Эволюция экосистем нижнепермских скелетных холмов Северного Урала

Бесплатный доступ
Рассмотрены экосистемы нижнепермских скелетных холмов Северного Урала. Выделено три внутренних и девять внешних факторов, влиявших на их эволюцию. Показано, что нижнепермские скелетные холмы были ограничены базисом действия волн. Экологическая сукцессия холмов определена как алло-аутогенная. Повторение стадий экологической сукцессии позволяет отнести нижнепермские разрезы рифогенного типа к ритмитам скелетных холмов.
Экосистемы, скелетные холмы, ассельский и сакмарский ярусы, северный урал
Короткий адрес: https://sciup.org/149129007
IDR: 149129007
Evolution of the ecosystems of lower Permian skeletal mounds of Northern Urals
The ecosystems of the Lower Permian skeletal mounds of the Northern Urals were examined. Three inner and nine outer factors, affected on their evolution, were extract. Indicate that the Lower Permian skeletal mounds were limited by a wave base. The ecologic succession of the mounds were defined as an allo-autogenous type. The recycling of the ecologic succession stage allow to take the Lower Permian reefogenic types sections to skeletal mounds rhytmits.
Текст научной статьи Эволюция экосистем нижнепермских скелетных холмов Северного Урала
Рифы и более простые постройки в современных морях и в геологическом прошлом представляют собой уникальные органогенные сооружения и являются своеобразными экосистемами, чуткими и точными показателями тектонического режима и палеогеографических условий, специфическими фациями карбонатонакопления, своеобразными формами рельефа морского дна [19]. Для определения генетического типа органогенного сооружения необходимо прежде всего рассмотреть его палеоценотические структуры. Именно палеоценотический и палеоэкологический анализ взаимоотношений между организмами и окружающей средой, запечатленных в породе, позволяет делать некоторые важные обобщения [1].
Используемая в статье классификация органогенных сооружений основана на экологической сукцессии рифовых экосистем [30]. Сукцессия рифовых систем — это закономерная смена биоценозов во времени, не связанная с эволюционными изменениями видов [9]. Развитие экосистемы состоит в изменении во времени видовой структуры и биоценотических процессов. В отсутствие внешних нарушающих процессов сукцессия рифовых экосистем представ- ляет собой направленный и, следовательно, предсказуемый процесс [8, с. 165]. Различаются аутогенная сукцессия, происходящая при развитии системы без вмешательства извне, и аллогенная, развивающаяся в результате внешнего воздействия [19]. Внешние (аллогенные) стрессы могут прерывать сукцессию еще до достижения экосистемой климакса. Если такие стрессы случаются регулярно, сукцессия становится циклической.
Нижнепермские органогенные постройки, широко распространенные на Северном Урале, вскрываются в разрезах рек Илыч, Печора и Унья (рис. 1). Стратиграфическая приуроченность этих органогенных сооружений дана в работах Н. В. Калашникова, З. П. Михайловой [5, 6] и Е. С. Пономаренко с соавторами [14]. Первые данные об их строении можно получить из работ В. А. Варсанофьевой [4] и А. И. Равикович [15]. Более детальные исследования были предприняты в начале XXI века. В результате было выявлено строение отдельных органогенных построек [11, 17, 18], выделены стадии экологической сукцессии [10, 11], а тип построек определен как скелетные холмы [1, 10, 11, 17]. После всестороннего анализа накопленного материала была создана схемати-
Puc. 1. Расположение нижнепермских разрезов

ческая модель строения нижнепермских скелетных холмов [3, 12], определена их общая трофическая структура [3], состоящая из шести ступеней (рис. 2). Несмотря на наличие большого количества работ, в которых рассматриваются нижнепермские скелетные холмы, вопрос о влиянии окружающих сред на экосистемы до сих пор освещен недостаточно. Прежде всего необходимо выяснить взаимосвязи между организмом и средой, которые позволят не только наиболее точно определить генезис органогенной постройки, но и проследить историю ее развития от зарождения до исчезновения.
Ниже приводятся результаты исследования влияния различных факторов (биотических и абиотических) на эволюцию экосистем нижнепермских скелетных холмов, а также реакции организмов на изменение среды.
Развитие нижнепермских скелетных холмов
Изучение нижнепермских органогенных построек показало, что в них присутствуют только пионерные стадии: стабилизация и колонизация [30], что свидетельствует о неполноте экологической сукцессии. Специфические образования нижнепермских скелетных холмов были выделены в финальную стадию [29] и позднее названы стадией деструкции [10, 11]. Согласно П. Куперу [23] у каждой экосистемы может быть своя трофическая структура, которая определяется экологией и способом питания организмов. На палеобиоценозы оказывали влияние не только абиотические факторы, но и биотические — представители других биоценозов. Выяснение взаимосвязей как между отдельными организмами, так и между их ассоциациями, а также выяснение влияния абиогенных факторов, позволяет наиболее полно охарактеризовать возникновение и развитие органогенных структур.
В разрезе Мича Ласта (обн. 82, 83 на р. Илыч) развиты две органогенные постройки типа скелетных холмов: ассель-ская и нижнесакмарская, а разрез в целом можно определить или классифицировать как ритмит скелетных холмов (рис. 3).
