Эндогенные (биоморфологические свойства растений) и экзогенные (природные и антропогенные) факторы в организации и динамике растительных сообществ (на примере пойменных лугов средней Оки)
 и экзогенные (природные и антропогенные) факторы в организации и динамике растительных сообществ (на примере пойменных лугов средней Оки)")
Автор: Егорова Валентина Николаевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
В данной статье рассмотрены динамика флоры и растительности пойменной экосистемы реки Оки и взаимосвязь влияния эндогенных и экзогенных факторов и механизмов в ходе природных и антропогенных сукцессий.
Пойменная экосистема, динамика, структура, ценопопуляции, оптимальные возрастные спектры, сукцессии
Короткий адрес: https://sciup.org/148200793
IDR: 148200793 | УДК: 581.
Endogenous (bio morphological properties of plants) and exogenous (natural and anthropogenic) factors in the organisation and dynamics of plant communities (on the case of floodland meadows of Oka)
This article examines the dynamics of the flora and vegetation of the flood land ecosystems of the River Oka and the relationship between the influence of endogenous and exogenous factors and mechanisms in the natural and anthropogenic successions.
Текст научной статьи Эндогенные (биоморфологические свойства растений) и экзогенные (природные и антропогенные) факторы в организации и динамике растительных сообществ (на примере пойменных лугов средней Оки)
венное соотношение в структуре сообществ. В этих условиях ни один вид, независимо от ЖФ и способов размножения (семенного, преимущественно вегетативного или смешанного), не достигает положения преимущественного доминанта и содоми-нанта, а виды в составе сообществ на том или ином уровне реализуют присущие им биоморфологиче-ские свойства и функционируют при разной численности ЦП. В результате формируются и стабильно функционируют полидоминантные, сменодоминантные пойменные растительные сообщества с богатым флористическим (28-49 видов на 100 кв.м. на разных частях поймы) составом. Количественная перегруппировка обилия видов, обусловливающая сменодоминантность пойменных сообществ, имеет флуктуационный характер и связана в пойменных местообитаниях главным образом с режимами поемности и аллювиальности.
В условиях интенсивного антропогенного пресса динамика растительных сообществ сопровождается количественными и качественными изменениями основных параметров их структуры. С 1960 -1963 гг. по 1997– 2010 гг., существенно сократилось число видов в различных частях поймы. В прирусловой части поймы: – а) при всех способах использования растительности в 1,2 раза, б) при внесении высоких доз минеральных удобрений и пастбищном использовании растительности в 2,5 раза, в) при внесении высоких доз минеральных удобрений и сенокосном использовании растительности в 1,9 раза. Соответственно – в переходной части поймы – в 1.2, 2,5, 1, 8 раза; в центральной части поймы – в 1,2, 2, 5, 1, 4 раза; в притеррасной части поймы – в 2,3, 4,3, 3,0 раза.
В структуре пойменных ценозов существенно снизилась фитоценотическая роль большинства видов, сохранившихся во флоре к 1997 – 2010 гг. В различных частях поймы в качестве доминантов функционировало 4 вида (в 6.3 раза меньше, по сравнению с 1940 – 1960 гг.), содоминантов – 12 видов (в 3 раза меньше, по сравнению с 1940 – 1960 гг.). Доминанты характеризовались длиннокорневищной (2 вида), длиннокорневищной-плотно-дерновенной (1), рыхлокустовой (1) ЖФ; содоми-нанты – длиннокорневищной (1 вид), рыхлокусто- вой (5), плотнодерновинной (1), длиннокорневищ-ной-плотнодерновинной (2), длиннокорневищной-рыхлокустовой (1), ползучей (1) ЖФ. Относительная доля доминантов составляла 62,7 – 85,5 % от общей биомассы, содоминантов – 9,2 – 9,7 %, относительная доля остальных видов в ряду сообществ не превышала 13.0 – 20.0 %. Флористическая насыщенность сократилась до 14 – 29 видов на100 кв.м. Выпадают или снижают численность до критического состояния виды не зависимо от ЖФ, способа возобновления и самоподдержания ЦП. Фито-ценотическая обстановка, формирующаяся в ходе антропогенных сукцессий, выступает элиминирующим факторов, ограничивая для многих видов возможность реализации присущих им биоморфо-логических свойств. Динамика флоры и растительности приобретают однонаправленный и необратимый характер.
