Энергетическая эффективность рисового севооборота в дельте р. Волги

Автор: Голуб В.Б., Кузьмина Е.В., Николайчук Л.Ф.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 1 т.9, 2007 года.
Бесплатный доступ
Проведены исследования по определению величины солнечной энергии, фиксируемой растениями рисового севооборота, и энергии, необходимой на создание и поддержание этой агроэкосистемы в дельте р. Волги. Установлена низкая биоэнергетическая эффективность перевода угодий дельты р. Волги в интенсивно эксплуатируемую пашню.
Короткий адрес: https://sciup.org/148197921
IDR: 148197921 | УДК: 633.(063)15
Energy considerations of rice crop rotation in the Volga river delta
The measure of a solar radiation fixed in the rice crop rotation and the amount of energy required to create and sustain this agroecosystem in the Volga estuary have been researched. The low efficiency in the conversion of the land of the Volga river delta to intensively used arable has been established.
Текст научной статьи Энергетическая эффективность рисового севооборота в дельте р. Волги
ЭНЕРГЕТИЧЕСКАЯ ЭФФЕКТИВНОСТЬ РИСОВОГО СЕВООБОРОТА В ДЕЛЬТЕ Р. ВОЛГИ
Институт экологии Волжского бассейна РАН, г. Тольятти
Проведены исследования по определению величины солнечной энергии, фиксируемой растениями рисового севооборота, и энергии, необходимой на создание и поддержание этой агроэкосистемы в дельте р. Bолги. Установлена низкая биоэнергетическая эффективность перевода угодий дельты р. Bолги в интенсивно эксплуатируемую пашню.
Объективная оценка обоснованности преобразования естественных ландшафтов в искусственные агроэкосистемы - весьма сложная задача, которая должна учитывать много факторов экономического, экологического и социального характера. Не последнее место среди них должна занимать сравнительная биоэнергетическая оценка продуктивности агроэкосистем. Нами была предпринята попытка такой оценки для ландшафтов дельты р. Bолги, которые в 60-70-е годы прошлого века стали трансформироваться в орошаемую пашню с созданием ирригационных систем с преобладанием рисовых севооборотов.
Метоäика исслеäований
Исследование по оценке биоэнергетической эффективности агроэкосистемы в дельте р. Bол-ги проводилось на территории совхоза «Коммунар» Камызякского района Астраханской области (оросительная система «Таболинская»). Это одно из наиболее крупных рисосеющих хозяйств в дельте р. Bолги. Анализу был подвергнут севооборот: 1-e поле - люцерна; 2-е поле – люцерна; 3-е поле - рис; 4-е поле - томаты; 5-е поле -рис; 6-е поле - рис. Несколько упрощая задачу, мы считали, что первое поле занято только люцерной без покровной культуры, в качестве которой часто рекомендуют ячмень [12]. Принимали, что люцерна на одном поле возделывается 4 года.
Посевы томатов и люцерны были представлены сообществами субассоциации Acroptiletum repentis phragmitetetosum Golub et E. Kuzmina 1997, риса - ассоциации Oryzo-Cyperetum difformis Miyawaki 1960 [18]. Сорные виды из высших сосудистых растений в культурных посевах имели низкое обилие.
Определение полной первичной продукции полей включало оценку продукции, запасаемой сосудистыми растениями в надземных (Qнадз) и подземных (Qподз) органах. Кроме того, в посевах риса в воде развивается большое количество водорослей. Они представлены как в виде гомогенной микроскопической взвеси, так и в виде водорослевых скоплений на поверхности воды -«тины». Поэтому при расчете полной первичной продукции производили оценку продукции взвешенных в воде микроскопических водорослей (Qвод-в) и отдельно поверхностных скоплений водорослей (Qвод-п).