Первый puтм (acceльcкuй). Стадия стабилизации представлена био-кластовыми криноидными и тубифите-сово-криноидными известняками с участками палеоаплизиновых, тубифитесо-во-мшанковых и филлоидно-водоросле-вых биоцементолитов мощностью
|
2 с и S с |
IV уровень |
Хищники |
Палеоаплизины, аммоноидеи |
||
|
III уровень |
Всеядные |
Гастроподы, остракоды, трилобиты |
|||
|
II уровень |
Наддонные фильтраторы |
Ветвистые мшанки, криноидеи |
|||
|
I уровень |
Придонные фильтраторы |
Брахиоподы, сетчатые мшанки, мелкие фораминиферы и фузулиноиды |
|||
|
3 8 1 Он с |
II уровень |
Фототрофы |
Зеленые водоросли |
||
|
I уровень |
Хемо- и фототрофы |
Микробы, цианобактерии |
|||
Puc. 2. Трофическая структура рифовых экосистем нижнепермских холмов Северного Урала
|
л О m |
3 CL, |
с с |
§ > i ) |
3 й о 1 |
св |
Гидродинамика |
Контролирующие факторы |
||||
|
< |
| 3 •в< |
3 S S |
5 ! |
3 а о |
внутренние (аутогенные) |
внешние (аллогенные) |
|||||
|
1 Он о Он |
3S 1 CQ |
В |
<П |
\ / \ / \ / |
Коллапс экосистемы |
Достижение базиса действия волн Постепенная активизация гидродинамики Быстрая трансгрессия и достижение спокойноводных условий Осушение (резкая регрессия) Осушение (резкая регрессия) |
|||||
|
1 |
1 |
о Н |
1 1 1 1 1 1 |
1 1 IMI 1 1 1 Mil |
Смена вертикальных форм роста пластинчатыми Оккупация твердых субстратов, возникновение неблагоприятных условий Стабилизация субстрата кальцимикробными сообществами |
||||||
|
d |
1 5 |
tri |
|||||||||
|
и а о 81 РН |
’§ Сц 6 |
В ев Я |
S |
Остановка роста постройки |
|||||||
|
0 0__о __0__0__ 0 0.___0 __0__0__ 0 о__0 __О__0__ |
Оккупация твердых субстратов пластинчатыми организмами |
||||||||||
|
§ СТ I |
^ н |
Укрепление субстрата пластинчатыми организмами |
|||||||||
|м М| 1 |——I 2 |ИР 4 |%%| 5 ЕЗб
Puc. 3. Экологическая сукцессия и контролирующие факторы в нижнепермской органогенной постройке на р. Илыч (обн. 82, 83).
1—3 — формы роста: 1 — вертикальная, 2 — пластинчатая, 3 — ветвистая; 4 — кальцимик-робные сообщества; 5 — биокластовый материал; 6 — биокластовый материал в биологически индуцированном цементе

-
2.5 м. Зарождение скелетного холма происходило, по-видимому, следующим образом: накапливались криноидные биокластовые пески (вероятно, в обстановках течений), на которых поселялись палеоаплизиновые, тубифитесово-мшанковые и реже водорослевые сообщества, закреплявшие осадок. Трофическая структура начальной стадии органогенной постройки включает продуцентов I и II уровней, а также консументов I, II и IV уровней. Роль продуцентов здесь состояла не только в поставке питания для гетеротрофов, но и прежде всего в укреплении их скелетов в обстановках слабых течений, вымывавших карбонатный ил. Только с помощью раннедиагенетической крустификации по микробиальным пленкам создавался каркас нижнепермских скелетных холмов [2, 3].
Стадия колонизации представлена биогермными мшанковыми и тубифи-тесово-мшанковыми биоцементолита-ми с участками биокластовых известняков мощностью 24.1 м. На отдельных уровнях наблюдаются небольшие скопления мелких (до 5 мм) неокатанных обломков мшанковых биоцементоли-тов. Основными колонизаторами являются сетчатые мшанки, в незначительных количествах отмечаются брахиопо-ды, мелкие прикрепляющиеся форами-ниферы. По микробиальным пленкам на органических остатках развит раннедиагенетический крустификационный кальцитовый цемент [11]. Трофическая структура скелетного холма на стадии колонизации двухступенчатая, представлена продуцентами I уровня (фото- и хе-мотрофами: тубифитесами, кальцимик-робами) и консументами I уровня (придонными фильтраторами: мшанками, брахиоподами, фораминиферами) с преобладанием гетеротрофных организмов. Стадия колонизации прерывалась на границе ранне- и среднеассель-ского времени, что было связано с резким обмелением бассейна и возможным выводом пород в зону эрозии [11, 14]. Новая трансгрессия моря привела к восстановлению экосистемы без повторения стадии стабилизации, так как субстрат уже был сформирован [11]. Таким образом, историю формирования постройки на стадии колонизации можно охарактеризовать следующим образом: твердый субстрат, образовавшийся на стадии стабилизации, был оккупирован сначала тубифитесово-мшанковыми сообществами, а затем преимущественно сообществами сетчатых мшанок. Раз- витие скелетного холма регулировалось базисом действия волн [11].
Рост органогенной постройки на стадии колонизации завершился в связи с обмелением и крупным перерывом в осадконакоплении, охватывающим позд-неассельское (шиханское) время [14].
Втоpой puтм (paннecaкмapcкuй). Стадия стабилизации представлена строматолитоподобными и комковато-сгустковыми известняками суммарной мощностью 15.1 м и имеет раннесак-марский [14] возраст. Нижняя часть постройки (4.1 м) сложена строматолитоподобными известняками с остатками многочисленных ветвистых мшанок и редких брахиопод. На мшанки наросли колоний тубифитесов и цианобактерий [11]. В незначительных количествах отмечаются обломки члеников кринои-дей. В нижней части строматолитоподобных известняков наблюдаются фрагменты органогенного материала, трещины усыхания и ходы илоедов [11]. Верхняя часть отложений стадии стабилизации представлена комковато-сгустковы-ми известняками с многочисленными сетчатыми мшанками и редкими бра-хиоподами. Трофическая структура трехступенчатая и характеризуется преобладанием продуцентов I уровня (каль-цимикробов, тубифитесов, цианобактерий), поставлявших основное количество питательных веществ консументам. Гетеротрофные организмы представляют 3-ю и 4-ю ступени пищевой трофической пирамиды [3] и охарактеризованы придонными (консументами I уровня, брахиоподами) и наддонными (консументами II уровня, ветвистыми мшанками, криноидеями) фильтраторами. Таким образом, рост раннесакмарско-го скелетного холма стимулировался широким развитием кальцимикробных организмов, готовивших субстрат и поставлявших питание гетеротрофам. Быстрое углубление осадочного бассейна обусловило сохранение колоний мшанок в положении роста. Нараставшие на них тубифитесы и цианобактерии, вследствие быстрой кальцификации колоний, только укрепляли скелеты беспозвоночных. В конце стадии стабилизации, вероятно, произошло некоторое обмеление бассейна, приведшее к смене морфологии колоний мшанок.
