смоляных каналов на срезе (N1); 2) числа смоляных каналов с выпуклой стороны хвоинки (N2); 3) ширины (ℓ) и толщины (h) хвоинки. Для каждой выборки хвои разных лет формирования вычислены четыре индекса числа смоляных каналов (ИЧСК): N1/ ℓ; N1/ ℓ + 2h; N2/ ℓ; N2/ ℓ + 2h. Применение величины ℓ + 2h в качестве одного из параметров индексной оценки обусловлено тем, что она приблизительно соответствует длине дуги, образующей выпуклую сторону поперечного сечения хвои (рис.), но в отличие от неё легко определяется с помощью мерной шкалы микроскопа. Для оценки уровней изменчивости признаков использована шкала С.А. Мамаева [8].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
Исследования показали, что ширина поперечного среза хвои разных лет формирования в пределах кроны дерева варьирует на очень низком уровне (CV < 7%), а высота – на очень низком и низком (CV = 7-12%) (табл.1). Форма поперечного сечения хвои каждого дерева, оцениваемая отношением (индексом) высоты среза к ширине, отмечается большой стабильностью по годам. Например, индекс формы поперечного сечения хвои дерева № 1, сформировавшейся в 2009 г. равен 0,43, 2010г. – 0,41, 2011г. – 0,43. в 2012 г. – 0,42, а дерева № 6 – соответственно 0,44, 0,42, 0,45, 0,42. При этом абсолютные средние значения ширины и толщины ежегодно формирующейся на дереве хвои варьируют в определённых пределах (табл.1), что вероятно обусловлено спецификой погодных условий.
Таблица 1. Изменчивость размеров поперечного сечения хвои 2009-20012 гг. формирования у отдельных деревьев сосны обыкновенной
|
№ дерева |
Пределы изменчивости |
|||
|
ширины сечения хвои |
высоты сечения хвои |
|||
|
М ± m, мм |
CV, % |
М ± m, мм |
CV, % |
|
|
1 |
1,56 ± 0,009 –1,97 ± 0,018 |
4,2-6,6 |
0,67 ± 0,007 – 0,83 ± 0,010 |
4,9-8,6 |
|
2 |
1,71 ±0,014 – 1,93 ± 0,011 |
4,3-5,6 |
0,79 ± 0,005 – 0,88 ± 0,009 |
4,5-6,9 |
|
3 |
1,54 ± 0,008 –1,89 ± 0,008 |
3,8-4,2 |
0,67 ± 0,009 – 0,85 ± 0,009 |
5,0-9,0 |
|
4 |
1,38 ± 0,008 –1,64 ± 0,020 |
4,0-8,4 |
0,62 ± 0,004 – 0,72 ± 0,007 |
4,7-11,3 |
|
5 |
1,53 ± 0,014 –1,66 ± 0,009 |
3,1-6,6 |
0,67 ± 0,007 – 0,70 ± 0,005 |
5,0-9,0 |
|
6 |
1,69 ± 0,016 –1,89 ± 0,010 |
3,8-6,7 |
0,75 ± 0,008 – 0,79 ± 0,006 |
5,7-7,6 |
|
7 |
1,45 ± 0,004 –1,70 ± 0,009 |
2,1-4,4 |
0,61 ± 0,005 – 0,71 ± 0,004 |
4,3-6,6 |
|
8 |
1,61 ± 0,007 –1,88 ± 0,009 |
3,1-5,1 |
0,68 ± 0,006 – 0,84 ± 0,006 |
5,2-6,2 |
|
9 |
1,55 ± 0,009 –1,74 ± 0,010 |
3,8-4,6 |
0,67 ± 0,004 – 0,73 ± 0,006 |
4,2-6,4 |
|
10 |
1,64 ± 0,009 –1,85 ± 0,018 |
3,1-7,0 |
0,69 ± 0,004 – 0,81 ± 0,008 |
4,2-7,4 |
В хвое всех выборок имеются периферические смоляные каналы, прилегающие к гиподерме, и паренхиматические, находящиеся в складчатой паренхиме мезофилла. Смоляные каналы распо- лагаются с выпуклой и плоской стороны хвои. При этом смоляные каналы, находящиеся с выпуклой стороны хвои, являются, в основном, периферическими, а с плоской – паренхиматическими. Наибольшее количество смоляных каналов расположено с выпуклой стороны хвои (табл.2).
Среднее число смоляных каналов в хвое изучаемых выборок изменяется в больших пределах. Например, в хвое 2009г. оно варьирует от 5,0 (дерево № 4) до 10,6 (дерево № 9). Данный признак изменяется также в пределах каждой выборки и по годам формирования хвои (табл. 2).
Эндогенная изменчивость числа каналов, оцениваемая коэффициентом вариации (CV, %), находится в пределах 8-19 %, что по шкале С.А. Мамаева [8] соответствует низкому и среднему уровню (табл. 2).