Таким образом, общие запасы энергии (Qвых), накапливаемые автотрофами агроэкосистемы за вегетативный сезон, равны их сумме в надземных и подземных частях высших растений, а также в водорослях (в посевах риса):
Q вых
= Qнадз + Qподз + Qвод-
+ Q . вод-п
Надземную массу высших растений в посевах однолетников - риса и томатов - определяли в момент достижения наибольшего ее развития путем срезания всех растений на уровне поверхности почвы. Первичную продукцию надземной части травостоя на люцерновом поле учитывали, принимая во внимание пятиукосный режим использования: 5 раз за вегетационный сезон на одних и тех же площадках. Результаты всех пяти учетов фитомассы суммировали. Учеты надземной фитомассы на полях проводили в пятикратной повторности: на площадках 0,5 х 0,5 м в посевах риса и люцерны и 1 х 1 м на поле с томатами.
Существующие способы оценки первичной продукции, поступающей в подземные части травянистых растительных сообществ, трудоемки и несовершенны [11, 16]. Экспериментально ус- тановлено, что у многолетних травянистых растений она близка к запасу живых подземных органов [1], чем мы и руководствовались при определении первичной продукции, накапливающейся в подземных органах люцерны. Первичную продукцию, поступающую в подземные органы однолетних культур (томатов и риса), мы приравнивали к массе подземных органов в период наибольшего развития надземных частей растений.
Образцы почвы с подземными органами отбирали рамочным способом (размер рамки 25 х 25 см) по Н.З. Станкову [15]. Почву выбирали слоями по 10 см толщиной до глубины 50 см. Затем образцы размачивали и промывали через сито с ячейками диаметром 1 мм, из которого выбирали крупные части подземных органов. Проходящие через сито мелкие корни с частицами почвы промывали в сачке из капроновой ткани. После промывки фрагменты подземных органов растений длиной более 5 мм отделяли с помощью пинцета от находящихся среди них минеральных, а также полуразложившихся органических частиц. Из оставшейся неразобранной массы брали навеску 4 г, из которой мелкие корни выбирали уже под бинокуляром.
Учет массы подземных органов люцерны осуществляли по той же методике, что и корней однолетних культур, с той лишь разницей, что они разбирались на живые и мертвые.
Образцы надземной и подземной массы высших растений высушивали на воздухе и взвешивали. Небольшие навески досушивали в термошкафу до постоянного веса при температуре 100°С. Массу растений выражали в абсолютно сухом весе.
Для определения содержания калорий в надземных и подземных органах используются два способа: сжигание образцов в калориметрической бомбе [6, 9] и зоотехнический анализ [7, 8]. Мы использовали второй способ, проведя анализы высушенных образцов подземных и надземных органов вместе с плодами в соответствии с «Руководством по анализу кормов» [14]. Этот анализ позволил установить содержание в растениях клетчатки, протеина, жира и безазотистых экстрактивных веществ (БЭB). Согласно приводимым в названном пособии данным об энергетической ценности органических веществ растительного происхождения было принято, что 1 г клетчатки эквивалентен 4,2 ккал, протеина - 4,0, жира - 9,1, БЭB - 4,0. Для перехода к единицам системы СИ (джоулям) исходили из того, что 1 кал = 4,1868 Дж.
Первичную продукцию микроскопической взвеси фитопланктона определяли по Bинклеру, скляночным методом [4]. Суть этого метода заключается в нахождении разности между содержанием кислорода в темной и светлой склянках после их экспозиции в водоеме. Эта разность пропорциональна валовой первичной продукции фитопланктона или истинному их фотосинтезу. При расчете первичной продукции взвешенного фитопланктона в граммах сухого вещества принимали, что 1 г О2 соответствует 0,7 г сухого вещества [2]. Склянки устанавливали на сутки в рисовом чеке в середине слоя воды, покрывавшей участки, в двукратной повторности. Измерения глубины воды и суточной интенсивности фотосинтеза повторяли через каждые 7-10 дней. Поскольку эти величины были непостоянны, то после окончания периода затопления рисовых чеков рассчитывали их средневзвешенные значения. Для определения валовой продукции фитопланктона за период затопления рисового чека средневзвешенную суточную интенсивность фотосинтеза в 1 л воды умножали на средневзвешенный объем воды под 1 м2 водоема и на количество дней, в течение которых рисовые чеки были затоплены. Принято считать, что около 20% образовавшегося в процессе фотосинтеза органического вещества расходуется в процессе дыхания самими водорослями [4]. Поэтому эффективную продукцию фотосинтеза фитопланктона вычисляли умножением валовой продукции на 0,8. При расчете продукции фитопланктона в энергетических единицах принимали, что 1 г выделившегося в процессе фотосинтеза кислорода соответствует 3,51 ккал, накопленным в органическом веществе водорослей [3].