Стадия колонизации представлена пятнистым чередованием биогермных палеоаплизиновых, водорослево-туби-фитесовых и водорослевых биоцемен-толитов мощностью 2.0 м. Пятнистость обусловлена мозаичным расположени- ем скоплений остатков породообразующих организмов. Вверх по разрезу начинают преобладать остатки палеоапли-зин [11]. Трофическая структура состоит из трех ступеней: продуцентов I (ту-бифитесов, кальцимикробов) и II (зеленых филлоидных водорослей) уровней, а также консументов IV уровня (палео-аплизин). Стадия колонизации началась с оккупации твердых субстратов зелеными филлоидными водорослями. Эти водоросли имели вертикальную форму роста (erect growth [21, 22]) и могли активно развиваться в зоне между нижним пределом фотической зоны и волновой активностью [24, 25, 32]. Низкое биоразнообразие в филлоидно-водорослевых зарослях было замечено многими исследователями и объяснялось тем, что они могли продуцировать ядовитые вещества, которые ограничивали проникновение других организмов в области их развития [25—27, 32—34]. Химическая защита, используемая водорослями, известна и в современных рифах [28, 31]. Смену водорослевых сообществ палео-аплизиновыми можно объяснить постепенным падением уровня моря и изменением окружающей среды: незначительные волнения выводили ядовитые вещества, а усиление гидродинамики сделало невозможным благополучное существование тонких пластинчатых организмов с вертикальным ростом. Однако пластинчатые формы сессильных организмов, в частности палеоаплизин, были лучше приспособлены к более подвижным водам. Преобладание в конце стадии колонизации толстопластинчатых палеоаплизин свидетельствует не только об активизации гидродинамики, но и о значительно большем поступлении питательных веществ. Развитие органогенной постройки на стадии колонизации происходило в различных условиях — от спокойно-водных до слабоактивных, что нашло отражение в различии палеоэкотипов палеоаплизин и в целом в экосистемах скелетного холма.
Стадия деструкции. На прекращение роста органогенной постройки Мича Ласта прежде всего указывает накопление иловых известняков с линзами биокластовых палеоаплизиновых биоцементолитов, представляющих фации желобов [1]. Вышележащие поли-биокластовые биоцементолиты завершают разрез постройки [11]. Мощность отложений, характеризующих стадию деструкции, достигает 7.5 м. На этой стадии органогенная постройка была выведена в зону активной гидродинамики, что привело к разрушению хрупких скелетов основных организмов каркасо-строителей (палеоаплизин, филлоидных водорослей, мшанок и др.) еще до формирования биологически индуцированного крустификационного цемента вокруг биокластов [10, 11]. Коллапс экологической сукцессии завершил формирование органогенного сооружения.
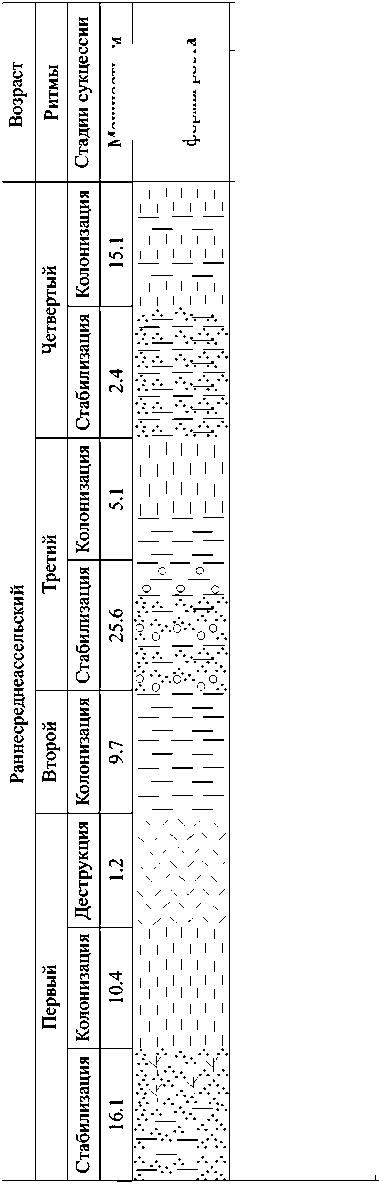
Разрез обн. 61 на р. Печора характеризует западные краевые зоны органогенной постройки раннесредне-ассельского возраста. Само органогенное сооружение мощностью 200— 250 м вскрыто восточнее — в бассейне р. Б. Шайтановка (обн. 63а по: [6]). Здесь также можно выделить ритмит скелетных холмов, состоящий из четырех ритмов (рис. 4).
Пepвый puтм. Стадия стабилизации представлена известняками мощностью 16.1 м — комковато-сгустковыми в нижней части разреза и цианобактериальными в верхней. В известняках обнаружены редкие остатки тубифите-сов, сетчатых мшанок, брахиопод и ост-ракод. Трофическая структура трехступенчатая: преобладают продуценты I уровня (кальцимикробы, цианобактерии, тубифитесы), консументы имеют незначительное распространение и представлены придонными фильтрато-рами (мшанками, брахиоподами, фора-миниферами) и всеядными (остракода-ми, трилобитами). Отложения стадии стабилизации формировались ниже базиса действия волн в условиях со спокойной гидродинамикой.