С целью выявления соотношения влияния эндогенных и экзогенных факторов на динамику природных сообществ, факторов и механизмов их ес-тесственных и антропогенных сукцессий были про- анализированы биоморфологические свойства злаков основных ценозообразователей пойменных сообществ и уровень их реализации в различных условиях. Фитоценотическая роль изученных 11 видов злаков в эколого-антропогенных рядах менялась от доминантов до сопутствующих видов, онтогенетическое развитие особей видов осуществлялось при разном уровне их жизненного состояния. Во всех изученных сообществах, независимо от уровня жизненного состояния и численности ЦП, особи видов полностью завершали онтогенетическое развитие, хотя и при разной продолжительности большого жизненного цикла и длительности жизни побегов. Изученные виды характеризовались ЖФ: рыхлокустовой (Agrostis gigantea, Alopoecurus pratensis, Dactylis glomerata, Festuca pratensis , Phleum pratense, Poa palustris); длинно-корневищно-рыхлокустовой (Poa trivialis); длинно-корневищно-плотнокустовой (Festuca rubra, Poa poratensis ); длиннокорневищной (Bromopsis iner-mis, Elytrigia repens).
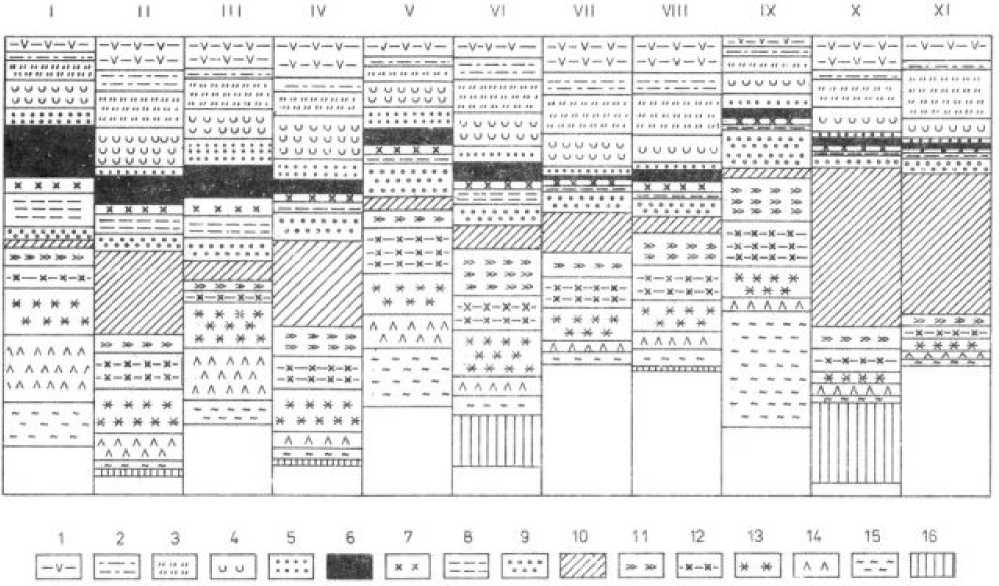
Рис. 1. Соотношение количественных и качественных характеристик признаков и свойств особей, ценопопуля-ций, консорций злаков. Условные обозначения: 1 - продолжительность большого жизненного цикла особей; 2 -продолжительность онтогенеза побегов; 3 - размер дерновин или число парциальных кустов; 4 -общее число побегов на особь; 5 - % генеративных; 6 – потенциальная семенная продуктивность; 7 – реальная семенная продуктивность; 8 – условно-реальная семенная продуктивность; 9 - % плодообразования; 10 – абсолютный вес семян; 11 – время созревания семян; 12 – энергичность осыпания; 13 – способ размножения; 14 – число семян на 0.25 м2; 15 – число проростков на 0.25 м2; 16 – число личинок галлиц на генеративный побег.
Масштаб. Продолжительность онтогенеза: особей – 2 года – 1мм; побегов – 1 год – 1мм; размер растения: 1см – 1 мм, 1 парциальный куст – 1 мм; число побегов: 30 побегов - 1см; % генеративных: 10 % - 1 мм; семенная продуктивность для всех элементов: 100 шт. – 1мм; время созревания семян, энергичность их осыпания и способ размножения: 1 балл – 2 мм; число личинок галлиц на генеративный побег: 50 шт – 1 см.