При этом следует отметить, что число каналов, расположенных с выпуклой стороны хвои, имеет меньшую эндогенную изменчивость, чем общее число их на поперечном срезе хвои. Этот признак варьирует в основном на низком уровне (CV = 712%) и только в редких случаях (например, в отдельных выборках хвои деревьев № 1, 4, 7, 8) – на среднем (CV= 13-20 %) (табл. 2).
Таблица 2. Изменчивость числа смоляных каналов в хвое 2009-2012 гг. формирования у деревьев сосны обыкновенной
|
№ дерева |
Общее количество смоляных каналов (верхняя строка) |
Изменчивость (CV,%) общего количества смоляных каналов (верхняя стр.) |
||||||
|
и в том числе с плоской стороны хвои (нижняя строка) |
и в том числе с плоской стороны хвои (нижняя строка) |
|||||||
|
по годам её формирования |
по годам её формирования |
|||||||
|
2009 |
2010 |
2011 |
2012 |
2009 |
2010 |
2011 |
2012 |
|
|
1 |
6,4±0,17 |
7,6±0,09 |
7,7±0,10 |
9,4±0,20 |
18,1 |
8,5 |
8,8 |
14,9 |
|
5,9±0,11 |
6,6±0,09 |
6,7±0,10 |
7,3±0,10 |
13,6 |
9,8 |
10,1 |
9,6 |
|
|
2 |
8,4±0,12 |
8,9±0,11 |
7,8±0,11 |
8,0±0,12 |
10,2 |
9,0 |
10,0 |
10,7 |
|
6,9±0,09 |
7,4±0,08 |
6,6±0,09 |
6,8±0,11 |
9,4 |
7,2 |
10,0 |
11,0 |
|
|
3 |
9,8±0,16 |
11,1±0,21 |
9,2±0,16 |
9,4±0,11 |
11,7 |
13,6 |
12,6 |
8,6 |
|
6,9±0,10 |
7,9±0,12 |
7,1±0,10 |
7,0±0,07 |
10,7 |
11,0 |
10,0 |
6,8 |
|
|
4 |
5,0±0,12 |
4,3±0,09 |
7,1±0,10 |
7,0±0,13 |
16,3 |
15,0 |
9,5 |
13,6 |
|
5,0±0,10 |
4,3±0,09 |
6,3±0,08 |
6,2±0,11 |
14,7 |
15,0 |
9,1 |
12,8 |
|
|
5 |
9,6±0,26 |
10,6±0,24 |
11,7±0,13 |
11,6±0,19 |
19,3 |
16,2 |
8,1 |
11,7 |
|
7,5±0,10 |
7,9±0,14 |
8,3±0,10 |
8,4±0,12 |
9,0 |
12,3 |
8,1 |
9,7 |
|
|
6 |
8,0±0,18 |
8,9±0,20 |
9,3±0,15 |
8,1±0,11 |
16,0 |
15,8 |
11,6 |
9,8 |
|
6,5±0,11 |
7,0±0,12 |
7,0±0,09 |
6,9±0,10 |
11,8 |
12,0 |
8,7 |
10,0 |
|
|
7 |
7,6±0,13 |
7,5±0,13 |
7,3±0,13 |
6,9±0,12 |
12,1 |
12,2 |
13,0 |
12,5 |
|
6,5±0,12 |
6,1±0,11 |
6,3±0,12 |
5,9±0,12 |
12,6 |
13,2 |
13,8 |
14,0 |
|
|
8 |
6,7±0,13 |
8,1±0,07 |
7,6±0,10 |
8,1±0,11 |
13,6 |
6,0 |
9,2 |
9,2 |
|
5,8±0,12 |
7,0±0,06 |
6,7±0,07 |
7,0±0,08 |
14,5 |
6,5 |
7,7 |
8,1 |
|
|
9 |
10,6±0,16 |
10,7±0,15 |
9,0±0,17 |
11,3±0,15 |
10,4 |
9,7 |
13,0 |
9,3 |
|
7,5±0,12 |
7,6±0,09 |
7,5±0,12 |
7,8±0,09 |
10,9 |
8,5 |
11,2 |
8,0 |
|
|
10 |
8,7±0,19 |
8,2±0,17 |
7,1±0,17 |
9,8±0,20 |
15,1 |
14,4 |
16,9 |
14,6 |
|
6,1±0,10 |
6,1±0,09 |
6,0±0,10 |
6,8±0,13 |
11,3 |
9,9 |
11,7 |
13,2 |
|
Таблица 3. Аллометрические индексы числа смоляных каналов в хвое деревьев сосны обыкновенной, сформировавшейся в 2009-2012 гг.