Стадия колонизации представлена цианобактериальными биоцементоли-тами мощностью 10.4 м с редкими обломками зеленых дазикладиевых и красных водорослей, мшанок, брахиопод, криноидей. Трофическая структура одноступенчатая и включает только продуцентов I уровня (цианобактерий, каль-цимикробов). Возможно, высокая гидродинамика среды и значительный при-внос биокластового материала не благоприятствовали развитию типичных нижнепермских экосистем, поэтому на стадии колонизации оккупация твердых субстратов здесь осуществлялась только цианобактериями.
Стадия деструкции представлена полибиокластовыми биоцементолита-ми мощностью 1.2 м с обломками члеников криноидей, колоний тубифитесов, зеленых дазикладиевых и красных водорослей, мелких фораминифер, мшанок и брахиопод. Танатоценоз характеризу-
|
4 D D г |
О д О о |
|
Гидродинамика |
Контролирующие факторы |
|||
|
5 о 3 и |
ю л 5 |
8 >s о Е U |
внутренние (аутогенные) |
внешние (аллогенные) |
|
Чередование пластинчатых и вертикальных форм роста |
Незначительные колебания гидродинамики |
|||
|
Укрепление субстрата кальцимикробами и сетчатыми мшанками |
Увеличение поступления биокластового материала |
|||
|
Смена пластинчатых форм роста вертикальными |
Постепенное понижение гидродинамики |
|||
|
Повышение |
||||
|
Повторение стадии стабилизации |
гидродинамики Повышение привноса биокластового материала Повышение привноса биокластового материала |
|||
|
Повторение стадии стабилизации |
||||
|
Оккупация твердых субстратов сетчатыми мшанками |
Постепенное понижение гидродинамики |
|||
|
Резкое обмеление и повышение привноса |
||||
|
биокластового материала |
||||
|
Оккупация твердых субстратов цианобактериями |
Привнос биокластового материала |
|||
|
Укрепление субстрата пластинчатыми мшанками и кальцимикробами |
||||
Puc. 4. Экологическая сукцессия и контролирующие факторы в нижнепермской органогенной постройке на р. Печора (обн. 61). Условные обозначения см. на рис. 3
ется наличием продуцентов I (кальци-микробов, тубифитесов) и II (красных и зеленых водорослей) уровней, а также консументов I (брахиопод, форамини-фер) и II (криноидей) уровней. Усиленный привнос биокластового материала, а также разрушение тонкоскелетных организмов, слагавших экосистему холма, вызвали её коллапс.
Втоpой puтм. Стадия колонизации представлена биогермными мшан-ковыми биоцементолитами мощностью
-
9.7 м. В незначительном количестве в них содержатся микробиальные комочки и сгустки, остатки тубифитесов, зеленых филлоидных водорослей, гастропод. В верхней части разреза отмечаются участки скоплений литокластов, свидетельствующие об интенсификации гидродинамики. Трофическая структура четырехступенчатая и представлена продуцентами I (кальцимикробами, ту-бифитесами) и II (зелеными филлоид-ными водорослями) уровней, а также
консументами I (мшанками) и III (гастроподами) уровней. При понижении гидродинамической активности происходила оккупация твердых субстратов сетчатыми мшанками, а резкое ее усиление могло привести к остановке развития скелетного холма и, возможно, к перерыву в осадконакоплении.
Tpeтuй puтм. Стадия стабилизации представлена чередованием разных известняков: комковато-сгустковых, цианобактериальных и биокластовых со спаритовым цементом. В первых двух типах пород часто встречаются сетчатые мшанки, мелкие фораминиферы, участками преобладают тубифитесы. Редко обнаруживаются остатки зеленых филлоидных водорослей и остракод, фрагменты члеников криноидей, брахиопод. В пятиступенчатой трофической структуре выявлены продуценты I (кальцимикробы, цианобактерии, тубифитесы) и II (зеленые филлоидные водоросли) уровней, а также консументы I (мшанки, фораминиферы, брахио-поды), II (криноидеи) и III (остракоды) уровней. Цианобактериальные сообщества обитали в мелководных гидродинамически активных условиях, что не способствовало развитию типичных раннепермских экосистем. На последних этапах стадии стабилизации накапливались преимущественно криноидные пески с поселениями мшанок. В незначительных количествах были развиты толстопластинчатые палеоаплизины, фузули-ноиды, брахиоподы. Трофическая структура этой части стадии стабилизации была четырехступенчатой. Продуценты I уровня были представлены каль-цимикробами, по которым развивались крустификационные оторочки, а гетеротрофные организмы были весьма многочисленными и разнообразными и представлены консументами I (мшанками, фузулиноидами, брахиоподами), II (криноидеями) и IV (палеоаплизина-ми) уровней. Начало стадии стабилизации было обусловлено локальным прогибанием морского дна и последовавшим за ним понижением гидродинамической активности. Это в свою очередь привело к усиленному привносу био-кластового материала с более возвышенных участков морского дна, что не благоприятствовало развитию экосистем. Последующее падение уровня моря привело к появлению в основном мета-зойных стабилизаторов.
