Эпигенетический полиморфизм как показатель влияния антропогенных загрязнителей на биологические системы

Автор: Пузынина Галина Георгиевна, Левых Ална Юрьевна, Барабанщикова Наталья Сергеевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Статья в выпуске: 3-6 т.15, 2013 года.
Бесплатный доступ
Изучена изменчивость неметрических признаков в популяциях модельных видов животных – клопа солдатика и узкочерепной полёвки в градиенте урбанизации. Отмечено возрастание эпигенетической изменчивости в импактных (городских) популяциях животных по сравнению с контрольными. В исследованных популяциях клопа-солдатика выявлены фены-маркеры. Показана значимая феногенетическая дифференцированность контрольных и городских популяций узкочерепной полёвки, превышающая уровень, характерный для подразделений одной популяции, обитающих на таком же расстоянии друг от друга.
Эпигенетический полиморфизм, фенотип, флуктуирующая асимметрия, полиморфизм, адаптация, популяция
Короткий адрес: https://sciup.org/148202131
IDR: 148202131 | УДК: 504.064:
Epigenetic polymorphism as the indicator of influence of anthropogenous pollutants on biological systems
Variability of nonmetric signs in populations of model animals – red soldier bug and narrow skulled vole in urbanization gradient is studied. Increase of epigenetic variability in impact (city) populations of animals in comparison with control is noted. In studied populations of red soldier bug phene-markers are revealed. The significant phenogenetic differentiation of control and city populations of the narrow skulled vole, exceeding level, the reference for the divisions of one population living at the same distance from each other is shown.
Текст научной статьи Эпигенетический полиморфизм как показатель влияния антропогенных загрязнителей на биологические системы
Развитие фенотипа в традиционных для вида условиях происходит в пределах генетически детерминированных границ. Это подтверждается и молекулярно-генетическими исследованиями последних лет, направленными на изучение состояния генов в зависимости от факторов окружающей среды. Показано, что при не изменившейся структуре гена, его экспрессия может усиливаться или ингибироваться вплоть до полного выключения. Все это зависит от тысячи факторов как внутренних, так и внешних. В настоящее время ген рассматривается не как закрытая, а открытая система, тонко чувствующая окружающую среду [1]. Единая для популяции эпигенетическая система в пределах пороговых ограничений обеспечивает определённую фенотипическую разно-качественность особей. В связи с этим, формирующийся эпигенетический полиморфизм имеет адаптационный функциональный «экологический» смысл [2]. К явлениям адаптационного полиморфизма Н.В. Тимофеев-Ресовский относил изменения рисунка надкрылий у двухточечной божьей коровки ( Adalia bipunc-tata ) [3]. Проявление индустриального мела-низма, контролируемого генетически, в течение полутора веков наблюдали в популяции бабочек берёзовой пяденицы ( Biston betularia )
Пузынина Галина Георгиевна, кандидат биологических наук, профессор кафедры биологии и методики её преподавания.
Барабанщикова Наталья Сергеевна, студентка фенотип, флуктуирующая асимметрия, поли в Великобритании [4]. Многочисленные примеры полиморфизма окраски покровов известны для многих насекомых [5-6]. В литературе описано достаточно примеров эпигенетической изменчивости у представителей разных видов, в том числе и у млекопитающих [2, 7-9].
Цель работы: изучение эпигенетической изменчивости в популяциях животных в градиенте действия антропогенных факторов.
Объекты исследования: клоп-солдатик (Insecta, Hemiptera, Pyrrhocoris apterus L ) и узкочерепная полёвка (Mammalia, Rodentia, Mi-crotus gregalis Pall. ), распространённые и многочисленные в районе исследования, что позволило собрать репрезентативный материал.
Материалы и методы исследования. Всего обследовали 400 экземпляров клопа-солдатика из восьми локальных популяций (лесные массивы за чертой г. Ишима, парки и скверы в городе и районы города с высокой антропогенной нагрузкой) по 50 единиц в каждой; 90 черепов M. gregalis по 45 экземпляров в каждой зоне. У клопа-солдатика анализировали четыре элемента рисунка надкрылий: чёрное верхнее пятно и нижнее, чёрное верхнее окаймление на красном фоне надкрылий и нижнее. Сбор материала проводили одновременно в конце мая 2013 г. Достоверность различий между фенотипическими классами внутри каждой популяции и между ними определяли с помощью критерия χ2 К. Пирсона.