|
№ дерева |
Индексы числа смоляных каналов в хвое разных лет |
|||||||||||||||
|
N 1 / ℓ |
N 1 / ℓ +2 h |
N 2 / ℓ |
N 2 / ℓ + 2 h |
|||||||||||||
|
2009 |
2010 |
2011 |
2012 |
2009 |
2010 |
2011 |
2012 |
2009 |
2010 |
2011 |
2012 |
2009 |
2010 |
2011 |
2012 |
|
|
1 |
4,10 |
4,44 |
4,56 |
4,77 |
2,21 |
2,47 |
2,45 |
2,57 |
3,79 |
3,84 |
3,96 |
3,70 |
2,04 |
2,13 |
2,13 |
1,99 |
|
2 |
4,35 |
4,79 |
4,56 |
4,21 |
2,27 |
2,47 |
2,37 |
2,30 |
3,58 |
3,96 |
3,88 |
3,56 |
1,87 |
2,05 |
2,02 |
1,94 |
|
3 |
5,19 |
5,90 |
5,97 |
5,66 |
2,73 |
3,10 |
3,09 |
3,13 |
3,65 |
4,19 |
4,61 |
4,23 |
1,92 |
2,20 |
2,38 |
2,33 |
|
4 |
3,33 |
3,12 |
4,55 |
4,27 |
1,76 |
1,64 |
2,37 |
2,30 |
3,31 |
3,12 |
4,01 |
3,80 |
1,74 |
1,65 |
2,09 |
2,05 |
|
5 |
6,04 |
6,93 |
7,41 |
6,99 |
3,23 |
3,69 |
3,94 |
3,86 |
4,73 |
5,18 |
5,27 |
5,05 |
2,53 |
2,76 |
2,79 |
2,79 |
|
6 |
4,73 |
4,86 |
5,34 |
4,29 |
2,50 |
2,65 |
2,82 |
2,34 |
3,83 |
3,84 |
4,02 |
3,64 |
2,03 |
2,09 |
2,13 |
1,99 |
|
7 |
4,47 |
4,78 |
4,71 |
4,76 |
2,46 |
2,54 |
2,46 |
2,58 |
3,81 |
3,91 |
4,08 |
4,06 |
2,10 |
2,08 |
2,13 |
2,19 |
|
8 |
4,16 |
4,31 |
4,67 |
4,43 |
2,25 |
2,33 |
2,43 |
2,31 |
3,59 |
3,70 |
4,12 |
3,87 |
1,94 |
2,00 |
2,12 |
2,02 |
|
9 |
6,19 |
6,33 |
5,81 |
6,49 |
3,34 |
3,48 |
3,11 |
3,68 |
4,39 |
4,49 |
4,84 |
4,46 |
2,37 |
2,46 |
2,59 |
2,53 |
|
10 |
4,97 |
4,85 |
4,33 |
5,29 |
2,71 |
2,65 |
2,35 |
2,82 |
3,46 |
3,63 |
3,61 |
3,65 |
1,89 |
1,99 |
1,96 |
1,95 |
Таблица 4 . Показатели временной устойчивости числа смоляных каналов в хвое деревьев сосны о быкновенной и их индексов за период 2009-2012гг.
|
№ дерева |
Отношение минимального значения предела временной изменчивости признака к максимальному |
|||||
|
Общее число смоляных каналов на срезе хвои (N 1 ) |
Число смоляных каналов с выпуклой стороны хвои (N2) |
Индексы |
||||
|
N 1 / ℓ |
N 1 / ℓ +2 h |
N 2 / ℓ |
N 2 / ℓ + 2 h |
|||
|
1 |
0,68 |
0,81 |
0,86 |
0,86 |
0,93 |
0,93 |
|
2 |
0,88 |
0,90 |
0,88 |
0,88 |
0,90 |
0,91 |
|
3 |
0,83 |
0,88 |
0,87 |
0,87 |
0,80 |
0,81 |
|
4 |
0,61 |
0,69 |
0,69 |
0,69 |
0,78 |
0,79 |
|
5 |
0,82 |
0,90 |
0,81 |
0,82 |
0,89 |
0,91 |
|
6 |
0,86 |
0,92 |
0,80 |
0,82 |
0,90 |
0,94 |
|
7 |
0,91 |
0,91 |
0,94 |
0,95 |
0,93 |
0,95 |
|
8 |
0,83 |
0,83 |
0,89 |
0,92 |
0,88 |
0,92 |
|
9 |
0,80 |
0,97 |
0,89 |
0,86 |
0,91 |
0,92 |
|
10 |
0,72 |
0,88 |
0,81 |
0,83 |
0,95 |
0,95 |
Среднее значение 0,79 0,87 0,84 0,85 0,89 0,90
Общее число смоляных каналов на срезе (N 1 ) и число их с выпуклой стороны хвои (N 2 ) у каждого дерева изменяются по годам (табл. 2). Однако второй признак по сравнению с первым имеет примерно в два раза меньшие пределы временной изменчивости. Например, у дерева № 1 значение признака N 1 варьирует в пределах 6,4 – 9,4, а N 2 - 5,9 – 7,3, у дерева № 4 соответственно 4,3 – 7,1 и 4,3 – 6,3, у дерева № 5 – 9,6 – 11,7 и 7,5 – 8,4.