Стадия колонизации (5.1 м) выделяется постепенной сменой биогермных палеоаплизиновых биоцементолитов сначала филлоидно-водорослевыми, а затем цианобактериальными. Они разделены прослоями биокластовых известняков со спаритовым цементом, мощность которых увеличивается вверх по разрезу. Шестиступенчатая трофическая структура состоит из продуцентов I (кальцимикробов, цианобактерий, ту-бифитесов) и II (зеленых филлоидных и дазикладиевых водорослей) уровней, а также их консументов I (мшанок, фора-минифер, брахиопод), II (криноидей), III (остракод) и IV (палеоаплизин) уровней. Смена сообществ экосистемы была вызвана постепенным понижением рельефа морского дна. На начальном этапе стадии колонизации преобладали па-леоаплизины, свидетельствующие о подвижной гидродинамике. Выше по разрезу их вытесняют зеленые филлоидные водоросли. Смена формы роста с горизонтальной (палеоаплизины) на вертикальную (филлоидные водоросли) была связана с понижением гидродинамики. Увеличение привноса биокластового материала обусловило смену экосистем в сторону более стрессоустойчивой.
Длительное локальное прогибание морского дна привело к остановке роста органогенной постройки в рассматриваемой области, на это указывает наличие полибиокластовых несортированных известняков с пелитоморфным цементом (мощность 32.4 м). Среди органических остатков в них значительное место занимают обломки колоний цианобактерий, тубифитесов, палеоапли-зин, мшанок и других организмов, что позволяет отнести эти известняки к коллювиальным образованиям пологих склонов построек [12].
Чeтвepтый puтм. Стадия стабилизации (2.4 м) представлена комкова-то-сгустковыми известняками с незначительным количеством остатков фил-лоидных водорослей, сетчатых мшанок, раковин фораминифер, брахиопод, гастропод, обломками члеников криноидей и колоний красных водорослей. Двухступенчатая трофическая структура характеризуется преобладанием продуцентов. На этой стадии происходила стабилизация мягких грунтов кальцимик-робами (продуцентами I уровня), сетчатыми мшанками (консументами I уровня) в гидродинамически спокойных условиях при уменьшении привноса биокластового материала.
На стадии колонизации (15.1 м) отчетливо проявляется чередование био-гермных филлоидно-водорослевых и палеоаплизиновых биоцементолитов.
Смена экосистем, по всей видимости, происходила из-за незначительных флуктуаций уровня моря со сменой гидродинамики.
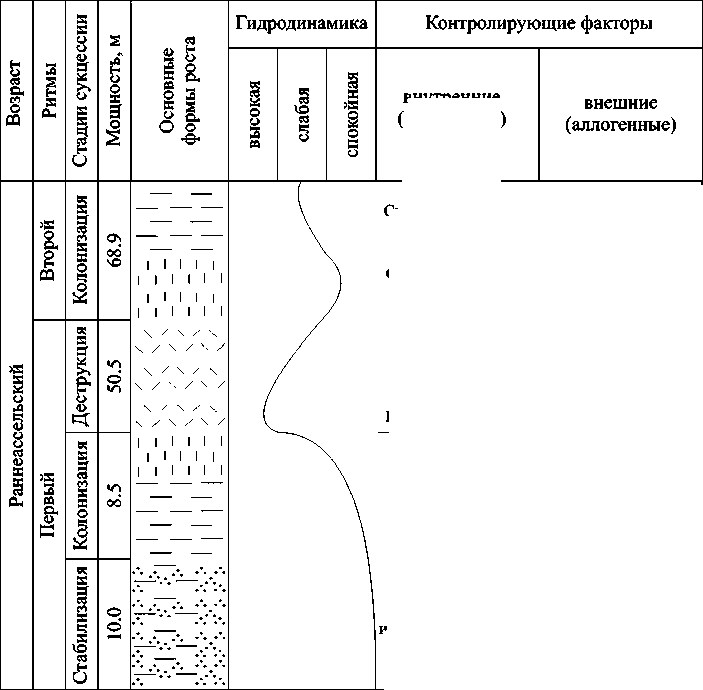
В разрезе Писаный Камень (обн. 28, р. Унья) вскрывается ритмит скелетных холмов, состоящий из двух ритмов (рис. 5). Здесь имеется уникальная возможность увидеть в нижней части разреза образования центральной части скелетных холмов, а в верхней — восточных краевых зон. Это связано с начавшейся в конце раннеассельского времени резкой трансгрессией моря, которая привела к миграции органогенных построек в западном направлении.
Пepвый puтм. Стадия стабилизации (10.0 м) в нижней части разреза представлена биокластовыми известняками с пелитоморфным цементом, в верхней — комковато-сгустковыми известняками с участками биогермных мшанковых биоцементолитов. В известняках местами отмечаются скопления раковин брахиопод, редкие фрагменты колоний цианобактерий, тубифитесов, раковин фораминифер, члеников кри-ноидей, щитков трилобитов. Трофическая структура четырехступенчатая. Нижняя ступень представлена продуцентами I уровня (кальцимикробами, цианобактериями, тубифитесами), а также консументами I (фораминифера-ми, брахиоподами), II (ветвистыми мшанками, криноидеями) и III (трилобитами) уровней. Формирование первого скелетного холма начиналось с укрепления мшанками и кальцимикро-бами мягкого илово-песчаного дна в спокойно-водных условиях.
Стадия колонизации (8.5 м) выделяется преобладанием биоцементоли-тов — в основании биогермных мшанковых, а выше по разрезу — биогерм-ных филлоидно-водорослевых. В незначительном количестве отмечаются колонии цианобактерий, тубифитесов, раковины фораминифер, брахиопод, обломки члеников криноидей. На начальных этапах стадии колонизации трофическая структура органогенной постройки была трехступенчатой, в ней существовали продуценты I уровня (кальцимикробы, тубифитесы), но большую роль играли консументы I (брахиоподы, фораминиферы, сетчатые мшанки) и II (криноидеи) уровней. Позднее произошла смена экосистемы, что в трофической структуре проявилось в преобладании продуцентов II уровня — зеленых филлоидных водорослей, обладавших более высокой кон-
Коллапс экосистемы
Укрепление мягкого илового дна мшанками и кальцимикробами
Повышение гидродинамики внутренние (аутогенные)
Достижение базиса действия волн
Снижение гидродинамики
Оккупация твердого дна филлоидными водорослями
Вытеснение мшанок филлоидными водорослями Оккупация твердого дна сетчатыми мшанками
Смена вертикальных форм роста пластинчатыми
Puc. 5. Экологическая сукцессия и контролирующие факторы в нижнепермской органогенной постройке на р. Унья (обн. 28). Условные обозначения см. на рис. 3
курентоспособностью, обусловленной выделением ядовитых веществ.