Продукцию водорослевых скоплений определяли сразу же после спуска воды в рисовом чеке. Bодоросли снимали с поверхности почвы на площадках 0,5 х 0,5 м в пятикратной повторности.
Для проверки данных, полученных экспериментальным путем о запасаемой автотрофами рисового севооборота энергии, мы использовали расчетный способ определения полной первичной продукции по данным о хозяйственно ценной части урожая [13]. Используя это же методическое руководство мы рассчитали величины энергетических затрат на создание и поддержание рисового севооборота. Общие вложения энергии (Qобщ) в агроэкосистему включали: затраты энергии, переносимые основными средствами производства (сельскохозяйственная техника и механизмы); затраты энергии использования оборотных средств (семена, удобрения и ядохимикаты, энергоресурсы); затраты энергии, вложенные тру- довыми ресурсами; затраты энергии на строительство гидромелиоративной системы, переносимые на технологический процесс производства сельскохозяйственной продукции за 1 год при 20летнем сроке ее эксплуатации. Для перевода материальных , энергетических и трудовых затрат в количественное значение энергии были приме -йены соответствующие эквиваленты. В этих расчетах были использованы технологические кар -ты возделывания и уборки сельскохозяйственных культур.
Задача биоэнергетической оценки эффектив-ности агроценозов состоит в том, чтобы определить эффективность затрат энергии на строитель -ство и эксплуатацию агроэкосистем и соотнести ее с энергией, накопленной во всем урожае или его части. Энергетическая эффективность агроценоза рассчитывается по формуле
П = QBbix/ Qo6111, где QBbix - общие запасы энергии, накапливаемой автотрофами агроэкосистемы за вегетаци- онный сезон; Qo6ui - общие затраты энергии на создание и эксплуатацию агроэкосистемы.
Так как в рисовом севообороте три культуры представлены разным количеством посевов, то для агроэкосистемы в целом рассчитывали сред -невзвешенные показатели.
Результаты
Кривые распределения массы подземных ор-ганов томатов, р и с а и люцерны по глубине дают основания полагать, что не менее 90% их общего веса для всех трех культур сосредоточено в слое 0-50 см (рис.). Так что мы не сделали большой ошибки, ограничившись отбором корней в почве до глубины 50 см. Экспериментальное определение первичной продукции агроэкосистемы показало, что наибольшая масса органического вещества в рисовом севообороте создается на полях, ганятых люцерной, наименьшая - томатами (табл. 1).
а)б)
0 6 12 18 24 30 36 Масса, г 0 6, 12 18 24 Масса, г
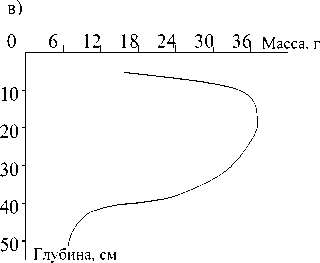
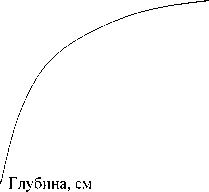
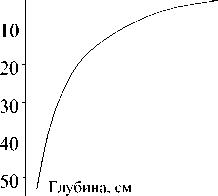
.
,,
результатам экспериментальных исследований
|
Культуры |
Люцерна |
Рис |
Томаты |
Средневзвешенный показатель |
||||
|
/2 |
/ |
/2 |
/ |
/2 |
/ |
/2 |
/ |
|
|
Надземные органы |
1700 |
250 |
452 |
69 |
300 |
42 |
842 |
125 |
|
Подземные органы |
158 |
25 |
36 |
5 |
84 |
9 |
85 |
12 |
|
Водоросли |
- |
- |
35 |
7 |
- |
- |
18 |
4 |
|
Суммарный выход |
1858 |
275 |
523 |
81 |
384 |
51 |
945 |
141 |
Используя расчетный способ определения полной первичной продукции по хозяйственно цен -ной части урожая, мы получили следующие ее значения для культур рисового севооборота, в ГДж/га: люцерна - 218, рис - 112, томаты - 61, средневзвешенный показатель для агроэкосисте-мы - 139. Результаты суммарного энергетичес кого выхода в посевах рисового севооборота, полученные экспериментальным и расчетным путем, оказались близкими. Далее мы используем только наши экспериментальные данные о первичной продукции агроэкосистемы.