Аллометрические признаки, показывающие соотношение числа смоляных каналов (N 1 и N 2 ) и параметров поперечного сечения хвои (ℓ и ℓ + 2h), также варьируют по годам (табл. 3).
Для оценки стабильности числа смоляных каналов и аллометрических индексов во времени нами используется отношение нижнего предела временной изменчивости признака к верхнему. При этом, чем ближе значение показателя к 1, тем меньше временная изменчивость признака.
Расчёты показали, что изучаемые счётные признаки и аллометрические индексы отличаются по величине этого показателя (табл. 4).
Следовательно, они характеризуются различной временной изменчивостью. Наибольшие значения оценочного показателя имеют число смоляных каналов с выпуклой стороны хвои (N 2 ), а из аллометрических индексов – отношение N 2 / ℓ + 2h. Поэтому они отличаются от других изучаемых признаков наиболее высокой временной стабильностью. Это может, вероятно, являться следствием того, что их развитие жёстко контролируется генотипом дерева.
ВЫВОДЫ
-
1. Ширина и высота поперечного среза хвои отличаются очень низким уровнем эндогенной изменчивости, как правило, не превышающим 7 %. Абсолютные значения этих признаков изменяются во времени, что, вероятно, обусловлено
-
2. Эндогенная изменчивость числа смоляных каналов в хвое деревьев составляет 8-19 %, что соответствует низкому (CV = 7-12 %) и среднему (CV = 13-20 %) уровням [8]. В годы с благоприятными погодными условиями количество смоляных каналов в хвое дерева увеличивается, в основном, за счёт образования их с плоской стороны хвои. Число смоляных каналов с выпуклой стороны хвои по сравнению с общим количеством их отличается меньшей эндогенной и временной изменчивостью.
-
3. Аллометрические индексы имеют низкую эндогенную и временную изменчивость, что обусловлено, вероятно, жёсткой генотипической детерминированностью их развития.
-
4. Аллометрический индекс N 2 / ℓ + 2h, а также число смоляных каналов с выпуклой стороны хвои (N 2 ) возможно могут являться признаками-маркерами популяционно-хорологической структуры P. sylvestris.
спецификой погодных условий в год формирования хвои. Форма поперечного сечения хвои дерева, оцениваемая отношением высоты среза к ширине, отличается высокой временной стабильностью.
Работа выполнена при поддержке РФФИ (проект № 12-04-00062-а).
Список литературы Эндогенная и временная изменчивость числа смоляных каналов в хвое деревьев сосны обыкновенной
- Видякин А.И. Популяционная структура сосны обыкновенной на востоке европейской части России: Автореф. дис.. д-ра биол.наук. Екатеринбург, 2004. 48 с.
- Видякин А.И. Индексная оценка признаков популяционной структуры сосны обыкновенной//Лесоведение. 1991. №1. С. 57-62.
- Видякин А.И. Изменчивость формы апофизов шишек в популяциях сосны обыкновенной на востоке Европейской части России//Экология. 1995. №5. С. 356-362.
- Видякин А.И., Глотов Н.В. Изменчивость количества семядолей у сосны обыкновенной на востоке Европейской части России//Экология. 1999. №3. С. 170-176.
- Видякин А.И. Методические основы выделения фенов лесных древесных растений (на примере сосны обыкновенной (Pinus sylvestris L.). Сыктывкар, 2010. 28с. (Новые научные методики и информационные технологии/Коми научный центр УрО РАН, вып. 65.)
- Видякин А.И. Феногеография как основа познания закономерностей формирования и организации популяционно-хорологической структуры древесных растений (на примере Pinus sylvestris L. северо-востока Русской равнины)//Биологическое разнообразие растительного мира Урала и сопредельных территорий: Матер. всерос. конф. с междунар. участ. Екатеринбург, 2012. С. 150-152.
- Правдин Л.Ф. Сосна обыкновенная. Изменчивость. Внутривидовая систематика и селекция. М.: Наука, 1964. 190 с.
- Мамаев С.А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале). М.: Наука, 1973. 283с.
- Видякин А.И. Влияние географического происхождения семян на рост сосны обыкновенной в таёжной зоне Кировской области: Автореф. дис. … канд. с.-х. наук. Минск, 1978. 20 с.
- Санников С.Н., Петрова И.В. Дифференциация популяций сосны обыкновенной. Екатеринбург: УрО РАН, 2003. 248 с.