Стадия деструкции (50.5 м) сложена биоцементолитами, полибиокласто-выми в нижней части разреза и биокла-стовыми палеоаплизиновыми — в верхней. Иногда в разрезе присутствуют линзовидные слои (0.6—1.5 м) биокластовых с пелитоморфным цементом или комковато-сгустковых известняков. В танатоценозе выявлены продуценты I (каль-цимикробы, тубифитесы, цианобактерии) и II (зеленые филлоидные и дазик-ладиевые водоросли, красные водоросли) уровней, а также консументы I (фо-раминиферы, мшанки, брахиоподы,), III (гастроподы) и IV (палеоаплизины, аммоноидеи) уровней. Смену водорослевых сообществ палеоаплизиновыми можно объяснить постепенным падением уровня моря, в результате чего даже незначительные волнения снижали концентрацию ядовитых веществ, а усиление гидродинамики сделало невозможным благополучное существование водорослей с вертикальным ростом. Достижение уровня вод с высокой гидродинамикой, возможно, было связано с большой скоростью роста филлоидных водорослей [22], а с не флуктуациями уровня моря. Таким образом, коллапс данной экосистемы на стадии деструк- ции был обусловлен высокой гидродинамикой среды.
Втоpой puтм. Стадия колонизации (68.9 м) представлена биогермны-ми филлоидно-водорослевыми и в верхней части разреза палеоаплизиновыми биоцементолитами. Трофическая структура состоит из трех ступеней: продуцентов I уровня (тубифитесов, каль-цимикробов) и II уровня (зеленых фил-лоидных водорослей), а также консументов IV уровня (палеоаплизин). Начало стадии колонизации характеризовалось оккупацией твердых субстратов зелеными филлоидными водорослями. Смена водорослевых сообществ палеоаплизи-новыми объясняется постепенным падением уровня моря и изменением окружающей среды. Развитие органогенной постройки на стадии колонизации происходило в различных условиях: от спокойно-водных до слабоактивных, что нашло отражение в экосистемах.
Начавшаяся в конце раннеассель-ского времени трансгрессия моря проявилась в миграции органогенной постройки на запад, в сторону внутреннего моря. Это фиксируется развитием филлоидно-водорослевых и палеоапли-зиновых биоцементолитов среднеас-сельского возраста в обн. 25 «Чертов стульчик» [5]. В разрезе Писаный Ка- мень средневерхнеассельские отложения, по всей видимости, представлены восточной краевой зоной органогенной постройки и характеризуются неравномерным чередованием комковато-сгустковых известняков, биогермных и био-кластовых биоцементолитов и биоклас-товых несортированных известняков, где сложно выделить отдельные, четко очерченные стадии экологической сукцессии. Биогермные образования часто чередуются с отложениями склонов построек. Основное влияние на экосистемы здесь оказывала интенсивность поступления биокластового материала с центральных и более возвышенных зон скелетных холмов. Шестиступенчатая трофическая структура этой части скелетных холмов характеризуется преобладанием продуцентов I уровня (кальцимик-робов, тубифитесов, цианобактерий). Продуценты II уровня представлены филлоидными водорослями. Гетеротрофные организмы имеют незначительное распространение и представлены консументами I (фораминиферами, брахиоподами), II (ветвистыми мшанками), III (гастроподами, остракодами, трилобитами) и IV (тонкопластинчатыми палеоаплизинами, аммонитами) уровней. Обломки члеников криноидей, колоний зеленых дазикладиевых и красных водорослей, раковин брахиопод, толстопластинчатых палеоаплизин и другие палеонтологические остатки являются аллохтонными и были привнесены с более возвышенных участков морского дна, т. е. с центральных зон скелетных холмов.
Контролирущие факторы
В разрезах скелетных холмов запечатлены важнейшие события (биогенные и абиогенные) и реакции экосистем на них. Как показали наши исследования, на эволюцию экосистем нижнепермских органогенных сооружений наряду с глобальными факторами [2] существенное влияние оказывали факторы регионального масштаба.
Контролирующие факторы разделяются на аутогенные и аллогенные (см. таблицу). К сожалению, на собранном материале очень трудно проследить всю цепочку причинно-следственных связей (событие/реакция). Как известно, гидродинамика среды зависит от изменения уровня моря, что может быть обусловлено как тектонической активностью, так и эвстатическими колебаниями. Только в одном случае достаточно уверенно удалось определить первопричину из-
менения структуры экосистем — локальное прогибание дна морского бассейна (см. таблицу).
Самым важным аутогенным фактором было развитие микробиальных пленок на сессильных организмах, что способствовало формированию биологически индуцированного синседимента-ционного кальцитового цемента [7] и образованию жесткого каркаса. Это определяло развитие специфических экосистем нижнепермских скелетных холмов с преобладанием сессильных организмов с арагонитовым и Mg-кальцитовым скелетом. В какой-то степени отрицательную роль в развитии экосистемы играли филлоидно-водорослевые сообщества, которые обедняли ее, но не приводили к коллапсу.