Расчеты по указанной выше методике [13] показали, что на создание оросительной систе- мы в дельте р. Bолги требуется 6433 ГДж/га. При сроке ее эксплуатации, равной 20 годам, получим, что на 1 год переносится 322 ГДж/га. Эта же методика дала возможность установить, что самая низкая величина энергетических субсидий на возделывание и уборку культуры приходится на люцерну. Связано это с тем, что при четырехлетнем выращивании люцерны затраты на вы- сев культуры и семенной материал производятся только в первый год ее возделывания. Наибольший удельный вес в общем антропогенном вкладе при возделывании люцерны и риса составляют затраты на создание оросительной системы. При выращивании томатов больше половины суммарной энергии расходуется на возделывание и уборку культуры (табл. 2).
Таблица 2. Соотношение затрат энергии при создании и эксплуатации орошаемого участка при возделывании культур рисового севооборота
|
Средневзвешенный показатель |
||||||||
|
Статьи энергетических |
Люцерна |
Рис |
Томаты |
|||||
|
расходов |
||||||||
|
ГДж/га |
% |
ГДж/га |
% |
ГДж/га |
% |
ГДж/га |
% |
|
|
Создание оросительной системы |
322 |
90 |
322 |
82 |
322 |
47 |
322 |
75 |
|
Эксплуатация (возделывание и уборка) |
31 |
10 |
66 |
18 |
352 |
53 |
102 |
25 |
|
Суммарный антропогенный вклад |
353 |
100 |
388 |
100 |
674 |
100 |
424 |
100 |
Средневзвешенная величина энергии по всей рисовой системе в дельте р. Bолги, затраченная на создание оросительной системы, в три раза превышает затраты на ее эксплуатацию, что резко снижает энергетическую эффективность производства сельскохозяйственных культур (табл. 3). Связано это с тем, что здесь для защиты от половодий и уменьшения отрицательного влияния близко залегающих минерализованных грун- товых вод необходимо строить сложные и поэтому дорогостоящие инженерные оросительные конструкции. Энергетические затраты на производство бахчевых культур в дельте р. Bолги гораздо меньшие. B условиях орошения они в среднем равны 100 ГДж/га, а в богарных – не превышают 70 ГДж/га. При этом показатель энергетической эффективности (η ) не ниже 0,9 [10].
Таблица 3. Энергетическая эффективность рисового севооборота в дельте р. Волги,( η )
|
Энергетическая эффективность |
Люцерна |
Рис |
Томаты |
Средневзвешенный показатель для агроэкосистемы |
|
Без учета затрат на строительство |
8,8 |
1,2 |
0,1 |
1,4 |
|
С учетом затрат на строительство |
0,8 |
0,2 |
0,1 |
0,3 |
Конечно, главная цель возделывания сельскохозяйственных культур – не получение биологической массы вообще, а производство продукции, которая необходима человеку. При низком ее уровне высокая биоэнергетическая производительность агроэкосистемы утрачивает смысл. Однако в последнее время очевидным становятся экологические императивы при использовании природных ресурсов. Отрицательные экологические последствия хозяйственной деятельности трудно, а подчас и невозможно оценить в энергетических или экономических показателях. Строительство оросительных систем в низовьях Bол-ги сопровождается уничтожением ценнейших водно-болотных биотопов, имеющих мировое значение, и загрязнением водоемов, которое приводит к снижению их рыбохозяйственной продук- тивности [17]. Следует также учитывать, что оросительные системы в дельте р. Bолги создавались на месте высокопродуктивных естественных хозяйственных угодий: лугов и нерестилищ. Как было показано ранее [5], энергия, запасаемая за вегетационный сезон в природных сообществах средней части дельты р. Bолги, порядка 170 ГДж/га. Существовавшие здесь экстенсивно использовавшиеся биоценозы являлись поставщиками таких продуктов питания, как мясо и рыба. Кроме того, биогенные элементы с заливаемых во время половодий угодий дельты р. Bолги поступали в акваторию Северного Каспия, обеспечивая его высокую биологическую продуктивность. Поэтому, исходя из экологических приоритетов, следует ограничивать перевод естественных угодий дельты р. Bолги в интен- сивно эксплуатируемую пашню, тем более что с позиций биоэнергетики это оказывается неэффективным мероприятием.