I – Agrostis gigantea ; II – Dactylis glomerata ; III – Phleum pratense ; IV – Festuca pratensis ; V – Poa palustris ; VI – Alopecurus pratensis L .; VII – Festuca rubra ; VIII – Poa pratensis ; IX – Poa trivialis ; X – Bromus inermis, XI – Agropyron repens.
Виды разных ЖФ характеризовались способами размножения - семенным (рыхлокустовые злаки ), смешанным (семенным и вегетативным - длинно-корневищно-рыхло- плотнокустовые злаки), преимущественно вегетативным (длиннокорневищные злаки). Для характеристики семенного размножения изучали потенциальную, условно-реальную, реальную семенную продуктивность, плодообразо-вание, потенциальный запас семян ЦП видов, динамику проростков в сообществах. Для выяснения роли насекомых в семенном возобновлении злаков были проведены исследования структуры консорций их генеративной сферы. Были получены качественные и количественные характеристики комплекса фитоценотически значимых биоморфологи-ческих свойств особей, ЦП, консорций, которые позволили рассмотреть в определенной взаимосвязи и взаимообусловленности закономерности их формирования. Во всех случаях использовали средние данные за период исследования, полученные в сообществах, в которых каждый из анализируемых видов функционировал как доминант или содоминант. Количественные и качественные характеристики особей приведены для их средневозрастного генеративного состояния. Растения в этом возрастном состоянии наиболее полно проявляют присущие им свойства, имеют законченное структурное выражение и, таким образом, наиболее полно отражают наследственные свойства.
Сравнительный анализ показал, что у каждого вида в процессе эволюционного становления био-морфологические свойства формируются во взаимосвязи и взаимообусловленности. Для каждого вида был выявлен свой индивидуальный коррелятивный ряд комплекса биоморфологических свойств (рис. 1). Было установлено, что уровень репродуктивной способности растений непосредственно и прямолинейно не связан со способом размножения, хотя некоторые тенденции в этом отношении и наблюдаются. Структура консорций также в определенной степени взаимосвязана с биомор-фологическими свойствами растений. Это дает основание считать, что коадаптация растений и их фитофагов осуществляется в процессе сопряженной эволюции и обеспечивает функционирование растений и животных в природных биоценозах. Эволюционное становление видов, коадаптация растений и фитофагов, формирование биоценозов идут параллельно [3 и др.].
Полученные материалы позволили определить общие и специфические черты функционирования ЦП видов и рассмотреть представление об оптимальных возрастных спектрах, структура которых обусловлена биоморфологическими свойствами растений, сформированными в процессе эволюционного становления (эндогенными факторами). Биоценоз в этой ситуации выступает как среда, обеспечивающая оптимальную (наиболее полную) реализацию присущих растениям биоморфологиче- ских свойств. Равновесное состояние между пополнением ЦП в процессе размножения и отмиранием особей в конце онтогенеза направлено на стабильность их функционирования. В этом случае отсутствует нарушение непрерывности самопод-держания ЦП, закономерного, характерного для каждого вида, хода онтогенетического развития, что обеспечивает в структуре ЦП определенный уровень, обусловленный свойствами вида, участия особей каждого возрастного состояния и предупреждает их скопление в каком-либо возрастном состоянии. В данной ситуации организация ЦП видов будет определяться преимущественно био-морфологическими (внутренними, эндогенными) свойствами растений, сформированными в процессе эволюционного становления, а, следовательно, будет наиболее благоприятной, оптимальной.