Наиболее разнообразными были внешние (аллогенные) факторы, контролировавшие развитие биоценозов. Так, в рассмотренных выше разрезах хорошо прослеживаются признаки того, что экосистемы нижнепермских скелетных холмов периодически подвергались существенному воздействию внешних факторов, в основном гидродинамических. Можно отметить некоторые неизбежные изменения экосистем, наиболее ярко представленные в центральных зонах нижнепермских скелетных холмов. Например, при повышении активности гидродинамики среды, как правило, происходила смена организмов с вертикальной формой роста пластинчатыми, что было связано с большей устойчивостью пластинчатых организмов к волнениям. Кроме того, подвижность вод способствовала удалению ядовитых веществ, выделяемых филлоидными водорослями. Это позволяло поселяться другим организмам. Другой гидродинамический индикатор — это базис действия волн, выше которого существовали уже активно-водные условия. В развитии рифов этот уровень определял повышение биоразнообразия экосистемы и соответствовал стадии диверсификации [30]. Для скелетных холмов нижняя граница базиса действия волн являлась финальной [29] в развитии экосистем, что приводило к формированию отложений стадии деструкции и коллапсу всей экосистемы [10, 11]. Рассматривая влияние аллогенных контролирующих факторов в различных зонах холмов (рис. 6), можно отметить четкую зависимость экосистем центральных зон от гидродинамической активности. Это подтверждает выводы Н. П. Джеймса и П. А. Бурка [30] о преимущественно вертикальной зональности холмов.
Возникает правомерный вопрос: если рост скелетных холмов в вертикальном направлении ограничивался бази-
Основные факторы, влияющие на развитие экосистем
|
Факторы |
Зоны |
Последствия |
|
АУТОГЕННЫЕ |
||
|
Появление микробиальных пленок на сессильных организмах |
Центральная |
Формирование каркаса с помощью биологически индуцированного цемента на всех стадиях развития органогенных построек |
|
Краевые |
Формирование каркаса с помощью биологически индуцированного цемента на стадиях колонизации и деструкции |
|
|
Укрепление субстрата |
Центральная и краевые |
Проявление стадии колонизации |
|
Филлоидно-водорослевые заросли |
То же |
Загрязнение вод, обеднение экосистемы |
АЛЛОТИГЕННЫЕ
|
Спокойная гидродинамика |
Краевые |
Развитие кальцимикробных сообществ с многоклеточными организмами, обладающими ветвистой, вертикальной и пластинчатой формами роста |
|
Центральная |
Стабилизация субстрата кальцимикробными сообществами с многоклеточными организмами, имеющими ветвистую форму роста |
|
|
Слабая гидродинамика |
Центральная и краевые (преимущественно восточные, реже западные) |
Развитие кальцимикробных сообществ и многоклеточных организмов с пластинчатой формой роста на стадии стабилизации Развитие прямостоящих, реже сетчатых форм роста на стадии колонизации |
|
Активная гидродинамика |
Краевые (западные) |
Коллапс экосистемы. Появление нетипичных сообществ |
|
Центральная |
Коллапс экосистемы |
|
|
Постепенная активизация гидродинамики |
Краевые (западные) и центральная |
Смена форм роста организмов с вертикальной на пластинчатую |
|
Постепенное снижение гидродинамической активности |
Краевые (западные, редко восточные) |
Смена пластинчатых форм роста вертикальными |
|
Центральная |
Проявление стадии колонизации без стадии стабилизации |
|
|
Резкое понижение уровня моря (регрессия) |
-—//-- |
Остановка роста органогенной постройки, коллапс экосистемы |
|
Незначительный привнос биокластового материала |
Краевые (в основном восточные, редко западные) |
Появление нетипичных экосистем на стадии колонизации. Циклическое повторение стадии стабилизации |
|
Повышенный привнос биокластового материала |
То же |
Коллапс экосистемы. Проявление стадии стабилизации с кальцимикробными сообществами |
|
Прогибание дна осадочного бассейна |
-—//-- |
Увеличение привноса биокластового материала |
сом действия волн, то почему они в дальнейшем не развивались в латеральном направлении, как рифы? Чтобы ответить на него, необходимо рассмотреть обстановки, в которых развивались раннепермские скелетные холмы. Так, В. А. Салдин [20], проанализировав морфологию дна Североуральского осадочного бассейна в раннепермское время, показал, что скелетные холмы были ограничены с запада мелководным открытым шельфом, а с востока — умеренно глубоководными депрессиями с конденсированным осадконакоплением. В конденсированных разрезах на Полярном Урале была обнаружена органогенная постройка типа илового холма, что позволило определить ее образование на глубинах не более 150 м [16]. Позднее при изучении общего строения нижнепермских скелетных холмов было выделено два типа их склонов [3, 12]: западный пологий, граничащий с открытым мелководным шельфом, и восточный крутой, соседствующий с депрессиями. Внешние факторы, контролировавшие развитие западных краевых зон холмов, были связаны не только с гидродинамикой. Существенное влияние оказывал здесь привнос биокластового материала, который приводил к появлению нетипичных стрессоустойчивых (цианобактериальных) сообществ на стадии колонизации, к ритмичному повторению
Основные внешние (аллотигенные) факторы, влияющие на экосистемы

Краевая западная зона Центральная зона Краевая восточная зона
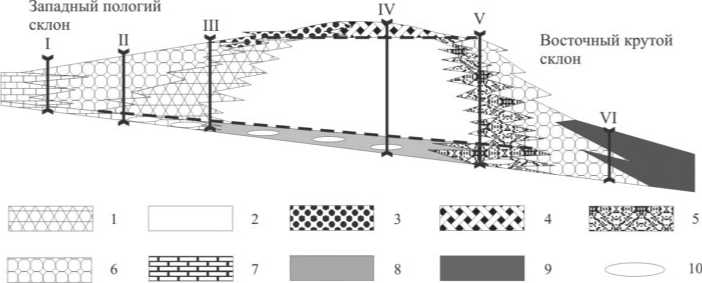
Puc. 6. Идеализированная схема строения нижнепермских скелетных холмов Северного Урала (по: Пономаренко, 2010; Антошкина и др., 2010; с дополнениями). I — обн. 24, р. Унья (среднеассельская часть); II — обн. 61, р. Печора; III — обн. 84, р. Илыч; IV — обн. 28, р. Унья (нижнеассельская часть); V — обн. 28, р. Унья (средневерхнеассель-ская часть); VI — обн. 36, р. Илыч (сакмарская часть)
1—4 — биоцементолиты: 1 — мшанковые, 2 — филлоидно-водорослевые, 3 — полибиокла-стовые, 4 — биокластовые палеоплизиновые; 5—8 — известняки: 5 — кальцимикробные, 6 — биокластовые склоновые, 7 — биокластовые открытого шельфа, 8 — пелитоморфные; 9 — депрессионные отложения; 10 — поселения стадии стабилизации
стадии стабилизации или к коллапсу экосистемы. Следует отметить, что основная масса биокластового материала сносилась с центральных зон скелетных холмов. Роль гидродинамических контролирующих факторов в восточных, а именно более глубоководных зонах скелетных холмов была невелика. Более важное значение в контроле над экосистемами имело количество поступавшего биокласто-вого материала. Основной реакцией экосистем этой зоны было проявление ритмичности в развитии стадии стабилизации с кальцимикробными комочками и сгустками, пластинчатыми и сетчатыми или ветвистыми метазоями как отражение периодичности поступления скелетного материала. То есть латеральное развитие нижнепермских скелетных холмов ограничивалось поступлением биокла-стового материала с центральных зон. Можно сказать, что в этом случае экосистема ограничивала сама себя.