Узкочерепных полёвок (Microtus gregalis Pall.) отлавливали методом ловушко-линий, исследовали изменчивость фенов и морфотипическую структуру жевательной поверхности первого нижнего коренного зуба (M1). В работе использовали каталог фенов и определитель морфотипов жевательной поверхности зубов коренных зубов [8-9]. Фены и морфотипы подсчитывали на левой и правой сторонах черепа. Признаки, проявившие существенную (R≥0,3) достоверную связь с полом, возрастом, годом и местом сбора исключали из дальнейшего анализа [2]. Частоты фенов и морфотипов, фе-нетические дистанции (MMD) между выборками и их средние стандартные отклонения (MSD) рассчитывали на основе общепринятых формул [10-12]. Для оценки стабильности развития рассчитывали показатели флуктуирующей асимметрии (ФА) [7]. Описанные расчеты производили с помощью пакетов прикладных программ Statan и Phen [10; 12].
Результаты исследования и их обсуждение. Все выборки клопа-солдатика разбили на три группы по степени выраженности урбанизации среды. Первая группа – слабая, вторая – умеренная и третья – сильная. Представители первой группы отловлены на окраине города в лесу и Народном парке. Вторую группу составляли популяции скверов и парков в черте города. Третья группа – это выборки из районов интенсивной антропогенной нагрузки, например, района мясокомбината, автозаправок, обочин автомагистралей и городской свалки. На основании анализа полиморфизма рисунка надкрылий у клопа-солдатика суммарно по всем элементам рисунка выявлено 158 вариаций (рис. 1).
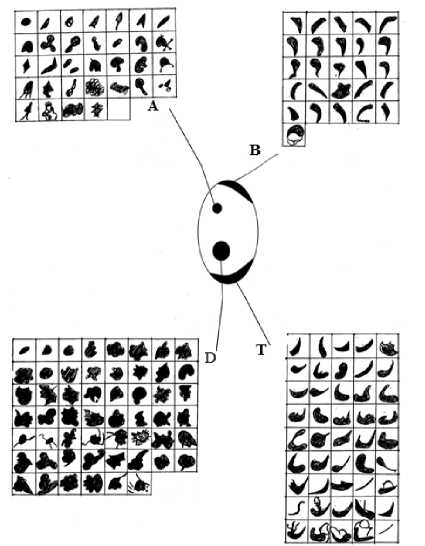
Рис. 1. Структура рисунка надкрылий клопа-солдатика
Высокий уровень вариативности характерен для элемента Д – 54 (нижнее чёрное пятно) и элемента Т – 45 (нижнее чёрное окаймление). В первой группе выявлено 29 фенов по всем элементам, во второй – 35 фенов и в третьей – 46. Частотный состав выявленных фенов в каждой из популяций различен. Из 33 фенов по чёрному верхнему пятну в первой группе встречается 6-8 , во второй группе – 1011 и в третьей – 13-14. Аналогичные результаты наблюдаются по другим элементам рисунка надкрылий. Так, по чёрному нижнему пятну из 54 фенов в первой группе встречается 6-8 фенов, во второй – 10-11 и в третьей – 11-13. По верхнему чёрному окаймлению из 26 выявленных фенов в первой группе представлено 7, во второй – 8 и в третьей – 11. По нижнему чёрному окаймлению из 45 фенов в первой группе представлено 7-8, во второй – 8 и третьей – 10. Наблюдается высокая вариативность по частотному составу фенов в каждой из популяций.
Следует отметить, что в каждой из популяций наметились господствующие фены, достоверно (Р≤0,05) отличающиеся по встречаемости от других. Так, в первой группе преобладают фены А3, А10, А14, А15, Д5, Д17, В1, В9, Т6, Т11. Во второй группе – фены А1, А3, А5, А10, Д5, Д6, Д9, В1, В3, В5, В6, В7, В10, В11, В12,Т3, Т11. В третьей группе фены – А1, Д5, Д6, В1, Т1, Т17. В первой и второй группах общими господствующими фенами являются: А3, А10, В1, Т11, Д5. Фены Д5 и В1 встречаются с высокой частотой во всех трёх группах. Сходство фенов во всех трёх группах, возможно, обусловлено существованием их в одной природно-климатической зоне.