В этой связи для каждого вида оптимальным мы рассматриваем такой возрастной спектр ЦП, когда в основу определения относительной доли особей каждого возрастного состояния положены биомор-фологические свойства, достаточно полная реализация которых в ценозах обеспечивает равновесное состояние между поступлением молодых растений, развившихся из диаспор и отмиранием в результате старения, а также характерную для каждой биоморфы последовательность перехода особей из одного возрастного состояния в следующее с учетом элиминации в тех возрастных состояниях (проростки, ювенильные растения), в которых активный процесс отмирания наблюдается не только в критических ситуациях для развития растений. При расчете оптимального возрастного спектра относи -тельная доля каждого возрастного состояния определяется следующей зависимостью: P = t/T или P = n.t/T, где Р - относительная доля каждого возрастного состояния в оптимальном возрастном спектре; Т - продолжительность большого жизненного цикла; t - продолжительность возрастного состояния; n - число партикул, возникающие в процессе деградации семенной особи или число особей клона каждого возрастного состояния. Первая зависимость применима для биоморф, у которых в течение всего онтогенеза либо отсутствует партикуля-ция семенных растений, либо она не приводит к их фитоценотической разобщенности. В этом случае при ценопопуляционных исследованиях в качестве счетной единицы используется особь, развившаяся из зародыша семени. При определении оптимального возрастного спектра для биоморф, у которых с процессом партикуляции и вегетативного размножения связано формирование новых, в разной степени омоложенных, физически и фитоценотически обособленных структур применима вторая зависимость. Эти структуры (омоложенные партикулы, особи клона и др.) как и особи семенного происхождения выступают в качестве единиц счета при ценопопуляционных исследованиях в природных сообществах. В наших исследованиях мы рассчи- тали оптимальные возрастные спектры для 11 видов злаков. При их расчете использовали данные продолжительности пребывания особей в каждом возрастном состоянии, полученные в ценозах, где они занимают доминирующее положение, характеризуются хорошей жизненностью, осуществляют закономерный для каждого вида переход из одного возрастного состояния в следующее, полностью завершают онтогенетическое развитие. При расчете эмпирических возрастных спектров видов использованы фитоценотические параметры (число осо- бей каждого возрастного состояния на единицу площади), которые получены в пойменных сообществах, где фитоценотическое положение каждого вида менялось от доминирующего до сопутствующего (рис. 2).
Сопоставление оптимальных и эмпирических возрастных спектров достаточно четко демонстрирует уровень реализации биоморфологических свойств растений в различной эколого-фитоценотической обстановке.
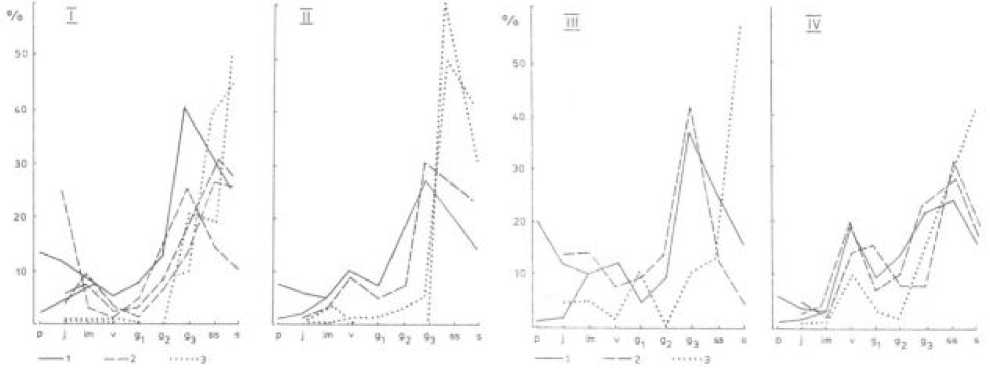
Рис. 2 . Соотношение эмпирических и оптимальных возрастных спектров злаков, характеризующихся различными жизненными формами. 1- оптимальные возрастные спектры, 2 – эмпирические возрастные спектры, полученные в ценозах, где виды входят в группу доминантов или содоминантов; 3 – эмпирические возрастные спектры, полученные в ценозах, где виды входят в сопутствующую группу.
I - Dactylis glomerata L ., II - Bromus inermis Leyss ., III - Poa trvialis L. , IV - Poa pratensis L.
Список литературы Эндогенные (биоморфологические свойства растений) и экзогенные (природные и антропогенные) факторы в организации и динамике растительных сообществ (на примере пойменных лугов средней Оки)
- Егорова В. Н. Динамика видового состава и спектров жизненных форм флоры поймы реки Оки в ходе естественных и антропогенных сукцессий//Бот. журн. 2004. Т. 89, N 6. С. 957-973.
- Егорова В. Н. Динамика видового состава и таксономической структуры флоры поймы реки Оки (Дединовское расширение) в ходе естественных и антропогенных сукцессий//Бот. журн. 2007. Т. 92, № 5. С. 702-722.
- Егорова В. Н., Мамаева Х. П., Фирсов С. Н. Структура консорций генеративной сферы злаков и возобновление их ценопопуляций в пойменных ценозах реки Оки//Бот. журн. 2001. Т. 86, № 7. С. 26-38.