Таким образом, экологическая сукцессия нижнепермских скелетных холмов контролировалась как аутогенными, так и аллогенными факторами. Аутогенная сукцессия пионерных сообществ проявлялась только в те этапы развития морского бассейна, когда внешние физико-химические условия были относительно стабильными. Об этом свидетельствует направленный процесс развития сообществ и видов от стадии стабилиза- ции до стадии колонизации. Значительные изменения внешних факторов, связанные скорее всего с тектонической активностью во время проявления уральского коллизионного орогенеза, приводили к прерыванию экологической сукцессии или к появлению нетипичных для нижнепермских скелетных холмов цианобактериальных сообществ. Исходя из вышесказанного, экологическую сукцессию нижнепермских скелетных холмов Северного Урала можно определить как алло-аутогенную. Как пример нижнепермской преимущественно аутогенной экологической сукцессии можно назвать только разрез нижнепермского илового холма Розя Кырта [13].
Если развитие рифов контролировало обстановки седиментации на платформе, то формирование скелетных и иловых холмов, напротив, зависело от обстановок седиментации.
Заключение
Основные результаты исследований сводятся к следующему:
-
1. Нижнепермские органогенные постройки типа скелетных холмов формировались на склонах депрессий и соседствовали на западе с мелководным открытым шельфом, на востоке с умеренноглубоководными депрессиями.
-
2. В строении нижнепермских скелетных холмов Северного Урала выделяются зоны западного пологого склона, западной краевой зоны, центральной зоны, восточной краевой зоны и восточного крутого склона. Типичные раннепермские экосистемы развивались только в центральной зоне.
-
3. На экосистемы и экологическую сукцессию оказывали сильное влияние внешние, аллогенные факторы. На развитие центральных зон очень интенсивно воздействовали гидродинамические факторы, в краевых зонах более важную роль играло количество поступаемого биокластового материала. Развитие скелетных холмов в вертикальном направлении ограничивалось базисом действия волн, в латеральном — засыпанием экосистемы биокластовым материалом, поставщиком которого были центральные зоны скелетных холмов.
-
4. Наиболее важным аутогенным фактором является развитие микробиальных пленок, которое приводило к формированию каркаса и способствовало появлению своеобразной экосистемы раннепермских скелетных холмов из мелких сессильных организмов.
-
5. Экологическая сукцессия нижнепермских скелетных холмов определяется как алло-аутогенная. Аутогенная сукцессия, характеризующая стадии стабилизации и колонизации, проявлялась в этапы развития осадочного бассейна, когда внешние физико-химические условия не претерпевали значительных изменений. Влияние внешних факторов приводило к проявлению аллогенной сукцессии, на которую указывают стадия деструкции и появление стрессоустойчивых (цианобактериальных) экосистем. Повторяемость стадий экологической сукцессии позволяет относить нижнепермские рифогенные толщи к ритмитам скелетных холмов.
Исследования проводились в рамках программы Президиума РАН № 15/2 № 09-П-5-1008.
Список литературы Эволюция экосистем нижнепермских скелетных холмов Северного Урала
- Антошкина А. И. Рифообразование в палеозое (на примере севера Урала и сопредельных территорий). Екатеринбург: УрО РАН, 2003. 303 с.
- Антошкина А. И. Причинно-следственная связь дискретности палеозойского рифообразования на севере Урала//Рифы и карбонатные псефитолиты: Материалы Всерос. литол. совещ. Сыктывкар: Геопринт, 2010. С. 15-17.
- Антошкина А. И., Пономаренко Е. С., Рябинкина Н. Н. и др. Эволюционный тренд палеозойской рифовой экосистемы как отражение эволюции гео-биологических систем на примере севера Урала. Сыктывкар: Геопринт, 2010. Отчетная серия № 2 (83). 44 с.
- Варсанофьева В. А. Геологическое строение территории Печоро-Илычского государственного заповедника//Тр. Печоро-Илычского гос. заповедника. Вып. 1. М., 1940. С. 5-214.
- Калашников Н. В., Михайлова З. П. К стратиграфии верхнего карбона р. Уньи//Геология Северо-Востока европейской части СССР и севера Урала. Сыктывкар, 1971. С. 55-65. (Тр. Ин-та геологии Коми фил. АН СССР. Вып. 14).