Выявлены фены-маркеры. В первой группе фены-маркеры найдены только для нижнего чёрного окаймления (Д19, Д29) по другим элементам отсутствуют. Во второй группе фены-маркеры выявлены по всем элементам рисунка. Так, по элементу А определено 6 маркеров (А: 7,12,24,30,32, 37), по элементу Т-8 (Т:2, 4, 10, 26, 33, 34, 36, 41), по элементу Д-11 (Д: 1, 4, 13,14, 20, 33, 34, 36, 37, 39, 46), по элементу В-1 фен (В21). В третьей группе также представлены фены-маркеры по всем элементам рисунка: А28; В14; В25; Д: 7, 45, 48, 49, 50, 53, 54; Т: 38, 40, 41, 45. Возрастание антропогенной нагрузки приводит к увеличению количества вариаций и формированию особых для каждой зоны фенов-маркеров, которые могут служить индикаторами каких-либо антропогенных загрязнителей.
Анализировали сходство и различие вариаций по всем элементам левого и правого надкрылий. Результаты представлены в табл. 1. Как видно из таблицы процент асимметрии возрастает в зависимости от степени урбани-зированности популяции по элементам А, Д и Т. Исключение составляет элемент В (верхнее чёрное окаймление), по которому асимметрия от первой ко второй группе возрастает, а в третьей группе резко снижается. Отмеченная стабильность элемента В согласуется с литературными данными [5-6].
Таблица 1. Асимметричность проявления фенов в элементах A-T рисунка надкрылий (%)
|
Элементы |
Группы по степени урбанизированности |
||
|
I |
II |
III |
|
|
А |
35,0 |
66,6 |
100 |
|
В |
40,0 |
56,7 |
36,8 |
|
D |
35,7 |
48,7 |
52,2 |
|
T |
26,6 |
52,9 |
65,0 |
В исследуемых популяционных группах узкочерепной полёвки выявлено 60 фенов жевательной поверхности нижних коренных зубов. Каждая популяционная группа имеет особую фенетическую структуру, которая определяется качественным составом и количественным соотношением разных фенов. Городская выборка характеризуется наиболее разнообразным составом фенов (60 признаков). В выборке из естественного местообитания отмечен 51 фен. Однообразие местообитаний на территории г. Ишима позволяет предположить, что разнообразие фенетической структуры ишимской популяционной группы поддерживается за счёт дестабилизирующего влияния на процессы развития животных изменённой городской среды, насыщенной загрязнителями различного происхождения.
Количественные различия проявляются в различном частотном распределении отдельных фенов в каждой выборке. Так, в контрольной выборке 100%-ной встречаемостью характеризуются фены Cins1 и Sics1; на 2-м месте по встречаемости находится фен Ppt2 (92%); на 3-м месте – фен Ali1 (76%); высокой частотой отличатся фены Lipt1 (72%) и Ili2 (50%); наиболее редко (1%) встречаются фены Pat3, Ppt3, Ain4, Phc4, Cens3, Lipt3, Ili4, Ili5. B ишимской выборке самым распространённым является фен Cins1 (93%), на втором месте – фен Ali1 (79%), на 3-м – Lipt1(75%); чрезвычайно редки фены Fli9 (0,04%), Ppt4 (0,04%), очень редки – Ali7 (0,4%), Ili7 (0,4%), редки фены Ali6 (1%), Fli7 (1%), Fli8 (1%), Ili (1%). Множественное сравнение по отдельным признакам выявило статистически значимые межгрупповые различия по 26 фенам (табл. 2).
Таблица 2. Множественное фенетическое сравнение разных популяционных групп узкочерепной полевки
|
№ п/п |
Фены |
Фенетическая дистанция |
χ 2 |
|
1 |
Ali 2 |
-0,319 |
20,37*** |
|
2 |
Ali 4 |
-0,095 |
6,82*** |
|
3 |
Fli 1 |
0,432 |
28,3*** |
|
4 |
Fli 2 |
0,048 |
4,00* |
|
5 |
Fli 3 |
0,132 |
9,32** |
|
6 |
Cr 1 |
0,158 |
11,0*** |
|
7 |
Cr 2 |
0,471 |
30,72*** |
|
8 |
Cin 2 |
0,060 |
4,77* |
|
9 |
Pat 3 |
0,701 |
45,1*** |
|
10 |
Cem 3 |
0,093 |
6,81** |
|
11 |
Ppt 2 |
0,928 |
58,6*** |
|
12 |
Ppt 3 |
1,440 |
90,1*** |
|
13 |
Ain 1 |
0,083 |
6,12* |
|
14 |
Ain 4 |
0,305 |
19,9*** |
|
15 |
Phc 4 |
0,284 |
18,9*** |
|
16 |
Pats 1 |
0,079 |
5,96* |
|
17 |
Pats 2 |
0,234 |
15,7*** |
|
18 |
Pats 3 |
0,585 |
37,8*** |
|
19 |
Pats 4 |
0,072 |
5,54* |
|
20 |
Lips 3 |
0,387 |
25,5*** |
|
21 |
Ppts 2 |
0,859 |
54,8*** |
|
22 |
Cins 1 |
5,395 |
335,3*** |
|
23 |
Cins 2 |
1,979 |
124,3*** |
|
24 |
Sics 1 |
1,647 |
104,2*** |
|
25 |
Sics 2 |
2,082 |
130,7*** |
|
26 |
Cint 1 |
0,069 |
5,26* |
|
MMD |
0,308 |
||
|
MSD |
0,0029 |
||
|
χ 2 |
1230,4*** |
||
|
d.f. |
MMD>2 MSD |
||
Примечание: в таблице представлены фены, по которым выявлены достоверные межгрупповые различия
Уровень фенетической дистанции между выборками выше, нежели определяет степень пространственно-географической изолированности [2]. Это позволяет предположить, что полученные результаты обусловлены влиянием антропогенных загрязнителей. В изучаемых выборках узкочерепной полёвки выявлены 5 морфотипов первого нижнего коренного зуба, соответствующих литературным данным [9].