Список литературы Энергетическая эффективность рисового севооборота в дельте р. Волги
- Базилевич Н.И., Гребенщиков О.С., Тишков А.А. Географические закономерности структуры и функционирования экосистем. М.: Наука, 1986.
- Бульон В.Б. Первичная продукция водоемов. Л.: Наука, 1983.
- Винберг Г.Г. Первичная продуктивность водоемов. Минск: Изд-во АН БССР, 1960.
- Винберг Г.Г. Современное состояние и задачи изучения первичной продукции водоемов//Первичная продукция морей и внутренних вод. Минск, 1961.
- Голуб В.Б., Лосев Г.А., Пилипенко В.Н., Сокольская Н.И. Опыт определения первичной продукции трех растительных сообществ дельты р. Волги//Бот. журн. 1989. Т. 73, № 6.
- Голубев В.Н., Махаева Л.В., Кожевникова С.К. Опыт калориметрического изучения динамики продуктивности надземной части растительности крымской яйлы//Бот. журн. 1967. Т. 52, № 9.
- Грищенко О.М. Биологический круговорот химических элементов и энергии в пырейных лиманах Северного Прикаспия//Материалы по флоре и растительности Северного Прикаспия. Вып. 3. Ч. 2. Л.: Изд-во Геогр. о-ва СССР, 1968.
- Грищенко О.М. Динамика калорийности биомассы сообществ степных пырейных лиманов Северного Прикаспия//Материалы по флоре и растительности Северного Прикаспия. Вып. 4. Ч. 2. Л.: Изд-во Геогр. о-ва СССР, 1969.
- Зейналов Ю.А. Калориметрическое определение энергии, аккумулированной в фитомассе отдельных групп растений Азербайджана//Изв. АН АзССР. Сер. биол. наук. 1977, № 4.
- Коринец В.В., Мачулкина В.А., Иванова Е.И., Санникова Т.А. Биоэнергетическая оценка технологии производства бахчевых культур//Коняевские чтения. Сб. ст. Всерос. науч.-практ. конф. Екатеринбург: Изд-во УрГСХА, 2006.
- Малиновский К.А., Работнов Т.А. Изучение луговых биоценозов//Программа и методика биогеоценологических исследований. М.: Наука, 1974.
- Научно обоснованные системы земледелия Астраханской области. Волгоград: Нижне-Волжское кн. изд-во, 1983.
- Прищеп Л.Г., Шумаков Б.Б., Макаров И.П. (ред.). Методика биоэнергетической оценки эффективности технологий в орошаемом земледелии. М., 1990.
- Руководство по анализам кормов. Объединение «Союзсельхозхимия». М.: Колос, 1982.
- Станков Н.З. Корневая система полевых культур. М.: Колос, 1964.
- Титлянова А.А. Продуктивность травяных экосистем//Биологическая продуктивность травяных экосистем. Новосибирск: Наука, 1988.
- Холина А.П. Подходы к определению ущерба, наносимого рыбному хозяйству участниками водохозяйственной системы//Хачатуров Т.С., Теверовский Е.Н. (ред.). Экономические оценки в системе охраны природной среды СССР. Л.: Гидрометеоиздат, 1988.
- Golub V.B., Kuzmina E.V. The segetal vegetation communities of the Lower Volga valley//Укр. фитоценологiчний збiрник. Вип. 2 (7). Сер. А. Фiтосоцiология. Киiв: Фiтосоцiоцентр, 1997.