Выборки узкочерепной полёвки из естественного местообитания и городских лесопарков значимо различаются только по частоте встречаемости II морфотипа (в естественном местообитании - 0,17±0,04; в городской среде - 0,33±0,05; t=2,35; при P≤0,05). В городской выборке M. gregalis отмечается возрастание доли флуктуирующей асимметрии по указанному морфотипу (c 0,02 % до 96%). Очевидно, можно сделать вывод о дестабилизирующем влиянии городской среды на эпигенетические процессы в популяциях.
Выводы: исследование эпигенетической изменчивости в популяциях клопа-солдатика и узкочерепной полёвки показало, что в градиенте урбанизации возрастает полиморфизм по анализируемым признакам. Так, у клопа-солдатика в третьей группе, испытывающей наибольшую антропогенную нагрузку, происходит изменение соотношения фенов, увеличивается их количество и возрастает их разнообразие, появляются фены-маркеры, которые могут служить индикаторами антропогенных загрязнителей. Практически по всем элементам рисунка отмечается рост асимметрии в зависимости от уровня антропогенной нагрузки. В городской популяции узкочерепной полёвки увеличивается разнообразие фенов неметрических признаков, изменяется их встречаемость, возрастает флуктуирующая асимметрия по ряду признаков и фенетическая дистанции между выборками.
Список литературы Эпигенетический полиморфизм как показатель влияния антропогенных загрязнителей на биологические системы
- Сеньков, О. Гены, которые мы меняем//В мире науки. 2008. №11. С.50-59.
- Васильев, А.Г. Эпигенетические основы фенетики: на пути к популяционной мерономии. -Екатеринбург: Академкнига, 2005. -640 с.
- Лусис, Я.Я. О биологическом значении полиморфизма окраски у двуточечной божьей коровки Adalia bipunctata//Latv. Entomologs. 1961. №4. C. 3-29.
- Кайданов, Л.З. Генетика популяций. -М.: Высшая школа, 1996. 320 с.
- Батлуцкая, И.В. Изменчивость меланизированного рисунка насекомых в условиях антропогенного воздействия: Монография. -Белгород: БелГУ, 2003. 168 с.
- Батлуцкая, И.В. Применение МАИ для биоиндикаторной оценки изменчивости меланизированного рисунка покровов насекомых из различных наземных экосистем//Науч. вед. Серия экология. 2005. Вып. 3, №1 (21). С. 30-37.
- Захаров, В.М. Асимметрия животных. -М.: Наука, 1987. 216 с.
- Ларина, Н.И. Каталог основных вариаций краниологических признаков у грызунов/Н.И. Ларина, И.В. Еремина//Фенетика природных популяций. -М.: Наука, 1988. С. 8-53.
- Бородин, А.В. Определитель зубов полёвок Урала и Западной Сибири (поздний плейстоцен-современность). -Екатеринбург: УрО РАН, 2009. 99 с.
- Гашев, С.Н. Статистический анализ для биологов (Руководство по использованию пакета программ «Statan-1996»). -Тюмень: ТГУ, 1998. 22 с.
- Васильев, А.Г. Пакет прикладных программ «Фен» Phen 3,0. Путеводитель для пользователей. -Екатеринбург, 1995. 113 с.
- Hartman, S.E. Geographic variation analysis of Dipodomys ordii using nonmetric cranial traits//J. Mammal. 1980. V.61. №3. P. 436-448.
